1. Introduction
Weaning is one of the most critical periods in the piglet life, with severe consequences on performance throughout their full productive lifetime. In commercial farms, piglets are weaned around 21–28 days of life. At this early age, weaning entails a big challenge for the pig, as they are exposed to new social partners, an abrupt dietary change, and an immature immune system [
1]. As a consequence of the high stress to which pigs are subjected, a decreased feed intake is frequently observed. These circumstances are associated with a decrease in nutrients supply and their digestive capacity, a low weight gain, and a high diarrhea incidence, which could even lead to death [
2]. The emergence of opportunistic pathogens during this stage, such as
Escherichia coli, can trigger post-weaning diarrhea (PWD), which is also called post weaning enteric colibacillosis [
3].
To prevent and avoid the occurrence of these series of events, antimicrobial compounds have been extensively used in the past, either as antibiotic growth promoters (AGPs) or at sub-therapeutic quantities to increase animal growth rates and to improve feed efficiency. Nevertheless, due to the strong political and social pressure to prevent antibiotic resistance in pathogenic microbiota, the supplementation of AGPs in animal feed was finally banned in the European Union, and in the most recent years, different programs have been implemented looking for a responsible use of antibiotics in livestock (European Surveillance of Veterinary Antimicrobial Consumption (ESVAC) project) [
4]. This decision lessened the productivity and profitability of animal production systems [
5] but also provided an opportunity to implement better zootechnical practices and fundamental research to develop new feed additives (i.e., probiotic, prebiotic, acidifiers, enzymes, phytogenic, etc.) as an alternative to AGPs in animal production [
6].
Phytogenic feed additives (often called phytobiotics or botanicals) are plant-derived compounds that may have positive effects on animal growth and health due to their antibacterial, anti-inflammatory, and antioxidant properties [
7,
8]. In addition, their aromatic and oily characteristics have been shown to increase the palatability of feed with no flavor or aroma difference in the finished meat product [
9]. Phytogenic substances utilized in phytogenic feed additives include herbs, spices, essential oils, and non-volatile extracts [
10].
Studies evaluating the effects of phytogenic supplementation in swine have reported in many occasions positive results in growth performance, feed conversion ratio, and nutrient digestibility [
11], amongst others. Positive results in performance have been attributed to different modes of action including direct antimicrobial effects on particular bacterial groups [
12], positive effects on intestinal morphology and intestinal function [
13], immunomodulatory activities in the host [
14], and antioxidant properties [
15]. Moreover, the effects of phytogenic compounds on maternal transfer have also been studied as an effective alternative way to improve the response of the offspring [
16]. However, a closer analysis of the action of phytogenics on intestinal health and particularly on the microbial environment and the cross-talk with the host is necessary to understand how plant extracts can influence animal performance.
Thus, following the hypothesis that it is possible to improve the response of piglets to weaning and to reduce the incidence of post-weaning diarrhea by including phytogenic blends in the diet as an alternative to antimicrobials, the objective of the present study was to evaluate, in weaned piglets, the potential of two plant-based in-feed additives in front of an oral ETEC F4 challenge, analyzing their effects on performance, clinical response, immune system, and gut health. For that, we compared two diets supplemented with different phytogenic blends with a non-supplemented diet and a diet including pharmacological levels of ZnO as a positive control considering its largely known efficacy to prevent post-weaning colibacillosis [
17].
2. Materials and Methods
The trial was performed at the Experimental Unit of the Universitat Autònoma de Barcelona (UAB) conducted as a Level 2–High Risk Biosecurity Procedure. It received prior approval (Permit No. CEEAH: 4026 DMAH: 10118) from the Animal and Human Experimental Ethical Committee of this Institution. The treatment, management, housing, husbandry, and slaughtering conditions conformed to European Union Guidelines [
18]. All efforts were made to minimize animal suffering.
2.1. Animals and Housing
For the trial, 96 male piglets ((Landrace × Large White) × Pietrain) were obtained from a commercial farm, from non-vaccinated sows with E. coli. The piglets were weaned at 21 days of age and had an average body weight (BW) of 4.8 ± 0.62 kg. Piglets were transported to facilities of the UAB, identified, weighed, and distributed among 32 pens (3 animals per pen) within 4 rooms (8 pens per room) on arrival. Each pen was readjusted by weight to start the experiment with the same initial average weight in all pens. Each pen (3 m2) had a feeder and a water nipple to provide food and water ad libitum. The weaning rooms were equipped with automatic heating, forced ventilation, and an individual heat-light per pen. The experiment was conducted during the winter season (January), with an average room temperature of 28 °C ± 4 °C.
2.2. Experimental Diets
Four dietary treatments were included in the study: (1) a control basal diet (T1); (2) T1 supplemented with ZnO (as ZINCOTRAX at 3100 mg/kg of feed, equivalent to 2500 ppm Zn/kg of feed) (T2); (3) T1 supplemented with plant supplements based on essential oils (ColiFit Icaps C, 1 kg/tm) (T3); and (4) T3 supplemented with plant supplements based on non-volatile compounds (Phyto Ax’Cell, 1.5 kg/tm) (T4).
All diets were given in a mash form. Basal diet (
Table 1 and
Table 2) was formulated to satisfy the nutrient requirement standards for pigs [
19]. Basal diet was made in a single batch and subsequently divided into four batches to include the different additives to form the four experimental diets. Before being included in the mixer, the required amount of the additives was pre-mixed by hand in approximately two kg of basal diet. Chemical analyses of the diets (
Table 2) including dry matter (DM), ash, gross energy, crude protein, and crude fat (diethyl ether extract) were performed according to the Association of Official Agricultural Chemists standard procedures [
20]. Neutral-detergent fiber and acid-detergent fiber were determined according to the method of Van Soest et al. [
21].
2.3. Plant Supplements Analyses
The tested products, ColiFit Icaps C and Phyto Ax’Cell, were two commercial products supplied and analyzed by Phytosynthese, Mozac, France.
The product ColiFit Icaps C consisted of a blend of essential oils rich in trans-cinnamaldehyde, eugenol, carvacrol, thymol, and diallyl disulfure. Phyto Ax’Cell (Phytosynthese, Mozac, France) was based on a blend of standardized plants, plant extracts, and green propolis extract. The active compounds contained in the product were curcuminoids, carnosic derivatives, naringin flavonoids, salicylic derivatives, and artepillin-C.
The quantification of these active compounds was performed by Phytosynthese lab with high liquid chromatography or gas chromatography according to components. Detailed description of the methodology can be found in
Appendix A.
2.4. Bacterial Strain
For the trial, the enterotoxigenic
Escherichia coli (ETEC) F4 strain COLI30/14-3 used (positive for virulence factors F4ab, F4ac, LT, STb, and EAST1 and negative for K99, F6, F18, F41, STa, VT1, VT2, and EAE) was isolated from feces of a 14-week-old pig from a farm with a clinical course of colibacillosis and provided by the Veterinary Laboratory of Diagnosis of Infectious Diseases of UAB. This strain was actually used with the same purpose in previous studies of our group [
22]. The oral inoculums were prepared by an overnight incubation at 37 °C in Brain Heart Infusion (BHI) with slow agitation (1×
g) in an orbital incubator. The final inoculum was 2 × 10
8 CFU/mL. The final culture broth was used as the oral inoculums by preparing 96 doses of 6 mL. To quantify the inoculums (CFU/mL), serial dilutions were cultured in Tryptic Soy Agar (TSA) plates (overnight, 37 °C).
2.5. Experimental Design, Procedure, and Sampling
Pens were allocated to the 4 treatment groups. Animals received the experimental diets over 16 d, mortality rate was registered, and no antibiotic treatment was administered to any animals in the trials. After the adaptation period (experimental days 0 to 7), all animals were orally challenged with the pathogen ETEC F4 strain COLI30/14-3 as a single dose (1 × 109 CFU). In order to ensure that the stomach was full at the time of inoculation and thus facilitate bacterial colonization, feed withdrawal was performed at 21:00 h of the previous day and provided back 30 min before inoculation.
One animal of each pen (8 per treatment) was euthanized on day 8 post-inoculation (PI). Feed intake by pen was registered at day 0, 7, 8, 9, 10, 11, 14, and 15, and individual weight was measured on day 0, 4, and 8 PI. The average daily gain (ADG), average daily feed intake (ADFI), and gain–feed ratio (G:F) were calculated for each pen.
After the ETEC F4 challenge, animals were checked daily for clinical signs to evaluate their PI status (i.e., dehydration, apathy, and diarrhea). Fecal consistency was registered on day 0, 1, 2, 3, 4, and 7 PI; score consistency was measured using a scale from 1 to 4: 1 = normally shaped feces, 2 = shapeless soft feces, 3 = thin or liquid feces, and 4 = very liquid feces (translucid) or with blood. Rectal temperature was assessed before the challenge (d 0 PI) and day 1 and 2 PI with a digital thermometer. Blood samples were taken on day 4 and 8 PI from the initial intermediate BW piglet from each pen. For microbiological analysis, fecal samples were taken aseptically before inoculation at day 0, and at days 4 and 8 PI. Samples were always collected from the same piglet from each pen, corresponding to that with intermediate BW the first day of the trial. Fecal samples were collected after spontaneous defecation associated with manipulation of the animal or by rectal stimulation. Samples were first stored in ice until analysis.
On day 8 PI, the pig of medium weight of each pen on arrival (N = 32) was euthanized. Sampling took place during the morning (between 8:00 and 14:00). Prior to euthanasia, a 10 mL sample of blood was obtained by venipuncture of the cranial vena cava using 10 mL tubes without anticoagulant (Aquisel; Madrid, Spain), and immediately after sampling, piglets received an intravenous, lethal injection of sodium pentobarbital in the same vein (200 mg/kg BW; Dolethal, Vetoquinol S. A.; Madrid, Spain). Once dead, the animals were bled, their abdomens were immediately opened, and the whole gastrointestinal tract was excised. Digesta (approximately 40 mL) from the ileum and proximal colon (considered to be 0.75 m from the ileocecal junction) was homogenized, and samples were collected for bacterial counts. The pH of the homogenized digesta content was immediately determined with a pH-meter (Crison 52-32 electrode, Net Interlab; Barcelona, Spain). For histological study, 3 cm sections were taken from the proximal ileum and proximal colon, were opened longitudinally, washed thoroughly with sterile phosphate-buffered saline (PBS), and fixed by immersion in a formaldehyde solution (4%). Blood samples were centrifuged (3000× g for 15 min at 4 °C) to obtain serum, and the serum obtained was divided into different aliquots and stored at −80 °C. For intestinal gene expression analysis, 3 cm long sections were removed from the mid jejunum, opened longitudinally, and washed exhaustively with sterile PBS. A small piece of fresh tissue was cut at around 0.5 cm2 and placed in 5–10 volumes of RNAlater solution. Samples were stored at 4 °C overnight (to allow the solution to penetrate the tissue) and then moved to −80 °C until RNA isolation. For analyzing enterobacteria, coliforms and E. coli F4 attached to the ileal mucosa, 5 cm long sections of ileum were collected from each animal, washed thoroughly three times with sterile PBS, opened longitudinally, and scraped with a microscopy glass slide to obtain the mucosa scraping contents. One aliquot of each scraping sample was kept on ice until traditional microbiological analysis, and a second aliquot was kept on dry ice and subsequently in the freezer (−80 °C) until DNA analysis.
2.6. Analytical Procedures
For microbial counts of enterobacteria and total coliforms, the ileum and colon contents, ileum scrapings, and feces were suspended in PBS (1:10) and homogenized for 5 min. Thereafter, 10-fold serial dilutions were made in PBS to seed in chromogenic and Rogosa agar for E. coli and lactobacilli counting, respectively. Counts were read after 24 h of incubation at 37 °C.
For morphological measures, tissue samples were dehydrated and embedded in paraffin wax, sectioned in 4 µm thick slices, and stained with hematoxylin and eosin. Morphological measurements of ileal sections were performed with a light microscope (BHS, Olympus) using the technique described in Nofrarías et al. [
23]. Measured parameters included villus height, crypt depth, villus/crypt ratio, intraepithelial lymphocytes (IEL), goblet cells (GC), and mitosis.
Serum concentrations of tumor-necrosis factor-α (TNF-α) were determined using Quantikine Porcine TNF-α kits (R&D Systems). The Pig major acute-phase protein (Pig-MAP) concentration was determined using a sandwich-type ELISA (Pig MAP Kit ELISA, Pig CHAMP Pro Europe S.A.), according to the manufacturer’s instructions.
Total RNA was extracted from 50 mg of mid jejunum tissue through homogenization with a Polytron device (IKA, Staufen, Germany) in 1 mL of TRIzol reagent (Thermo Fisher Scientific, Waltham, MA, USA), followed by of the use of the Ambion RiboPure Kit (Thermo Fisher Scientific, Waltham, MA, USA) according to the manufacturer’s protocol. The quantification and purity valuation of the extracted RNA was assessed by measuring the absorbance at 230, 260, and 280 nm using a NanoDrop ND-1000 spectrophotometer (NanoDrop Technologies Inc., Wilmington, DE, USA). The RNA integrity was checked with Agilent Bioanalyzer-2100 equipment (Agilent Technologies, Santa Clara, CA, USA), following the producer’s protocol.
For cDNA synthesis, 1 μg of total RNA was reverse-transcribed into cDNA in a final volume of 20 µL. A High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA, USA) and random primers were used, and the following thermal profile was applied: 25 °C for 10 min; 37 °C for 120 min; 85 °C for 5 min; 4 °C hold. cDNA samples were stored at −20 °C until use.
Intestinal gene expression analysis was made using a custom OpenArray plate. A total of 56 genes (including 4 house-keeping genes:
ACTB,
B2M,
GAPDH, and
TBP) related to intestinal health were selected, as described by Reyes-Camacho et al. [
16] (
Table A1). The description of forward and reverse primers for each gene is found in
Table A2. One replicate per sample was run in a Taqman Open Array gene expression custom plate format for gene expression with 56 assays of 48 samples per plate (OpenArray plate) in a QuantStudio 12K Flex Real-Time PCR System (Applied Biosystems, Foster City, CA, USA). Gene expression data were analyzed using the ThermoFisher Cloud software 1.0 (Applied Biosystems, Foster City, CA, USA) applying the 2
−ΔΔCt method for relative quantification (RQ) and using the sample with the lowest expression as a calibrator.
2.7. Statistical Analyses
Microbiological counts were log transformed for analysis. The general linear and/or the mixed models of SAS version 9.2 (SAS Institute Inc., Cary, NC, USA) were used to analyze the effect of experimental treatments on different parameters except gene expression (see below). The diet was considered a fixed effect, and random effect was used to account for variation between pens. When treatment effects were established, treatment means were separated using the probability of differences function adjusted by Tukey–Kramer. The pen was considered the experimental unit. The α-level used for the determination of significance was p = 0.05. The statistical trend was considered for p < 0.10.
For gene expression statistical analysis, the open-source software R v3.5.3. was used. The RQ data matrix was used and normalized accordingly to the reference genes. Genes with normal distributions were analyzed using One-Way ANOVA, while genes with non-normal distributions were analyzed using the non-parametric Kruskal–Wallis test. p-values were adjusted using the False Discovery Rate (FDR) method, and Tukey tests were performed for genes with significant differences between treatments.
4. Discussion
The present study aimed to evaluate the efficacy of two plant-based feed supplementations in comparison to ZnO to fight colibacillosis in weaned piglets by using an experimental model of disease. To assess this objective, two different phytogenic additives containing extracts and essential oils from plants were tested in this trial; T3 included a blend of different essential oils while T4 contains in addition non-volatile compounds with putative antioxidant, anti-inflammatory, and immunomodulatory properties.
In general terms, performance was not significantly affected by the dietary treatments compared to the plain diet before or after the challenge. The limited number of replicates in this kind of controlled-challenge trials probably precluded us to detect significant differences in performance. Despite this, some numerical trends were detected for treated groups with ZnO or plant supplements.
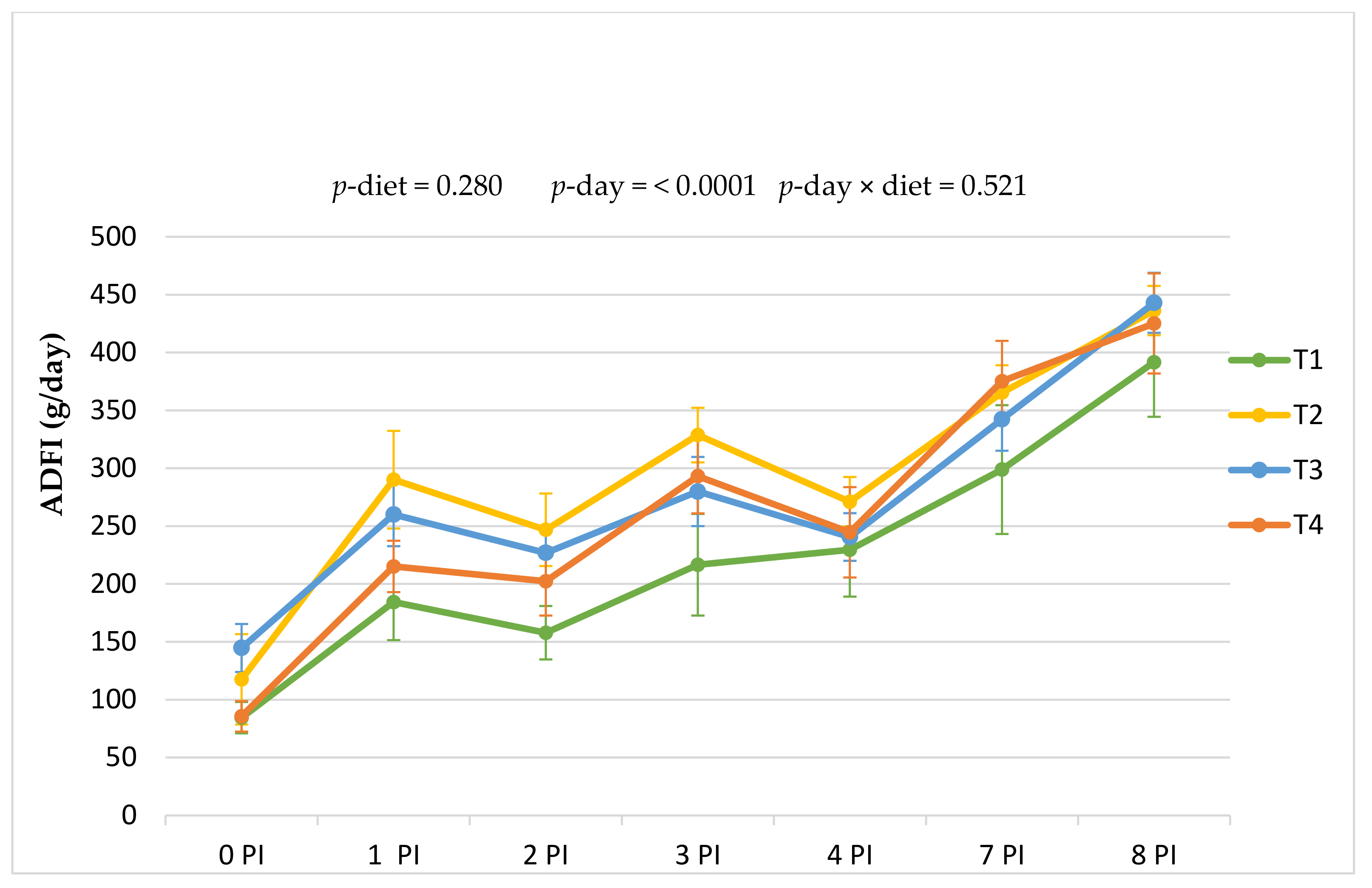
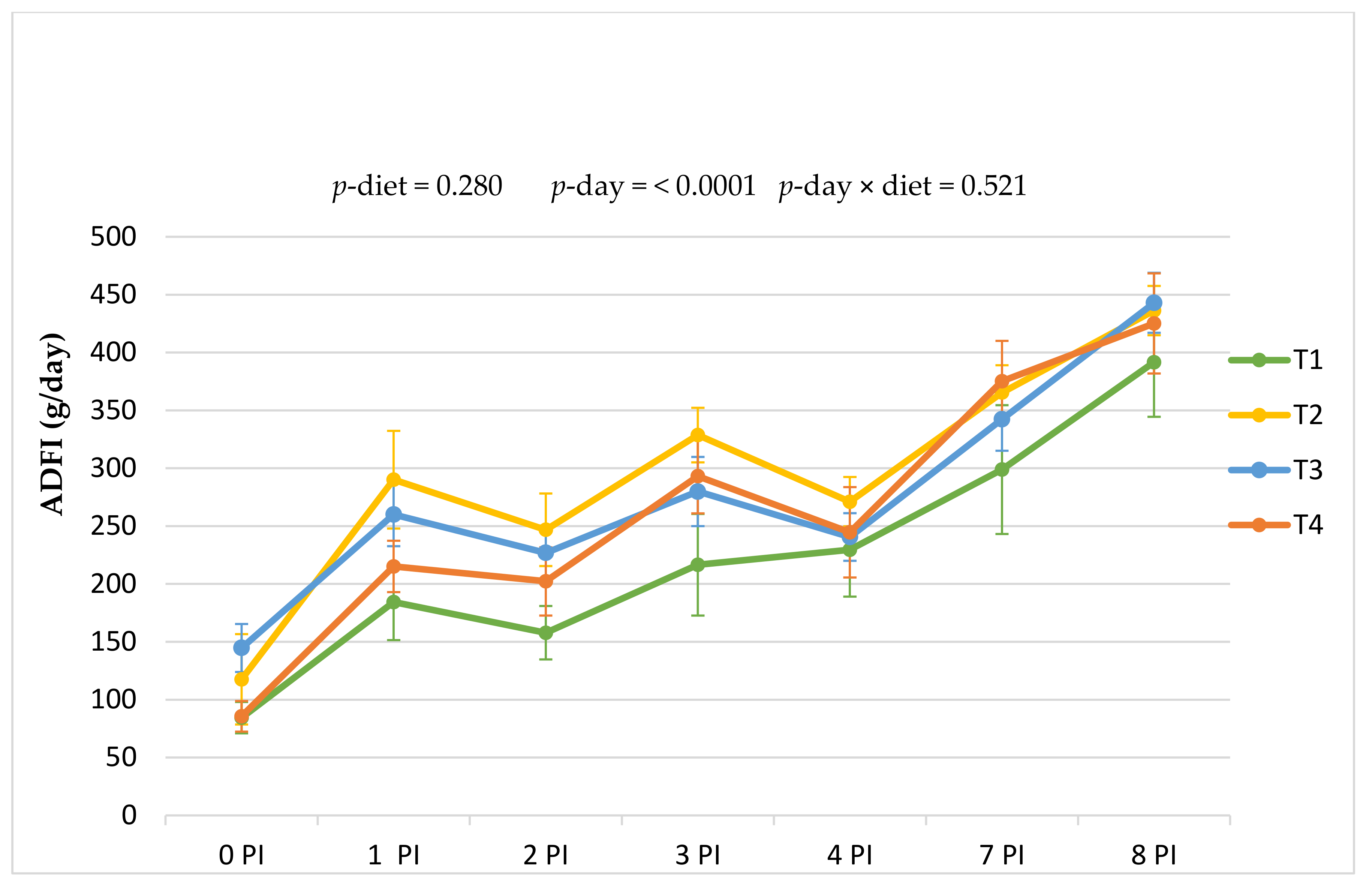
Regarding the impact of phytogenics on feed intake, results found in the literature on ADFI in pigs are variable. In the present study, the dietary inclusion of the two botanic feed supplements did not significantly affect the overall ADFI intake, which is in line with other studies in weaned piglets [
8,
25,
26,
27]. However, Platel and Srinivasan [
28] described how many botanical compounds and spices can improve food intake in humans and other mammal species. A digestive stimulant action mediated by an increase of salivary, gastric, or bile secretions, and reduction in food transit time, may result in a stimulation of appetite. In this regard, Cho et al. [
29], Kommera et al. [
30], and Maenner et al. [
31] reported an enhanced ADFI with the use of phytogenics containing fenugreek, clove, cinnamon; anis oil, citrus oil, oregano oil, natural flavor; and menthol and cinnamon aldehyde, respectively. In contrast to this, other authors observed a decrease of the ADFI in weaned piglets due to the supplementation of cinnamon, thyme, oregano; and buckwheat, thyme, curcuma, black pepper, and ginger, respectively [
32,
33]. Therefore, the results found in the literature are controversial and probably respond to different products, blends, and doses tested.
Regarding ADG, in this work, we did not find statistical differences between treatments in the adaptation week, but differences were found in the period immediately after the challenge (0–4 PI period;
p = 0.035)). Four days after the challenge, the T2 ADG (including ZnO) was improved significantly compared to the basal diet T1, while animals receiving plant blends (T3 and T4) showed intermediate values. These results are in concordance with other studies that evaluated the effect of the addition of essential oils [
8] and phytogenic products based on plant extracts plus essential oils as anise oil, clove oil, citrus oil, and oregano oil, amongst others [
23,
30,
31,
34]. In contrast, Namkung et al. [
32] described a decrease of the ADG in weaned piglets, which were offered a diet with a herbal extract containing cinnamon, thyme, and oregano. Other authors have not found differences in weaned piglets fed diets supplemented with different essential oil blends [
35,
36], which is concurrent with other nursery pig studies involving phytogenic feed additives in which no effect on ADG and/or BW and/or ADFI were reported [
9,
26,
37,
38].
The increase of ADG in the supplemented diets was also reflected in the significant increment of gain/feed ratio (G:F) during the 0–4 PI period in the piglets receiving the T2 diet compared to T1, while plant-supplemented diets (T3 and T4) showed again intermediate values. In this respect, Kommera et al. [
30] also reported an improvement of the G:F in weaned piglets that were offered diets containing anise, citrus, and oregano oils. In contrast, Namkung et al. [
32] attributed negative feed efficiencies found for piglets during the first post-weaning week to the strong smell of cinnamon, thyme, and oregano, which was consistent with the findings of other works that also reported lower G:F in groups of piglets offered diet supplemented with essential oils (cinnamaldehyde and thymol being the main active components) [
8,
39] or anise [
40].
In our study, increases in ADG and G:F observed with T2 and numerically with T3 and T4 were only found in the period immediately post-challenge but not the week before and neither in the 4–8 PI period. Therefore, these changes can be interpreted as an improvement of the animal response to the pathogen challenge. As we see, there is a wide variability on the performance results reported among authors due to the supplementation of phytogenic compounds in the swine diets. Clouard and Val-Laillet [
27] suggested that several factors, such as animal characteristics (i.e., age, sex, physiological status), experimental conditions, time of exposure and particularly dosage, and biochemical features of phytogenic might be decisive factors in the development of pig performance. In addition, the diversity of plant materials and the lack of description of tested substances available in the literature make it difficult to compare these results with other phytogenics.
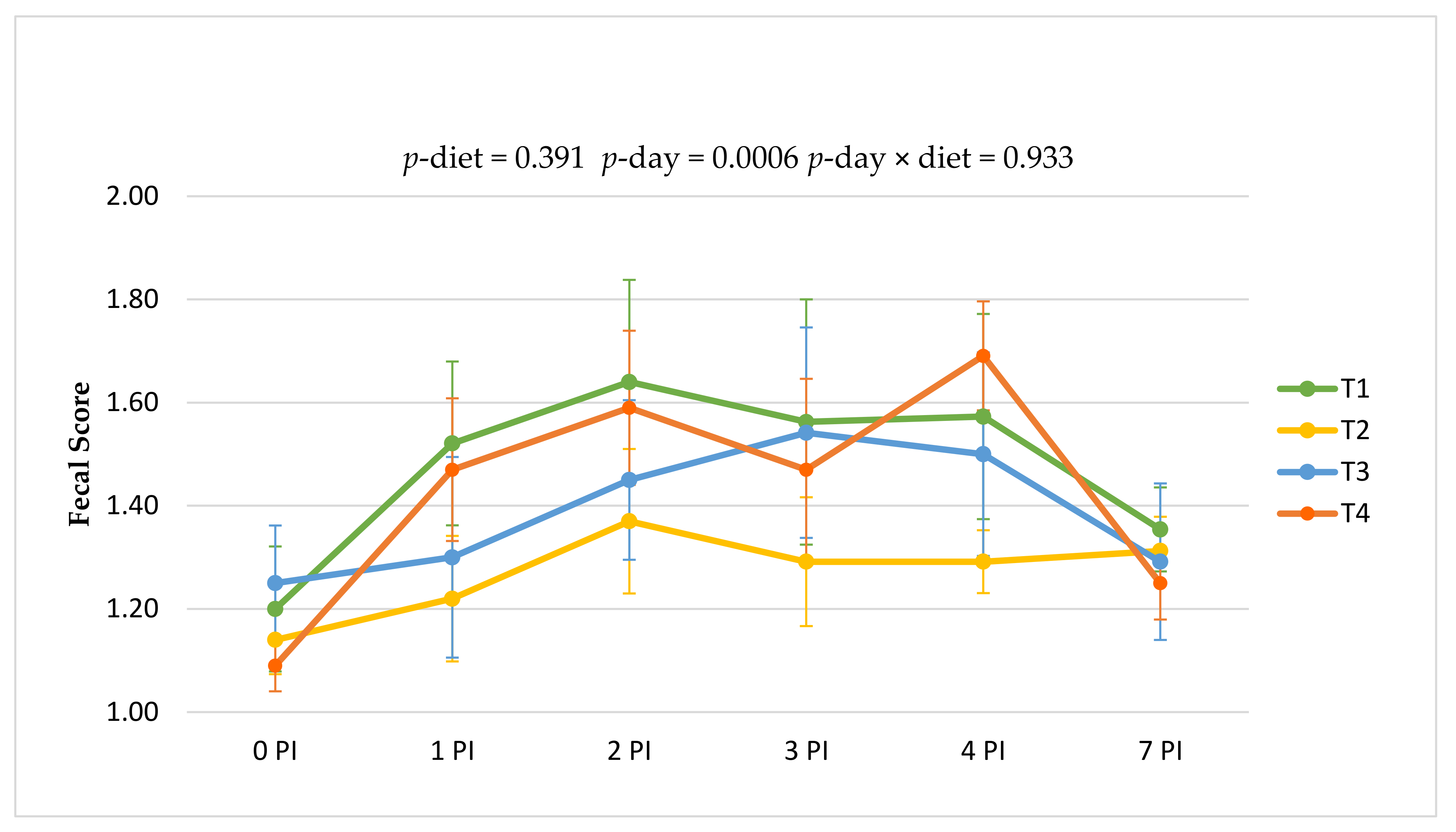
Post-weaning diarrhea is one of the many interdependent factors causing the high mortality rate in piglets. The addition of phytogenic feed additives has been reported to control clinical diarrhea [
8,
39,
41]. The reasons for such improvement are most likely associated with the reduction of
E. coli load in the gut, especially when piglets are raised under relatively poor environmental conditions. Lee et al. [
42] reported that both thymol and cinnamaldehyde have antimicrobial and anti-inflammatory effects. Different studies report that different combinations of essential oils exhibit antimicrobial in vitro activity against potentially pathogenic bacteria such as
E. coli,
Clostridium perfringens,
Salmonella,
Listeria, or
Staphylococcus, while they do not inhibit the growth or had less activity against beneficial microbes such as
Enterococcus,
Bacillus,
Lactobacillus, and
Bifidobacterium [
43,
44]. Specifically, a study made by Girard et al. [
45] with the same plant supplement as diet T3 (
ColiFit Icaps C) was able to inhibit
E. coli growth even at sub-MIC (minimum inhibitory concentration) concentrations with a bacteriostatic action. This effect was correlated to a reduction of membrane permeability of
E. coli with a significant degradation of their membrane polarity [
46]. Moreover Kerros et al. [
47], also with this essential oil mix, evidenced MICs on
Lactobacillus species equal or higher than those determined for pathogens
E. coli and
Clostridium jejuni, providing evidence of the selectivity of this phytogenic. According to this observation, in the present study, the numbers of lactobacilli in feces and colon digesta on day 8 PI were the highest with diets including the plant additives (T3 and T4) and the lowest with the T2 treatment. The lactobacilli/coliforms ratio, as a potential index of the microbiota balance, was also found to be increased at day 8 PI in feces by the T3 treatment when compared to the basal diet, showing diet supplemented with T4 intermediate values. However, in our study, we were not able to demonstrate a significant impact of the treatments on incidence of diarrhea prevalence or fecal score. Other authors also have failed to demonstrate significant improvement in these parameters supplementing phytogenic mixtures [
25,
26,
29,
30,
48,
49]. However, we cannot discard that limitations in the experimental models of disease and in the number of replicates would have precluded us from identifying significant changes.
The inclusion of phytogenic in the current diets did not affect the serological concentrations of cytokine TNF-α but showed a trend in the major acute-phase protein Pig-MAP to decrease at day 4 PI with T4 (
p = 0.07). These results would suggest that this phytogenic blend could exhibit anti-inflammatory properties in front of the tissue injury promoted by the ETEC challenge, since in swine, Pig-MAP have been related to acute inflammatory processes and also to the extent of tissue injury [
24]. In addition, recent study has demonstrated the usefulness of this biomarker to determine the intestinal injury degree and barrier integrity in recently weaned pigs subjected to an ETEC oral challenge [
50]. To appreciate this result, other authors have described how phytogenic agents could exert immune-modulating properties. In this regard, Machado et al. [
51] demonstrated that two Brazilian Green Propolis extracts containing p-coumaric acid, Artepillin-C, and other minor compounds could exert a strong local and systemic anti-inflammatory action from an immunomodulatory action on pro-inflammatory (IL-6 and TNF-
α) and anti-inflammatory (TGF-
β and IL-10) cytokines. Likewise, Hori et al. [
52] demonstrated that a Propolis Standardized Extract (EPP-AF), including caffeic, p-coumaric, trans-cinnamic acids, aromadendrin, and artepillin C could reduce the IL-1β secretion in mouse macrophages.
The supplementation with ZnO (T2) had a positive effect on the intestinal architecture, particularly for the villus/crypt ratio. In addition, we also found significant improvements with a T4 diet, and a trend with T3. These results agree with those from other studies conducted by Zeng et al. [
8] and Li et al. [
39], in which dietary supplementation of essential oils improved the villus/crypt ratio in the jejunum of weaned piglets. Particularly, the treatment T4, which showed a higher impact, could have prevented the intestinal damage probably mediated by a better controlled inflammatory response according to the reduction also observed in Pig-MAP values. It is important to remark these improvements in the epithelial integrity shortly after weaning, as they could be regarded as a very positive indicator of the potential of the tested supplements to prevent post-weaning diarrhea. After weaning, a transient reduction is produced in villus height as well as an increase in crypt depth [
53], which make animals more susceptible to pathogen infection and also decrease the absorptive capacity of the intestine with a reduction in feed efficiency [
54]. Intestinal morphology improvement has been observed by Maneewan et al. [
55] using turmeric in nursery pigs. Curcuminoids from turmeric may play a positive role on intestinal IL-1β [
56] correlated with a favorable effect on tight junctions in a CaCO
2 cell model [
57].
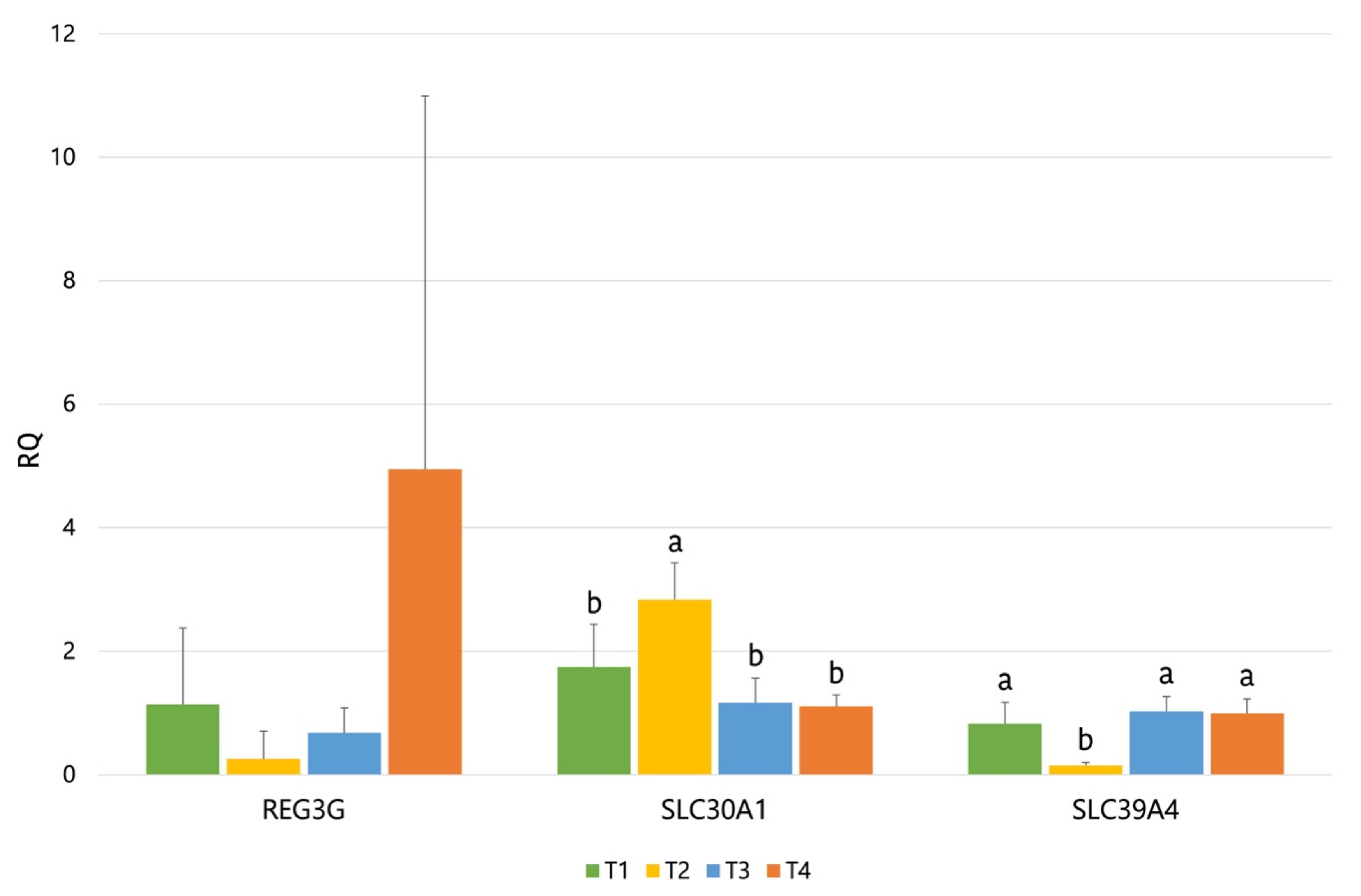
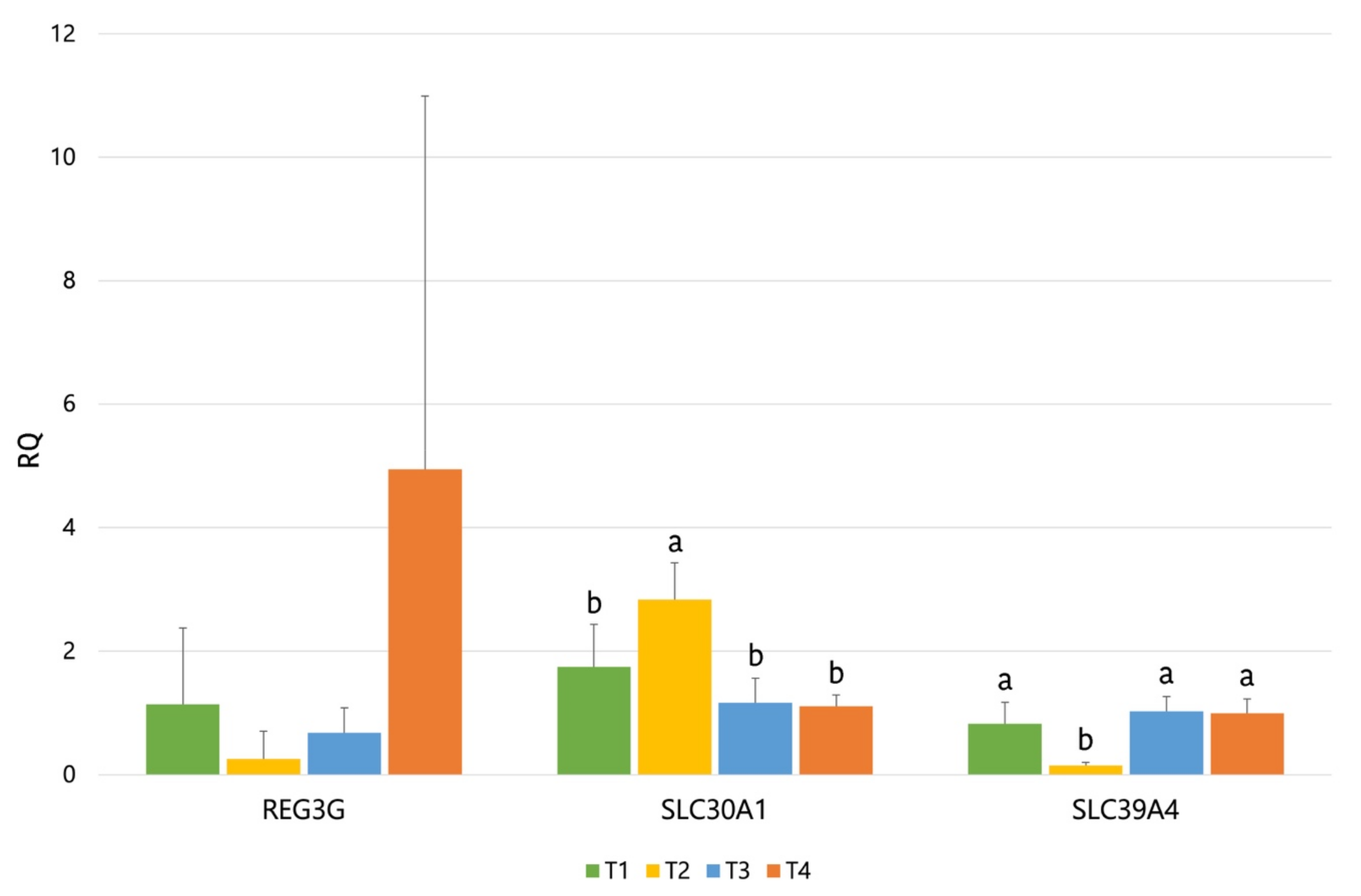
Concerning gene expression, our results showed an upregulation in
SLC30A1 by ZnO supplementation and a downregulation for
SLC39A4. Both
SLC30A1 and
SLC39A4 codify for solute carrier proteins, which are involved in the cellular transport of Zn. On the one hand,
SLC30A1 (Solute Carrier Family 30 Member 1) is involved in the transport of intracellular zinc into the extracellular matrix, and it has been shown to be upregulated by high dietary ZnO [
58]. On the other hand,
SLC39A4 (Solute Carrier Family 39 Member 4) is involved in the Zn uptake from the gut lumen and has been shown to be downregulated by high dietary ZnO in piglets [
58] and particularly in piglets challenged with ETEC [
59]. Therefore, these results are according to what would be expected from a high supplementation of ZnO (3100 ppm). The
REG3G gene also showed a numerical trend of being upregulated by the T4 diet. C-type lectins of the
REG3 family function as a barrier to protect body surfaces against microorganisms, modulating host defense process via bactericidal activity. A wide range of studies indicate that the
REG3G family plays an important role in the physical segregation of microbiota from the host as well as in the immune response induced by enteric pathogens [
60]. Moreover, this gene has been reported to be upregulated by ETEC challenge [
61] and also by dietary prebiotics (in mice) [
62]. Particularly, these last authors associated the increased expression of
REG3G to the changes observed in the microbiota with the prebiotic that could have counteracted the inflammation induced by high-fat diets. In the present study, it could be hypothesized that the increase observed in the
REG3G expression could be correlated to the pathogen control, to the intestine preserved from damages induced by the ETEC challenge (higher villus/crypt ratio), and to the lower expression of Pig-MAP (
p < 0.07) observed with the T4 diet.
The results of this study evidence that plant supplements can exert positive effects in front of an ETEC F4 challenge. However, it is difficult to elucidate which specific modes of action could had been involved, and moreover which active components of these blends would be primary responsible. Complex interactions between the microbiota and the host probably are behind some of these effects, together with a direct impact of these active compounds on the host mediated by their known antioxidant and antinflammatory properties. Further research is needed to fully understand the high potential of phytogenics to improve animal health and particularly to improve the adaptation of young piglets to weaning.


 ,
, 

{kind=link}
{kind=link}
{kind=link}