
A Standard Scale to Measure Equine Keeper Status and the Effect of Metabolic Tendency on Gut Microbiome Structure


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Development of the Equine Keeper Status Scale
2.2. Development and Testing of the EKSS
2.2.1. Sampling Protocol
Diet Estimations
2.2.2. Statistical Analysis
2.3. Microbiome Surveying of EKSS Horses
2.3.1. Sampling Protocol
2.3.2. DNA Extraction and Sequencing
2.3.3. Bioinformatic and Statistical Analysis
3. Results
3.1. Development and Testing of the EKSS
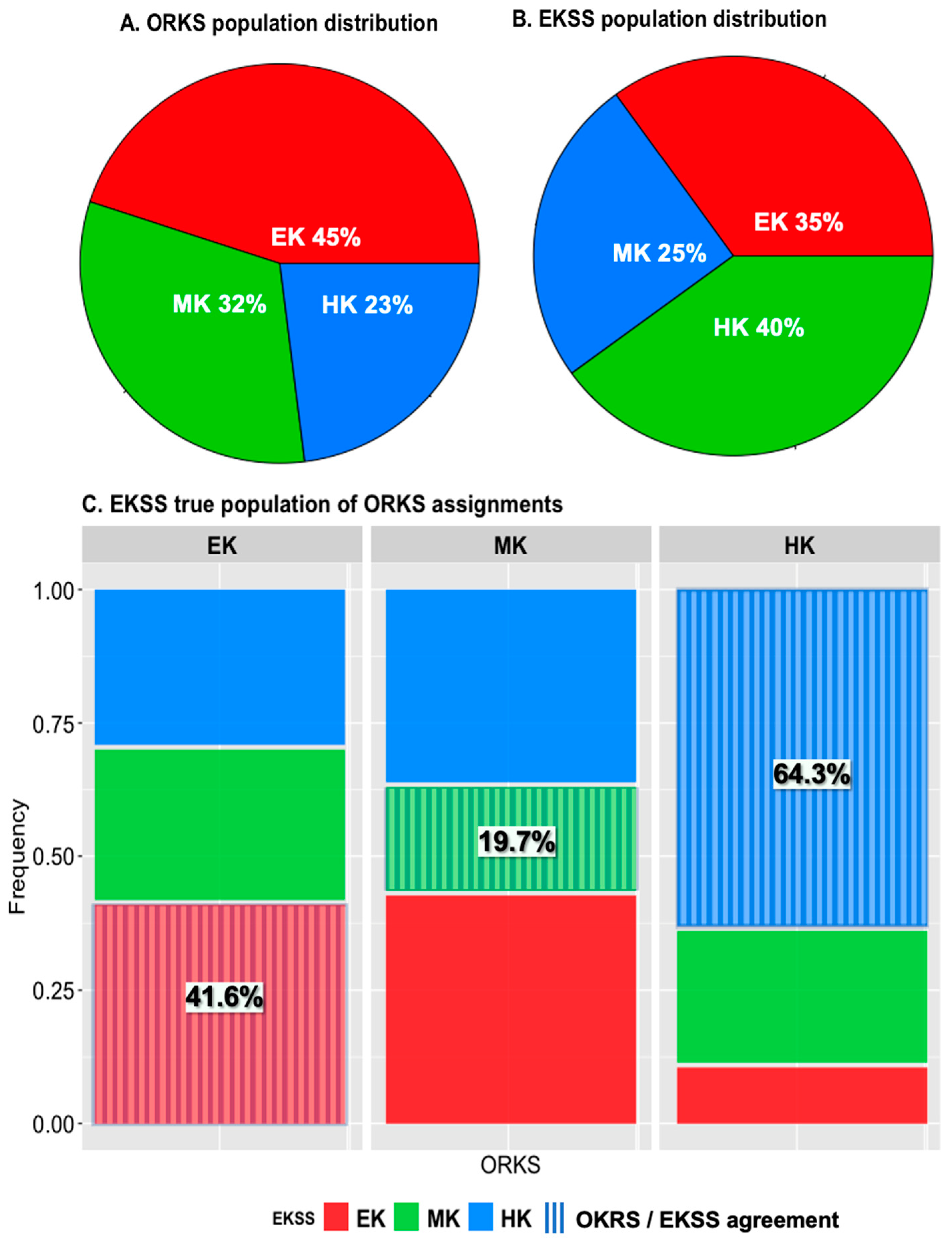
3.1.1. Keeper Status Distribution
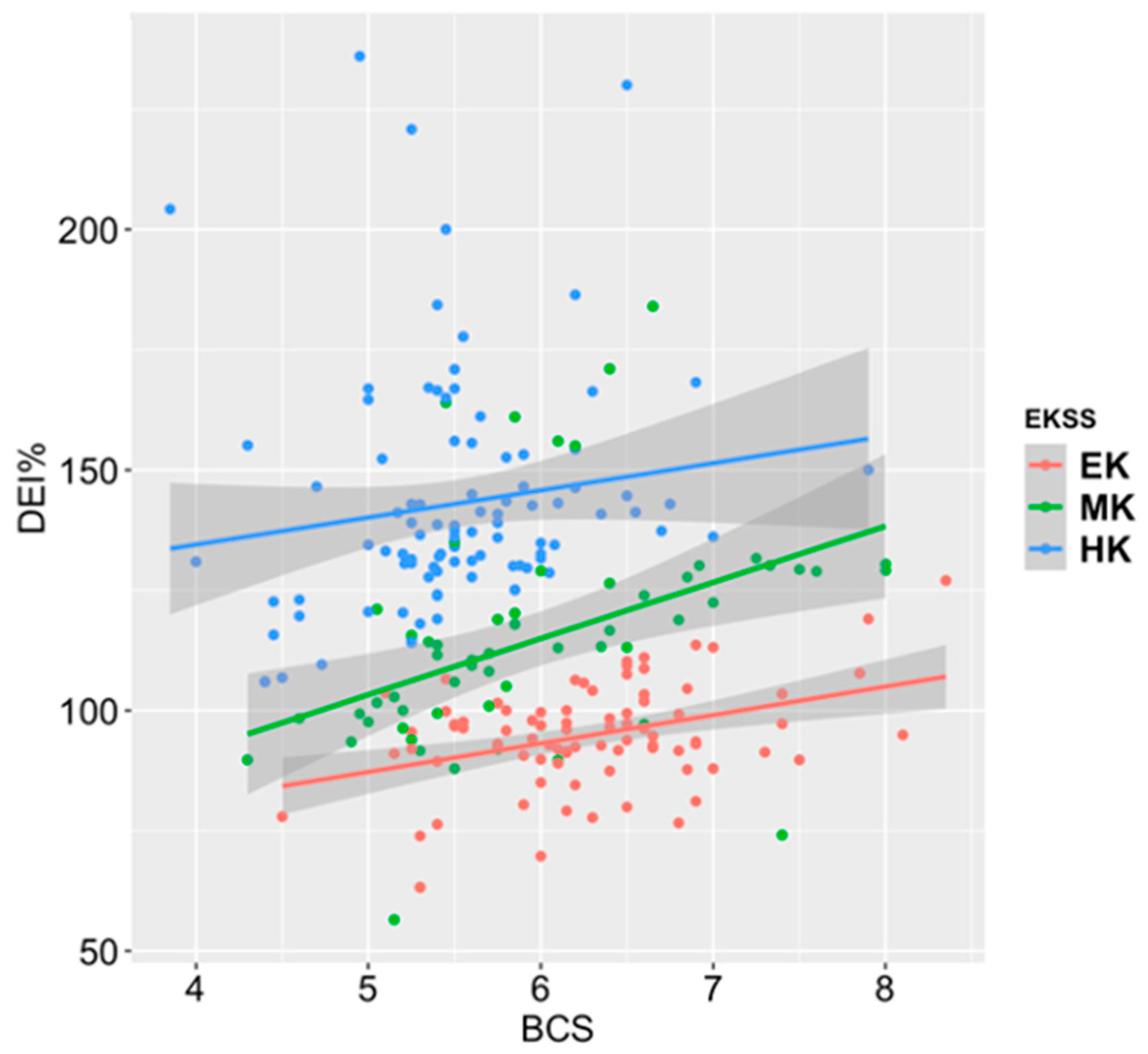
3.1.2. Equine Body Characteristics and Dietary Composition
3.2. Microbiome Survey of EKSS Horses
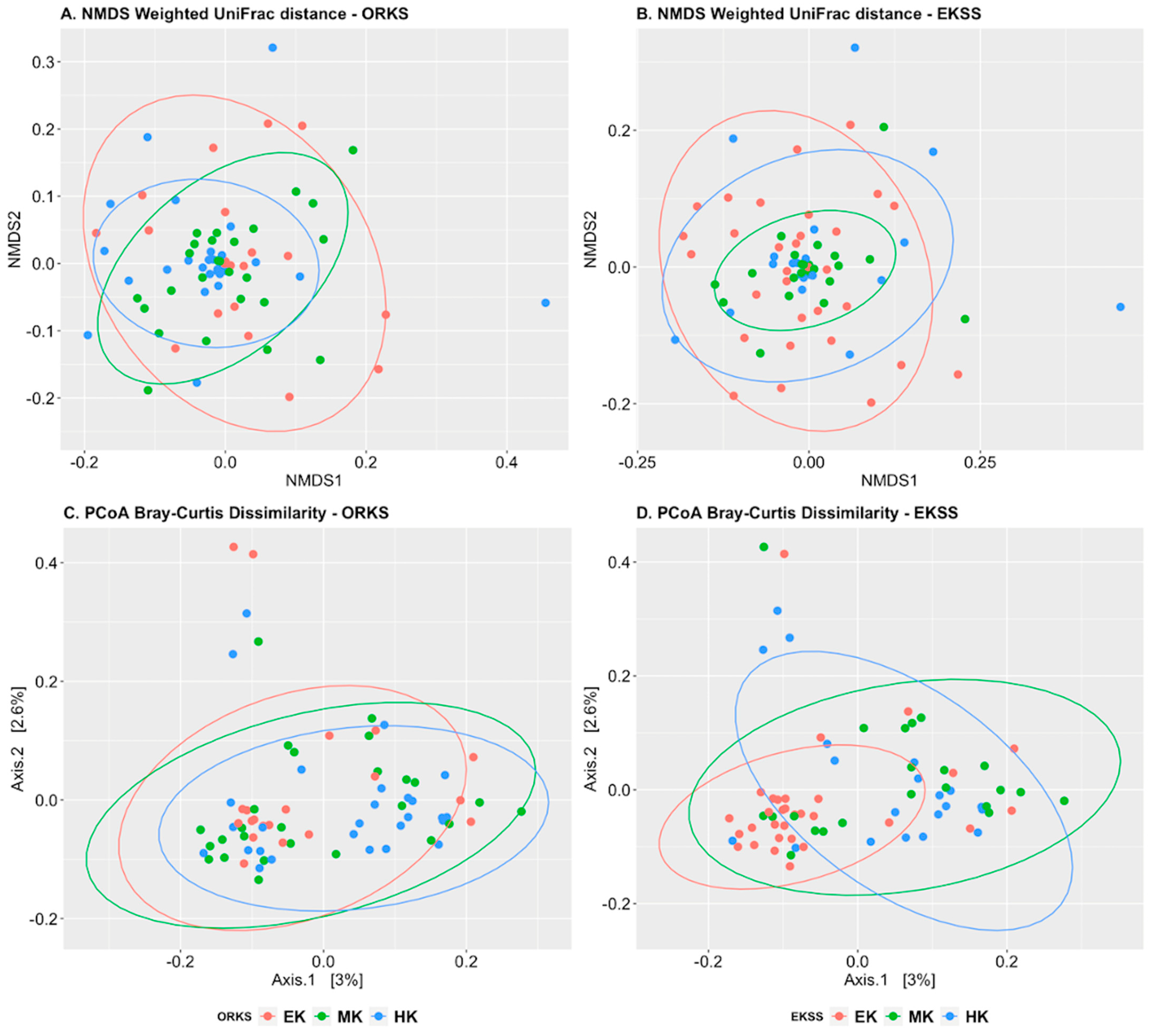
3.2.1. Alpha and Beta Diversity
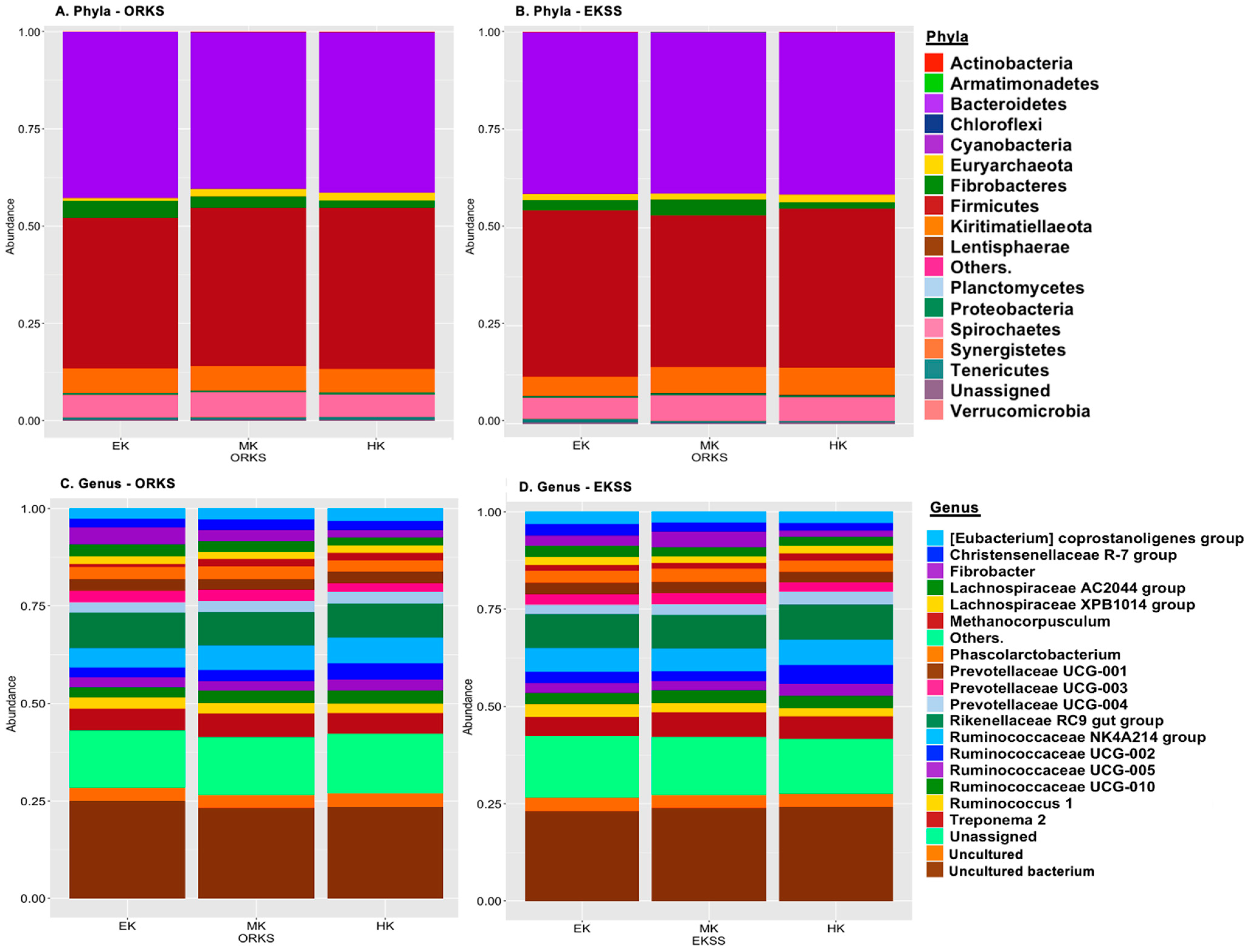
3.2.2. Relative Abundance, Differential Abundance Testing, Spearman Correlations
4. Discussion
4.1. Assigning Metabolic Tendency Using the EKSS
4.1.1. Keeper Status Distribution
4.1.2. Equine BCS and Dietary Composition
4.2. Gut Microbiome Difference Based on EKSS
4.2.1. Alpha and Beta Diversity
4.2.2. Relative Abundance, Differential Abundance Testing, Spearman Correlations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Robin, C.A.; Ireland, J.L.; Wylie, C.E.; Collins, S.N.; Verheyen, K.L.P.; Newton, J.R. Prevalence of and Risk Factors for Equine Obesity in Great Britain Based on Owner-Reported Body Condition Scores. Equine Vet. J. 2015, 47, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Henneke, D.R.; Potter, G.D.; Kreider, J.L.; Yeates, B.F. Relationship between Condition Score, Physical Measurements and Body Fat Percentage in Mares. Equine Vet. J. 1983, 15, 371–372. [Google Scholar] [CrossRef] [PubMed]
- Bäckhed, F.; Ding, H.; Wang, T.; Hooper, L.V.; Koh, G.Y.; Nagy, A.; Semenkovich, C.F.; Gordon, J.I. The Gut Microbiota as an Environmental Factor That Regulates Fat Storage. Proc. Natl. Acad. Sci. USA 2004, 101, 15718–15723. [Google Scholar] [CrossRef]
- Tudela, C.V.; Boudry, C.; Stumpff, F.; Aschenbach, J.R.; Vahjen, W.; Zentek, J.; Pieper, R. Down-Regulation of Monocarboxylate Transporter 1 (MCT1) Gene Expression in the Colon of Piglets Is Linked to Bacterial Protein Fermentation and pro-Inflammatory Cytokine-Mediated Signalling. Br. J. Nutr. 2015, 113, 610–617. [Google Scholar] [CrossRef]
- McGowan, T.W.; Pinchbeck, G.; Phillips, C.J.C.; Perkins, N.; Hodgson, D.R.; McGowan, C.M. A Survey of Aged Horses in Queensland, Australia. Part 2: Clinical Signs and Owners’ Perceptions of Health and Welfare. Aust. Vet. J. 2010, 88, 465–471. [Google Scholar] [CrossRef]
- Yngvesson, J.; Rey Torres, J.C.; Lindholm, J.; Pättiniemi, A.; Andersson, P.; Sassner, H. Health and Body Conditions of Riding School Horses Housed in Groups or Kept in Conventional Tie-Stall/Box Housing. Animals 2019, 9, 73. [Google Scholar] [CrossRef] [PubMed]
- German, A.J. The Growing Problem of Obesity in Dogs and Cats. J. Nutr. 2006, 136, 1940S–1946S. [Google Scholar] [CrossRef]
- Kienzle, E.; Bergler, R.; Mandernach, A. A Comparison of the Feeding Behavior and the Human–Animal Relationship in Owners of Normal and Obese Dogs. J. Nutr. 1998, 128, 2779S–2782S. [Google Scholar] [CrossRef]
- Argo, C.M.; Curtis, G.C.; Grove-White, D.; Dugdale, A.H.A.; Barfoot, C.F.; Harris, P.A. Weight Loss Resistance: A Further Consideration for the Nutritional Management of Obese Equidae. Vet. J. 2012, 194, 179–188. [Google Scholar] [CrossRef]
- Morrison, P.K.; Newbold, C.J.; Jones, E.; Worgan, H.J.; Grove-White, D.H.; Dugdale, A.H.; Barfoot, C.; Harris, P.A.; Argo, C.M. The Equine Gastrointestinal Microbiome: Impacts of Weight-Loss. BMC Vet. Res. 2020, 16, 78. [Google Scholar] [CrossRef]
- Dougal, K.; de la Fuente, G.; Harris, P.A.; Girdwood, S.E.; Pinloche, E.; Geor, R.J.; Nielsen, B.D.; Schott, H.C.; Elzinga, S.; Newbold, C.J. Characterisation of the Faecal Bacterial Community in Adult and Elderly Horses Fed a High Fibre, High Oil or High Starch Diet Using 454 Pyrosequencing. PLoS ONE 2014, 9, e87424. [Google Scholar] [CrossRef]
- Argenzio, R.A.; Southworth, M.; Stevens, C.E. Sites of Organic Acid Production and Absorption in the Equine Gastrointestinal Tract. Am. J. Physiol. 1974, 226, 1043–1050. [Google Scholar] [CrossRef]
- Hintz, H.F.; Argenzio, R.A.; Schryver, H.F. Digestion Coefficients, Blood Glucose Levels and Molar Percentage of Volatile Acids in Intestinal Fluid of Ponies Fed Varying Forage-Grain Ratios. J. Anim. Sci. 1971, 33, 992–995. [Google Scholar] [CrossRef] [PubMed]
- Daly, K.; Proudman, C.J.; Duncan, S.H.; Flint, H.J.; Dyer, J.; Shirazi-Beechey, S.P. Alterations in Microbiota and Fermentation Products in Equine Large Intestine in Response to Dietary Variation and Intestinal Disease. Br. J. Nutr. 2012, 107, 989–995. [Google Scholar] [CrossRef] [PubMed]
- De Fombelle, A.; Julliand, V.; Drogoul, C.; Jacotot, E. Feeding and Microbial Disorders in Horses: 1-Effects of an Abrupt Incorporation of Two Levels of Barley in a Hay Diet on Microbial Profile and Activities. J. Equine Vet. Sci. 2001, 21, 439–445. [Google Scholar] [CrossRef]
- De Fombelle, A.; Varloud, M.; Goachet, A.-G.; Jacotot, E.; Philippeau, C.; Drogoul, C.; Julliand, V. Characterization of the Microbial and Biochemical Profile of the Different Segments of the Digestive Tract in Horses given Two Distinct Diets. Anim. Sci. 2003, 77, 293–304. [Google Scholar] [CrossRef]
- Muhonen, S.; Julliand, V.; Lindberg, J.E.; Bertilsson, J.; Jansson, A. Effects on the Equine Colon Ecosystem of Grass Silage and Haylage Diets after an Abrupt Change from Hay. J. Anim. Sci. 2009, 87, 2291–2298. [Google Scholar] [CrossRef]
- Elzinga, S.E.; Weese, J.S.; Adams, A.A. Comparison of the Fecal Microbiota in Horses With Equine Metabolic Syndrome and Metabolically Normal Controls Fed a Similar All-Forage Diet. J. Equine Vet. Sci. 2016, 44, 9–16. [Google Scholar] [CrossRef]
- Louis, P.; Scott, K.P.; Duncan, S.H.; Flint, H.J. Understanding the Effects of Diet on Bacterial Metabolism in the Large Intestine. J. Appl. Microbiol. 2007, 102, 1197–1208. [Google Scholar] [CrossRef]
- Shirazi-Beechey, S.P. Molecular Insights into Dietary Induced Colic in the Horse. Equine Vet. J. 2008, 40, 414–421. [Google Scholar] [CrossRef]
- Respondek, F.; Goachet, A.G.; Julliand, V. Effects of Dietary Short-Chain Fructooligosaccharides on the Intestinal Microflora of Horses Subjected to a Sudden Change in Diet. J. Anim. Sci. 2008, 86, 316–323. [Google Scholar] [CrossRef]
- Willette, J.A.; Pitta, D.; Indugu, N.; Vecchiarelli, B.; Hennessy, M.L.; Dobbie, T.; Southwood, L.L. Experimental Crossover Study on the Effects of Withholding Feed for 24 h on the Equine Faecal Bacterial Microbiota in Healthy Mares. BMC Vet. Res. 2021, 17, 3. [Google Scholar] [CrossRef]
- Park, K.-D.; Park, J.; Ko, J.; Kim, B.C.; Kim, H.-S.; Ahn, K.; Do, K.-T.; Choi, H.; Kim, H.-M.; Song, S.; et al. Whole Transcriptome Analyses of Six Thoroughbred Horses before and after Exercise Using RNA-Seq. BMC Genom. 2012, 13, 473. [Google Scholar] [CrossRef]
- Willing, B.; Vörös, A.; Roos, S.; Jones, C.; Jansson, A.; Lindberg, J.E. Changes in Faecal Bacteria Associated with Concentrate and Forage-Only Diets Fed to Horses in Training. Equine Vet. J. 2009, 41, 908–914. [Google Scholar] [CrossRef]
- Blackmore, T.M.; Dugdale, A.; Argo, C.M.; Curtis, G.; Pinloche, E.; Harris, P.A.; Worgan, H.J.; Girdwood, S.E.; Dougal, K.; Newbold, C.J.; et al. Strong Stability and Host Specific Bacterial Community in Faeces of Ponies. PLoS ONE 2013, 8, e75079. [Google Scholar] [CrossRef] [PubMed]
- Wyse, C.A.; McNie, K.A.; Tannahil, V.J.; Love, S.; Murray, J.K. Prevalence of Obesity in Riding Horses in Scotland. Vet. Rec. 2008, 162, 590–591. [Google Scholar] [CrossRef]
- National Research Council (U.S.) (Ed.) Nutrient Requirements of Horses, 6th ed.; Animal Nutrition Series; National Academies Press: Washington, DC, USA, 2007; ISBN 978-0-309-10212-4. [Google Scholar]
- Gehlen, H.; Schwarz, B.; Bartmann, C.; Gernhardt, J.; Stöckle, S.D. Pituitary Pars Intermedia Dysfunction and Metabolic Syndrome in Donkeys. Animals 2020, 10, 2335. [Google Scholar] [CrossRef]
- Kohnke, J. Feeding and Nutrition: The Making of a Champion; Birubi Pacific: Pymble, Australia, 1992; pp. 163–166. [Google Scholar]
- Wagner, E.L.; Tyler, P.J. A Comparison of Weight Estimation Methods in Adult Horses. J. Equine Vet. Sci. 2011, 31, 706–710. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundatioon for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Dugdale, A.H.A.; Grove-White, D.; Curtis, G.C.; Harris, P.A.; Argo, C.M. Body Condition Scoring as a Predictor of Body Fat in Horses and Ponies. Vet. J. 2012, 194, 173–178. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science Using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Package ‘Vegan’, Community Ecology Package; R Package Version 2.5-6; 2019. Available online: https://CRAN.R-project.org/package=vegan (accessed on 23 March 2021).
- Stevens, A.J.; Purcell, R.V.; Darling, K.A.; Eggleston, M.J.F.; Kennedy, M.A.; Rucklidge, J.J. Human Gut Microbiome Changes during a 10 Week Randomised Control Trial for Micronutrient Supplementation in Children with Attention Deficit Hyperactivity Disorder. Sci. Rep. 2019, 9, 10128. [Google Scholar] [CrossRef] [PubMed]
- Ragnarsson, S.; Jansson, A. Comparison of Grass Haylage Digestibility and Metabolic Plasma Profile in Icelandic and Standardbred Horses. J. Anim. Physiol. Anim. Nutr. 2011, 95, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Giles, S.L.; Rands, S.A.; Nicol, C.J.; Harris, P.A. Obesity Prevalence and Associated Risk Factors in Outdoor Living Domestic Horses and Ponies. PeerJ 2014, 2, e299. [Google Scholar] [CrossRef]
- Morrison, P.K.; Harris, P.A.; Maltin, C.A.; Grove-White, D.; Barfoot, C.F.; Argo, C.M. Perceptions of Obesity and Management Practices in a UK Population of Leisure-Horse Owners and Managers. J. Equine Vet. Sci. 2017, 53, 19–29. [Google Scholar] [CrossRef]
- Obesity and the Show Ring—Horse & Hound. Available online: https://www.horseandhound.co.uk/horse-care/feeding/obesity-and-the-show-ring-62276 (accessed on 23 March 2021).
- Owers, R.; Chubbock, S. Fight the Fat! Equine Vet. J. 2013, 45, 5. [Google Scholar] [CrossRef]
- Thatcher, C.D.; Pleasant, R.S.; Geor, R.J.; Elvinger, F. Prevalence of Overconditioning in Mature Horses in Southwest Virginia during the Summer. J. Vet. Intern. Med. 2012, 26, 1413–1418. [Google Scholar] [CrossRef]
- Pratt-Phillips, S.E.; Owens, K.M.; Dowler, L.E.; Cloninger, M.T. Assessment of Resting Insulin and Leptin Concentrations and Their Association With Managerial and Innate Factors in Horses. J. Equine Vet. Sci. 2010, 30, 127–133. [Google Scholar] [CrossRef]
- Robie, S.; Janson, C.; Smith, S.; O’Connor, J., Jr. Equine Serum Lipids: Serum Lipids and Glucose in Morgan and Thoroughbred Horses and Shetland Ponies. Am. J. Vet. Res. 1975, 36, 1705–1708. [Google Scholar]
- Johnson, P.J. The Equine Metabolic Syndrome: Peripheral Cushing’s Syndrome. Vet. Clin. N. Am. Equine Pract. 2002, 18, 271–293. [Google Scholar] [CrossRef]
- Dougal, K.; Harris, P.A.; Edwards, A.; Pachebat, J.A.; Blackmore, T.M.; Worgan, H.J.; Newbold, C.J. A Comparison of the Microbiome and the Metabolome of Different Regions of the Equine Hindgut. FEMS Microbiol. Ecol. 2012, 82, 642–652. [Google Scholar] [CrossRef]
- Costa, M.C.; Arroyo, L.G.; Allen-Vercoe, E.; Stämpfli, H.R.; Kim, P.T.; Sturgeon, A.; Weese, J.S. Comparison of the Fecal Microbiota of Healthy Horses and Horses with Colitis by High Throughput Sequencing of the V3-V5 Region of the 16S RRNA Gene. PLoS ONE 2012, 7, e41484. [Google Scholar] [CrossRef] [PubMed]
- Hansen, N.C.K.; Avershina, E.; Mydland, L.T.; Næsset, J.A.; Austbø, D.; Moen, B.; Måge, I.; Rudi, K. High Nutrient Availability Reduces the Diversity and Stability of the Equine Caecal Microbiota. Microb. Ecol. Health Dis. 2015, 26. [Google Scholar] [CrossRef]
- Stewart, H.L.; Southwood, L.L.; Indugu, N.; Vecchiarelli, B.; Engiles, J.B.; Pitta, D. Differences in the Equine Faecal Microbiota between Horses Presenting to a Tertiary Referral Hospital for Colic Compared with an Elective Surgical Procedure. Equine Vet. J. 2019, 51, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Dougal, K.; de la Fuente, G.; Harris, P.A.; Girdwood, S.E.; Pinloche, E.; Newbold, C.J. Identification of a Core Bacterial Community within the Large Intestine of the Horse. PLoS ONE 2013, 8, e77660. [Google Scholar] [CrossRef] [PubMed]
- Biddle, A.S.; Tomb, J.-F.; Fan, Z. Microbiome and Blood Analyte Differences Point to Community and Metabolic Signatures in Lean and Obese Horses. Front. Vet. Sci. 2018, 5, 225. [Google Scholar] [CrossRef]
- Morrison, P.K.; Newbold, C.J.; Jones, E.; Worgan, H.J.; Grove-White, D.H.; Dugdale, A.H.; Barfoot, C.; Harris, P.A.; Argo, C.M. The Equine Gastrointestinal Microbiome: Impacts of Age and Obesity. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Kaboré, O.D.; Godreuil, S.; Drancourt, M. Planctomycetes as Host-Associated Bacteria: A Perspective That Holds Promise for Their Future Isolations, by Mimicking Their Native Environmental Niches in Clinical Microbiology Laboratories. Front. Cell. Infect. Microbiol. 2020, 10. [Google Scholar] [CrossRef]
- Kim, H.M.; Jung, J.Y.; Yergeau, E.; Hwang, C.Y.; Hinzman, L.; Nam, S.; Hong, S.G.; Kim, O.-S.; Chun, J.; Lee, Y.K. Bacterial Community Structure and Soil Properties of a Subarctic Tundra Soil in Council, Alaska. FEMS Microbiol. Ecol. 2014, 89, 465–475. [Google Scholar] [CrossRef]
- Miyashita, N.T. Contrasting Soil Bacterial Community Structure between the Phyla Acidobacteria and Proteobacteria in Tropical Southeast Asian and Temperate Japanese Forests. Genes Genet. Syst. 2015, 90, 61–77. [Google Scholar] [CrossRef][Green Version]
- Delmont, T.O.; Quince, C.; Shaiber, A.; Esen, Ö.C.; Lee, S.T.; Rappé, M.S.; McLellan, S.L.; Lücker, S.; Eren, A.M. Nitrogen-Fixing Populations of Planctomycetes and Proteobacteria Are Abundant in Surface Ocean Metagenomes. Nat. Microbiol. 2018, 3, 804–813. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An Obesity-Associated Gut Microbiome with Increased Capacity for Energy Harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef] [PubMed]
- Webb, P.; Annis, J. Adaptation to Overeating in Lean and Overweight Men and Women. Hum. Nutr. Clin. Nutr. 1983, 37, 117–131. [Google Scholar]
- Janssen, P.H.; Kirs, M. Structure of the Archaeal Community of the Rumen. Appl. Environ. Microbiol. 2008, 74, 3619–3625. [Google Scholar] [CrossRef]
- Newbold, C.J.; Ramos-Morales, E. Review: Ruminal Microbiome and Microbial Metabolome: Effects of Diet and Ruminant Host. Animal 2020, 14, s78–s86. [Google Scholar] [CrossRef]
- Edwards, J.E.; Shetty, S.A.; van den Berg, P.; Burden, F.; van Doorn, D.A.; Pellikaan, W.F.; Dijkstra, J.; Smidt, H. Multi-Kingdom Characterization of the Core Equine Fecal Microbiota Based on Multiple Equine (Sub)Species. Anim. Microbiome 2020, 2, 6. [Google Scholar] [CrossRef] [PubMed]
- Jensen, B.B. Methanogenesis in Monogastric Animals. Environ. Monit. Assess. 1996, 42, 99–112. [Google Scholar] [CrossRef] [PubMed]
- Ishaq, S.L.; Sundset, M.A.; Crouse, J.; Wright, A.-D.G. High-Throughput DNA Sequencing of the Moose Rumen from Different Geographical Locations Reveals a Core Ruminal Methanogenic Archaeal Diversity and a Differential Ciliate Protozoal Diversity. Microb. Genom. 2015, 1. [Google Scholar] [CrossRef] [PubMed]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut Microbiota Functions: Metabolism of Nutrients and Other Food Components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Tokuda, G.; Mikaelyan, A.; Fukui, C.; Matsuura, Y.; Watanabe, H.; Fujishima, M.; Brune, A. Fiber-Associated Spirochetes Are Major Agents of Hemicellulose Degradation in the Hindgut of Wood-Feeding Higher Termites. Proc. Natl. Acad. Sci. USA 2018, 115, E11996–E12004. [Google Scholar] [CrossRef]
- Tsoy, O.V.; Ravcheev, D.A.; Čuklina, J.; Gelfand, M.S. Nitrogen Fixation and Molecular Oxygen: Comparative Genomic Reconstruction of Transcription Regulation in Alphaproteobacteria. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef]
- Spain, A.M.; Krumholz, L.R.; Elshahed, M.S. Abundance, Composition, Diversity and Novelty of Soil Proteobacteria. ISME J. 2009, 3, 992–1000. [Google Scholar] [CrossRef]
- Kauter, A.; Epping, L.; Semmler, T.; Antao, E.-M.; Kannapin, D.; Stoeckle, S.D.; Gehlen, H.; Lübke-Becker, A.; Günther, S.; Wieler, L.H.; et al. The Gut Microbiome of Horses: Current Research on Equine Enteral Microbiota and Future Perspectives. Anim. Microbiome 2019, 1, 14. [Google Scholar] [CrossRef] [PubMed]
- Scales, B.S.; Dickson, R.P.; Huffnagle, G.B. A Tale of Two Sites: How Inflammation Can Reshape the Microbiomes of the Gut and Lungs. J. Leukoc. Biol. 2016, 100, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Winter, S.E.; Bäumler, A.J. Dysbiosis in the Inflamed Intestine. Gut Microbes 2014, 5, 71–73. [Google Scholar] [CrossRef]
- Winter, S.E.; Bäumler, A.J. Why Related Bacterial Species Bloom Simultaneously in the Gut: Principles Underlying the ‘Like Will to like’ Concept. Cell. Microbiol. 2014, 16, 179–184. [Google Scholar] [CrossRef]
- Al Jassim, R.A.M.; Andrews, F.M. The Bacterial Community of the Horse Gastrointestinal Tract and Its Relation to Fermentative Acidosis, Laminitis, Colic, and Stomach Ulcers. Vet. Clin. N. Am. Equine Pract. 2009, 25, 199–215. [Google Scholar] [CrossRef] [PubMed]
- Petri, R.M.; Schwaiger, T.; Penner, G.B.; Beauchemin, K.A.; Forster, R.J.; McKinnon, J.J.; McAllister, T.A. Changes in the Rumen Epimural Bacterial Diversity of Beef Cattle as Affected by Diet and Induced Ruminal Acidosis. Appl. Environ. Microbiol. 2013, 79, 3744–3755. [Google Scholar] [CrossRef]
- O’Herrin, S.M.; Kenealy, W.R. Glucose and Carbon Dioxide Metabolism by Succinivibrio Dextrinosolvens. Appl. Environ. Microbiol. 1993, 59, 748–755. [Google Scholar] [CrossRef]
- Mattila, H.R.; Rios, D.; Walker-Sperling, V.E.; Roeselers, G.; Newton, I.L.G. Characterization of the Active Microbiotas Associated with Honey Bees Reveals Healthier and Broader Communities When Colonies Are Genetically Diverse. PLoS ONE 2012, 7, e32962. [Google Scholar] [CrossRef]
- Kobayashi, Y. Inclusion of Novel Bacteria in Rumen Microbiology: Need for Basic and Applied Science. Anim. Sci. J. 2006, 77, 375–385. [Google Scholar] [CrossRef]
- Hess, M.; Sczyrba, A.; Egan, R.; Kim, T.-W.; Chokhawala, H.; Schroth, G.; Luo, S.; Clark, D.S.; Chen, F.; Zhang, T.; et al. Metagenomic Discovery of Biomass-Degrading Genes and Genomes from Cow Rumen. Science 2011, 331, 463–467. [Google Scholar] [CrossRef] [PubMed]
- Taxis, T.M.; Wolff, S.; Gregg, S.J.; Minton, N.O.; Zhang, C.; Dai, J.; Schnabel, R.D.; Taylor, J.F.; Kerley, M.S.; Pires, J.C.; et al. The Players May Change but the Game Remains: Network Analyses of Ruminal Microbiomes Suggest Taxonomic Differences Mask Functional Similarity. Nucleic Acids Res. 2015, 43, 9600–9612. [Google Scholar] [CrossRef]
- Salis, R.K.; Bruder, A.; Piggott, J.J.; Summerfield, T.C.; Matthaei, C.D. High-Throughput Amplicon Sequencing and Stream Benthic Bacteria: Identifying the Best Taxonomic Level for Multiple-Stressor Research. Sci. Rep. 2017, 7, 44657. [Google Scholar] [CrossRef] [PubMed]
- Solomon, R.; Wein, T.; Levy, B.; Reiss, V.; Zehavi, T.; Furman, O.; Jami, E. Rumen Protozoa Shape Microbiome Composition and Metabolic Output of Fermentation. bioRxiv 2020. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EKSS Horses (n = 240) | Fecal Sampling (n = 73) | |
|---|---|---|
| EKSS Group 1 | ||
| EK | 35 (84) | 42 (31) |
| MK | 25 (60) | 30 (22) |
| HK | 40 (96) | 27 (20) |
| EKSS Level 2 | ||
| E+ | 24 (57) | 30 (22) |
| E | 11 (27) | 12 (9) |
| M+ | 9 (22) | 11 (8) |
| M | 9 (21) | 11 (8) |
| M- | 7 (17) | 8 (6) |
| H | 6 (14) | 7 (5) |
| H- | 34 (82) | 21 (15) |
| BCS Rank 3 | ||
| Lean | 8 (18) | 7 (5) |
| Normal | 53 (128) | 62 (45) |
| Obese | 39 (94) | 32 (23) |
| EKSS and ORKS Keeper Agreement 4 | ||
| Yes | 40 (95) | 47 (34) |
| No | 60 (145) | 53 (39) |
| EK | MK | HK | p-Value | |
|---|---|---|---|---|
| BCS | 6.31 a (0.70) | 5.95 b (0.84) | 5.52 c (0.65) | <0.001 |
| eTBF% 1 | 9.84 a (1.09) | 9.29 b (1.31) | 8.61 c (1.01) | <0.001 |
| Forage intake, kg | 8.05 a (1.79) | 8.19 a (2.45) | 8.15 a (2.62) | <0.001 |
| Grain intake, kg | 1.63 c (1.36) | 2.38 b (1.87) | 3.36 a (2.96) | <0.001 |
| Total intake, % 2 | 2.05 b (0.19) | 2.02 b (0.23) | 2.33 a (0.53) | <0.001 |
| DEI, % 3 | 94.92 c (10.66) | 114.39 b (23.76) | 143.03 a (24.49) | <0.001 |
| CPI, % 4 | 167.91 c (35.24) | 197.34 b (33.68) | 225.96 a (33.20) | <0.001 |
| ORKS | EKSS | |||||||
|---|---|---|---|---|---|---|---|---|
| EK | MK | HK | p-Value | EK | MK | HK | p-Value | |
| Shannon | 5.28 a | 5.43 a | 5.50 a | 0.34 | 5.30 a | 5.62 b | 5.37 ab | 0.06 |
| Observed sp. | 351 a | 390 a | 432 a | 0.28 | 353 a | 469 b | 378 ab | 0.04 |
| EKSS Abundance | ANCOM | ||||
|---|---|---|---|---|---|
| EK | MK | HK | W 1 | clr 2 | |
| Planctomycetes 3 | 0.337 | 0.002 | 0.098 | 3 | 5.22 |
| Euryarchaeota 3 | 1.611 | 1.452 | 2.020 | 1 | 3.27 |
| Spirochaetes 3 | 4.595 | 0.055 | 5.402 | 1 | 2.88 |
| Proteobacteria 3 | 0.646 | 1.088 | 0.960 | 1 | 2.55 |
| Gammaproteobacteria 4 | 0.063 | 0.001 | 0.272 | 18 | 8.44 |
| EKSS | |||
|---|---|---|---|
| EK | MK | HK | |
| Phyla | |||
| Planctomycetes | 0.33 | −0.30 | |
| Proteobacteria | −0.30 | ||
| Class | |||
| Gammaproteobacteria 2 | −0.36 | ||
| Bacteroidia 3 | −0.32 | ||
| Order | |||
| Rickettsiales 2 | 0.32 | ||
| Aeromondales 2 | 0.36 | ||
| Izimaplasmatales 4 | 0.41 | ||
| Clostridiales 5 | 0.41 | −0.31 | |
| Family | |||
| Methanomethylophilaceae 1 | −0.32 | ||
| Succinivibrionaceae 2 | 0.36 | ||
| Burkholderiaceae 2 | −0.31 | ||
| Bacteroidales RF16 group 3 | −0.31 | ||
| Bacteroidales UCG 001 3 | 0.39 | −0.32 | |
| Muribaculaceae 3 | 0.31 | ||
| Mycoplasmataceae 4 | −0.32 | 0.30 | |
| Peptococcaceae 5 | 0.31 | −0.34 | |
| Gastranaerophilales, uncultured rumen bacterium 6 | 0.32 | ||
| Genera | |||
| Candidatus methanomethylophilus1 | −0.33 | ||
| Succinivibrionaceae, uncultured2 | −0.31 | 0.32 | |
| Bacteroidia3 | −0.32 | ||
| Bacteroidales, F0823 | −0.33 | ||
| Clostridiales; f_; g_5 | 0.41 | −0.31 | |
| Eubacterium oxidoreducens group5 | 0.39 | ||
| Lachnospiraceae UCG 0085 | −0.38 | ||
| Marvinbryantia5 | −0.31 | ||
| Ruminococcaceae NK4A214 group5 | −0.30 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johnson, A.C.B.; Biddle, A.S. A Standard Scale to Measure Equine Keeper Status and the Effect of Metabolic Tendency on Gut Microbiome Structure. Animals 2021, 11, 1975. https://doi.org/10.3390/ani11071975
Johnson ACB, Biddle AS. A Standard Scale to Measure Equine Keeper Status and the Effect of Metabolic Tendency on Gut Microbiome Structure. Animals. 2021; 11(7):1975. https://doi.org/10.3390/ani11071975
Chicago/Turabian StyleJohnson, Alexa C. B., and Amy S. Biddle. 2021. "A Standard Scale to Measure Equine Keeper Status and the Effect of Metabolic Tendency on Gut Microbiome Structure" Animals 11, no. 7: 1975. https://doi.org/10.3390/ani11071975
APA StyleJohnson, A. C. B., & Biddle, A. S. (2021). A Standard Scale to Measure Equine Keeper Status and the Effect of Metabolic Tendency on Gut Microbiome Structure. Animals, 11(7), 1975. https://doi.org/10.3390/ani11071975