
Antimicrobial Peptides from Black Soldier Fly (Hermetia illucens) as Potential Antimicrobial Factors Representing an Alternative to Antibiotics in Livestock Farming



Abstract
:Simple Summary
Abstract
1. Introduction
2. Medicinal Value of Antimicrobial Peptides
3. Diversity of Antimicrobial Peptides in Insects
4. Mechanism of Action of Antimicrobial Peptides
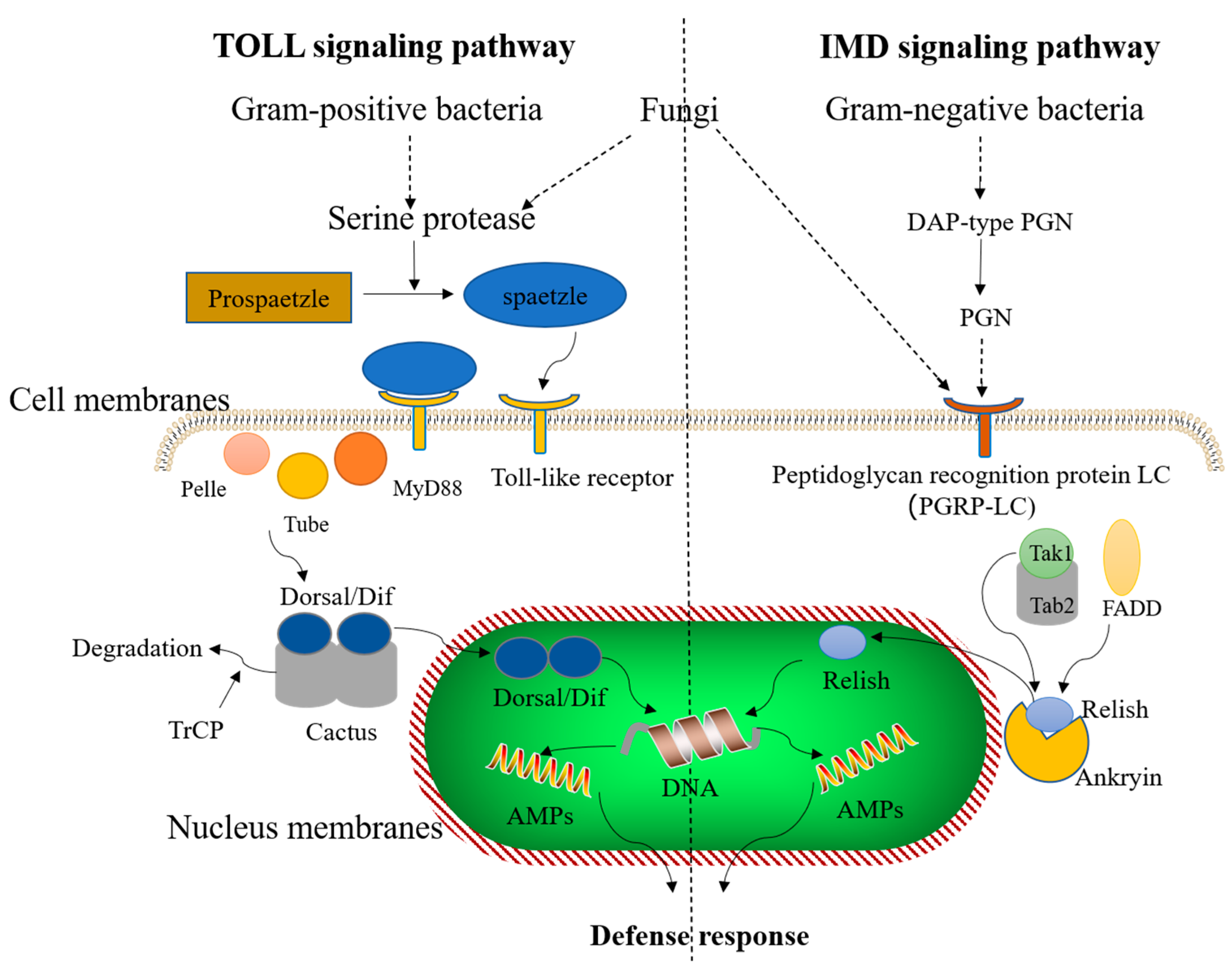
5. AMP Induction of Immune Signaling Pathways in Insects
6. Bacterial Resistance to Insect AMPs
7. Application of BSF-Derived AMPs in Livestock Production
8. Conclusions and Prospects
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Kim, S.W.; Less, J.F.; Wang, L.; Yan, T.; Kiron, V.; Kaushik, S.J.; Lei, X.G. Meeting Global Feed Protein Demand: Challenge, Opportunity, and Strategy. Annu. Rev. Anim. Biosci. 2019, 7, 17.01–17.23. [Google Scholar] [CrossRef]
- Nguyen, N.; Nguyen, C.; Guy, T.; Juan, C.M. Antimicrobial Usage and Antimicrobial Resistance in Animal Production in Southeast Asia: A Review. Antibiotics 2016, 5, 37. [Google Scholar] [CrossRef] [Green Version]
- Thakur, S.D.; Panda, A. Rational Use of Antimicrobials in Animal Production:A Prerequisite to Stem the Tide of Antimicrobial Resistance. Curr. Sci. 2017, 113, 1846–1857. [Google Scholar] [CrossRef]
- López-Gálvez, G.; López-Alonso, M.; Pechova, A.; Mayo, B.; Dierick, N.; Gropp, J. Alternatives to antibiotics and trace elements (copper and zinc) to improve gut health and zootechnical parameters in piglets: A review. Anim. Feed. Sci. Technol. 2020, 114727. [Google Scholar] [CrossRef]
- Jones, H.E.; O’Connell White, K.; Norman, W.V.; Guilbert, E.; Lichtenberg, E.S.; Paul, M. First trimester medication abortion practice in the United States and Canada. PLoS ONE 2017, 12, e0186487. [Google Scholar] [CrossRef] [Green Version]
- Phillips, I. Withdrawal of growth-promoting antibiotics in Europe and its effects in relation to human health. Int. J. Antimicrob. Agents 2007, 30, 101–107. [Google Scholar] [CrossRef]
- Bedford, M. Removal of antibiotic growth promoters from poultry diets: Implications and strategies to minimise subsequent problems. World’s Poult. Sci. J. 2000, 56, 347–365. [Google Scholar] [CrossRef]
- Flynn, W.T. The Judicious Use of Medically Important Antimicrobial Drugs in Food-Producing Animals; Center for Veterinary Medicine (HFV-1), Food and Drug Administration, US Department of Health and Human Services: Washington, DC, USA, 2012. [Google Scholar]
- El-Hack, A.; Mohamed, E.; Alagawany, M.; Shaheen, H.; Samak, D.; Othman, S.I.; Allam, A.A.; Taha, A.E.; Khafaga, A.F.; Arif, M. Ginger and Its Derivatives as Promising Alternatives to Antibiotics in Poultry Feed. Animals 2020, 10, 452. [Google Scholar] [CrossRef] [Green Version]
- Agnolucci, M.; Daghio, M.; Mannelli, F.; Secci, G.; Buccioni, A. Use of chitosan and tannins as alternatives to antibiotics to control mold growth on PDO Pecorino Toscano cheese rind. Food Microbiol. 2020, 92, 103598. [Google Scholar] [CrossRef] [PubMed]
- Shirzadi, H.; Shariatmadari, F.; Torshizi, M.A.K.; Rahimi, S.; Masoudi, A.A.; Zaboli, G.; Hedayat-Evrigh, N. Plant extract supplementation as a strategy for substituting dietary antibiotics in broiler chickens exposed to low ambient temperature. Arch. Anim. Nutr. 2020, 74, 206–221. [Google Scholar] [CrossRef]
- Makkar, H.P.S. Review: Feed demand landscape and implications of food-not feed strategy for food security and climate change. Animal 2018, 12, 1744–1754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oonincx, D.G.; van Broekhoven, S.; van Huis, A.; van Loon, J.J. Feed conversion, survival and development, and composition of four insect species on diets composed of food by-products. PLoS ONE 2019, 10, e0144601, Correction in PLoS ONE 2019, 14, e0222043. [Google Scholar] [CrossRef] [PubMed]
- Van Huis, A. Potential of Insects as Food and Feed in Assuring Food Security. Annu. Rev. Entomol. 2013, 58, 563–583. [Google Scholar] [CrossRef] [PubMed]
- Oonincx, D.G.A.B.; Joost, V.I.; Heetkamp, M.J.W.; Henry, V.D.B.; Van, L.J.J.A.; Arnold, V.H.; Hansen, I.A. An Exploration on Greenhouse Gas and Ammonia Production by Insect Species Suitable for Animal or Human Consumption. PLoS ONE 2010, 5, e14445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makkar, H.P.S.; Tran, G.; Heuzé, V.; Ankers, P. State-of-the-art on use of insects as animal feed. Anim. Feed Sci. Technol. 2014, 197, 1–33. [Google Scholar] [CrossRef]
- Wang, Y.S.; Shelomi, M. Review of Black Soldier Fly (Hermetia illucens) as Animal Feed and Human Food. Foods 2017, 6, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sogari, G.; Amato, M.; Biasato, I.; Chiesa, S.; Gasco, L. The Potential Role of Insects as Feed: A Multi-Perspective Review. Animals 2019, 9, 119. [Google Scholar] [CrossRef] [Green Version]
- Spranghers, T.; Noyez, A.; Schildermans, K.; De Clercq, P. Cold Hardiness of the Black Soldier Fly (Diptera: Stratiomyidae). J. Econ. Entomol. 2017, 110, 1501–1507. [Google Scholar] [CrossRef]
- Cickova, H.; Newton, G.L.; Lacy, R.C.; Kozanek, M. The use of fly larvae for organic waste treatment. Waste Manag. 2015, 35, 68–80. [Google Scholar] [CrossRef]
- Craig, S.D.; Tomberlin, J.K.; Joyce, J.A.; Kiser, B.C.; Sumner, S.M. Rearing Methods for the Black Soldier Fly (Diptera: Stratiomyidae). J. Med. Entomol. 2002, 39, 695–698. [Google Scholar] [CrossRef] [Green Version]
- Green, T.R.; Popa, R. Enhanced ammonia content in compost leachate processed by black soldier fly larvae. Appl. Biochem. Biotechnol. 2012, 166, 1381–1387. [Google Scholar] [CrossRef]
- Zheng, L.; Hou, Y.; Wu, L.; Yang, S.; Li, Q.; Yu, Z. Biodiesel production from rice straw and restaurant waste employing black soldier fly assisted by microbes. Energy 2012, 47, 225–229. [Google Scholar] [CrossRef]
- Liu, C.; Wang, C.; Yao, H.; Chapman, S.J. Pretreatment is an important method for increasing the conversion efficiency of rice straw by black soldier fly larvae based on the function of gut microorganisms. Sci. Total Environ. 2021, 762. [Google Scholar] [CrossRef] [PubMed]
- Banks, I.J.; Gibson, W.T.; Cameron, M.M. Growth rates of black soldier fly larvae fed on fresh human faeces and their implication for improving sanitation. Trop. Med. Int. Health 2013, 19, 14–22. [Google Scholar] [CrossRef]
- Lalander, C.; Diener, S.; Magri, M.E.; Zurbruegg, C.; Lindstroem, A.; Vinneras, B. Faecal sludge management with the larvae of the black soldier fly (Hermetia illucens)—From a hygiene aspect. Sci. Total Environ. 2013, 458, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Bae, S.; Park, K.; Lee, S.; Choi, Y.; Han, S.; Koh, Y. Biochemical characterization of digestive enzymes in the black soldier fly, Hermetia illucens (Diptera: Stratiomyidae). J. Asia-Pacif. Entomol. 2011, 14, 11–14. [Google Scholar] [CrossRef]
- Choi, W.H.; Choi, H.J.; Goo, T.W.; Quan, F.S. Novel antibacterial peptides induced by probiotics in Hermetia illucens (Diptera: Stratiomyidae) larvae. Entomol. Res. 2018, 48, 237–247. [Google Scholar] [CrossRef]
- Osama, E.; Dingzhong, Z.; Qi, S.; Aziz, S.A.; Minmin, C.; Longyu, Z.; Ziniu, Y.; Jibin, Z.; Humberto, L.M. Screening, Expression, Purification and Functional Characterization of Novel Antimicrobial Peptide Genes from Hermetia illucens (L.). PLoS ONE 2017, 12, e0169582. [Google Scholar] [CrossRef]
- Lalander, C.H.; Fidjeland, J.; Diener, S.; Eriksson, S.; Vinnerås, B. High waste-to-biomass conversion and efficient Salmonella spp. reduction using black soldier fly for waste recycling. Agron. Sus. Tain. Dev. 2015, 35, 261–271. [Google Scholar] [CrossRef]
- Feng, M.; Fei, S.; Xia, J.; Labropoulou, V.; Sun, J. Antimicrobial Peptides as Potential Antiviral Factors in Insect Antiviral Immune Response. Front. Immunol. 2020, 11, 2030. [Google Scholar] [CrossRef]
- Li, Z.; Mao, R.; Teng, D.; Hao, Y.; Chen, H.; Wang, X.; Wang, X.; Yang, N.; Wang, J. Antibacterial and immunomodulatory activities of insect defensins-DLP2 and DLP4 against multidrug-resistant Staphylococcus aureus. Sci. Rep. 2017, 7, 12124. [Google Scholar] [CrossRef] [PubMed]
- Chung, P.Y.; Khanum, R. Antimicrobial peptides as potential anti-biofilm agents against multidrug-resistant bacteria. J. Microbiol. Immunol. Infect. 2017, 50, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Broeck, J.V.; Schoofs, L.; De Loof, A. Insect neuropeptides and their receptors: New leads for medical and agricultural applications. Trends Endocrinol. Metab. 1997, 8, 321–326. [Google Scholar] [CrossRef]
- Mlcek, J.; Borkovcova, M.; Rop, O.; Bednarova, M. Biologically active substances of edible insects and their use in agriculture, veterinary and human medicine. JCEA 2014, 15, 225–237. [Google Scholar] [CrossRef]
- Jin, G.; Weinberg, A. Human antimicrobial peptides and cancer. Semin. Cell Dev. Biol. 2019, 88, 156–162. [Google Scholar] [CrossRef]
- Mylonakis, E.; Podsiadlowski, L.; Muhammed, M.; Vilcinskas, A. Diversity, evolution and medical applications of insect antimicrobial peptides. Philos. Trans. R. Soc. B. 2016, 371, 20150290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, J.; Liu, Y.; Suo, W.; Zhao, R.; Fan, H. Effects of the antimicrobial peptide of Tenebrio molitor Linnaeus on cell cycle of K562 and inhibitory effects of that on cell proliferation compared with hydroxyurea. Chin. J. Vector Biol. Control. 2010, 21, 324–326. [Google Scholar]
- Ratcliffe, N.A.; Mello, C.B.; Garcia, E.S.; Butt, T.M.; Azambuja, P. Insect natural products and processes: New treatments for human disease. Insect Biochem. Mol. Biol. 2011, 41, 747–769. [Google Scholar] [CrossRef]
- Feng, Y.; Zhao, M.; Ding, W.; Chen, X. Overview of edible insect resources and common species utilisation in China. J. Insects Food. Feed 2020, 6, 13–25. [Google Scholar] [CrossRef]
- Schiappa, J.; Van Hee, R. From ants to staples: History and ideas concerning suturing techniques. Acta Chir. Belg. 2012, 112, 395–402. [Google Scholar] [CrossRef]
- Hemingway, J.; Shretta, R.; Wells, T.N.; Bell, D.; Djimdé, A.A.; Achee, N.; Qi, G. Tools and strategies for malaria control and elimination: What do we need to achieve a grand convergence in malaria? PLoS Biol. 2016, 14, e1002380. [Google Scholar] [CrossRef] [PubMed]
- Tonk, M.; Pierrot, C.; Cabezas-Cruz, A.; Rahnamaeian, M.; Khalife, J.; Vilcinskas, A. The Drosophila melanogaster antimicrobial peptides Mtk-1 and Mtk-2 are active against the malarial parasite Plasmodium falciparum. Parasitol. Res. 2019, 118, 1993–1998. [Google Scholar] [CrossRef]
- Liu, C.; Wang, C.; Yao, H. Comprehensive Resource Utilization of Waste Using the Black Soldier Fly (Hermetia illucens (L.)) (Diptera: Stratiomyidae). Animals 2019, 9, 349. [Google Scholar] [CrossRef] [Green Version]
- Ushakova, N.; Dontsov, A.; Sakina, N.; Bastrakov, A.; Ostrovsky, M. Antioxidative properties of melanins and ommochromes from black soldier fly Hermetia illucens. Biomolecules 2019, 9, 408. [Google Scholar] [CrossRef] [Green Version]
- Diener, S.; Zurbruegg, C.; Tockner, K. Conversion of organic material by black soldier fly larvae: Establishing optimal feeding rates. Waste Manag. Res. 2009, 27, 603–610. [Google Scholar] [CrossRef] [PubMed]
- Gobbi, P.; Martinez-Sanchez, A.; Rojo, S. The effects of larval diet on adult life-history traits of the black soldier fly, Hermetia illucens (Diptera: Stratiomyidae). Eur. J. Entomol. 2013, 110, 461–468. [Google Scholar] [CrossRef]
- Yi, H.; Chowdhury, M.; Huang, Y.; Yu, X. Insect antimicrobial peptides and their applications. Appl. Microbiol. Biotechnol. 2014, 98, 5807–5822. [Google Scholar] [CrossRef] [Green Version]
- Boman, H.G.; Nilsson-Faye, I.; Paul, K.; Rasmuson, T. Insect immunity I. Characteristics of an inducible cell-free antibacterial reaction in hemolymph of Samia cynthia pupae. Infect. Immun. 1974, 10, 136–145. [Google Scholar] [CrossRef] [Green Version]
- Hultmark, D.; STEINER, H.; RASMUSON, T.; BOMAN, H.G. Insect immunity. Purification and properties of three inducible bactericidal proteins from hemolymph of immunized pupae of Hyalophora cecropia. Eur. J. Biochem. 1980, 106, 7–16. [Google Scholar] [CrossRef]
- Wang, Y.; Lai, R. Insect antimicrobial peptides: Structures, properties and gene regulation. Zool. Res. 2010, 31, 27–34. [Google Scholar] [CrossRef]
- Imler, J.L.; Hoffmann, J.A. Signaling mechanisms in the antimicrobial host defense of Drosophila. Curr. Opin. Mocrobiol. 2000, 3, 16–22. [Google Scholar] [CrossRef]
- Bulet, P.; Stöcklin, R.; Menin, L. Anti-microbial peptides: From invertebrates to vertebrates. Immunol. Rev. 2010, 198, 169–184. [Google Scholar] [CrossRef] [PubMed]
- Zyowska, M.; Wyszynska, A.; Jagusztyn-Krynicka, E.K. Antimicrobial peptides - Defensins. Postep. Mikrobiol. 2011, 50, 223–234. [Google Scholar]
- Park, S.-I.; Yoe, S.M. A novel cecropin-like peptide from black soldier fly, Hermetia illucens: Isolation, structural and functional characterization. Entomol. Res. 2017, 47, 115–124. [Google Scholar] [CrossRef]
- Park, S.-I.; Yoe, S.M. Defensin-like peptide3 from black solder fly: Identification, characterization, and key amino acids for anti-Gram-negative bacteria. Entomol. Res. 2017, 47, 41–47. [Google Scholar] [CrossRef]
- Lee, K.-S.; Yun, E.-Y.; Goo, T.-W. Antimicrobial Activity of an Extract of Hermetia illucens Larvae Immunized with Lactobacillus casei against Salmonella Species. Insects 2020, 11, 704. [Google Scholar] [CrossRef]
- Shin, H.S.; Park, S.-I. Novel attacin from Hermetia illucens: cDNA cloning, characterization, and antibacterial properties. Prep. Biochem. Biotechnol. 2019, 49, 279–285. [Google Scholar] [CrossRef]
- Park, S.-I.; Kim, J.-W.; Yoe, S.M. Purification and characterization of a novel antibacterial peptide from black soldier fly (Hermetia illucens) larvae. Dev. Comp. Immunol. 2015, 52, 98–106. [Google Scholar] [CrossRef]
- Čeřovský, V.; Bém, R. Lucifensins, the insect defensins of biomedical importance: The story behind maggot therapy. Pharmaceuticals 2014, 7, 251–264. [Google Scholar] [CrossRef] [Green Version]
- Cornet, B.; Bonmatin, J.-M.; Hetru, C.; Hoffmann, J.A.; Ptak, M.; Vovelle, F. Refined three-dimensional solution structure of insect defensin A. Structure 1995, 3, 435–448. [Google Scholar] [CrossRef] [Green Version]
- Epand, R.M.; Vogel, H.J. Diversity of antimicrobial peptides and their mechanisms of action. Biochim. Biophys. Acta 1999, 1462, 11–28. [Google Scholar] [CrossRef] [Green Version]
- Jozefiak, A.; Engberg, R.M. Insect proteins as a potential source of antimicrobial peptides in livestock production. A review. J. Anim. Feed. Sci. 2017, 26, 87–99. [Google Scholar] [CrossRef]
- Sato, H.; Feix, J.B. Peptide–membrane interactions and mechanisms of membrane destruction by amphipathic α-helical antimicrobial peptides. Biochim. Biophys. Acta 2006, 1758, 1245–1256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bessin, Y.; Saint, N.; Marri, L.; Marchini, D.; Molle, G. Antibacterial activity and pore-forming properties of ceratotoxins: A mechanism of action based on the barrel stave model. Biochim. Biophys. Acta 2004, 1667, 148–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, T.; Umegawa, Y.; Yamagami, M.; Suzuki, T.; Tsuchikawa, H.; Hanashima, S.; Matsumori, N.; Murata, M. The perpendicular orientation of amphotericin B methyl ester in hydrated lipid bilayers supports the barrel-stave model. Biochemistry 2019, 58, 2282–2291. [Google Scholar] [CrossRef] [PubMed]
- Duclohier, H. How do channel-and pore-forming helical peptides interact with lipid membranes and how does this account for their antimicrobial activity? Mini-Rev. Med. Chem. 2002, 2, 331–342. [Google Scholar] [CrossRef]
- Nicolas, P. Multifunctional host defense peptides: Intracellular-targeting antimicrobial peptides. FEBS 2009, 276, 6483–6496. [Google Scholar] [CrossRef]
- Cytryńska, M.; Rahnamaeian, M.; Zdybicka-Barabas, A.; Dobslaff, K.; Züchner, T.; Sacheau, G.; Innis, C.A.; Vilcinskas, A. Proline-rich antimicrobial peptides in medicinal maggots of Lucilia sericata interact with bacterial DnaK but do not inhibit protein synthesis. Front. Pharmacol 2020, 11, 532. [Google Scholar] [CrossRef]
- He, J.; Furmanski, P. Sequence specificity and transcriptional activation in the binding of lactoferrin to DNA. Nature 1995, 373, 721–724. [Google Scholar] [CrossRef]
- Singh, C.; Vaishna, R.; Kakkar, A.; Arunkumar, K.; Nagaraju, J. Characterization of antiviral and antibacterial activity of B ombyx mori seroin proteins. Cell. Microbiol. 2014, 16, 1354–1365. [Google Scholar] [CrossRef]
- Kurata, S. Recognition and elimination of diversified pathogens in insect defense systems. Mol. Divers. 2006, 10, 599–605. [Google Scholar] [CrossRef]
- Kurata, S.; Ariki, S.; Kawabata, S.-i. Recognition of pathogens and activation of immune responses in Drosophila and horseshoe crab innate immunity. Immunobiology 2006, 211, 237–249. [Google Scholar] [CrossRef]
- Royet, J.; Reichhart, J.-M.; Hoffmann, J.A. Sensing and signaling during infection in Drosophila. Curr. Opin. Immunol. 2005, 17, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Horng, T.; Medzhitov, R. Drosophila MyD88 is an adapter in the Toll signaling pathway. Proc. Natl. Acad. Sci. USA 2001, 98, 12654–12658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tauszig-Delamasure, S.; Bilak, H.; Capovilla, M.; Hoffmann, J.A.; Imler, J.-L. Drosophila MyD88 is required for the response to fungal and Gram-positive bacterial infections. Nat. Immunol. 2002, 3, 91–97. [Google Scholar] [CrossRef]
- Sun, H.; Bristow, B.N.; Qu, G.; Wasserman, S.A. A heterotrimeric death domain complex in Toll signaling. Proc. Natl. Acad. Sci. USA 2002, 99, 12871–12876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, T.; Towb, P.; Wasserman, S.A.; Sprang, S.R. Three-dimensional structure of a complex between the death domains of Pelle and Tube. Cell 1999, 99, 545–555. [Google Scholar] [CrossRef] [Green Version]
- Moncrieffe, M.C.; Grossmann, J.G.; Gay, N.J. Assembly of oligomeric death domain complexes during Toll receptor signaling. J. Biol. Chem. 2008, 283, 33447–33454. [Google Scholar] [CrossRef] [Green Version]
- Moynagh, P.N. The Pellino family: IRAK E3 ligases with emerging roles in innate immune signalling. Trends Immunol. 2008, 30, 33–42. [Google Scholar] [CrossRef]
- Valanne, S.; Wang, J.-H.; Rämet, M. The Drosophila toll signaling pathway. J. Immunol 2011, 186, 649–656. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Xu, W.; Gao, X.; Li, W.; Qi, S.; Guo, D.; Ajayi, O.E.; Ding, S.-W.; Wu, Q. lncRNA sensing of a viral suppressor of RNAi activates non-canonical innate immune signaling in Drosophila. Cell Host Microbe 2020, 27, 115–128. [Google Scholar] [CrossRef]
- Kaneko, T.; Goldman, W.E.; Mellroth, P.; Steiner, H.; Fukase, K.; Kusumoto, S.; Harley, W.; Fox, A.; Golenbock, D.; Silverman, N. Monomeric and polymeric gram-negative peptidoglycan but not purified LPS stimulate the Drosophila IMD pathway. Immunity 2004, 20, 637–649. [Google Scholar] [CrossRef] [Green Version]
- Sarvari, M.; Mikani, A.; Mehrabadi, M. The innate immune gene Relish and Caudal jointly contribute to the gut immune homeostasis by regulating antimicrobial peptides in Galleria mellonella. Dev. Comp. Immunol. 2020, 110, 103732. [Google Scholar] [CrossRef] [PubMed]
- Atefyekta, S.; Pihl, M.; Lindsay, C.; Heilshorn, S.C.; Andersson, M. Antibiofilm elastin-like polypeptide coatings:functionality, stability, and selectivity. Acta Biomater. 2019, 83, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Yu, Y.; Zhan, S.; Tomberlin, J.K.; Huang, D.; Cai, M.; Zheng, L.; Yu, Z.; Zhang, J. Dual oxidase Duox and Toll-like receptor 3 TLR3 in the Toll pathway suppress zoonotic pathogens through regulating the intestinal bacterial community homeostasis in Hermetia illucens L. PLoS ONE 2020, 15, e0225873. [Google Scholar] [CrossRef]
- Fariz Zahir Ali, M.; Ohta, T.; Ido, A.; Miura, C.; Miura, T. The dipterose of black soldier fly (Hermetia illucens) induces innate immune response through toll-like receptor pathway in mouse macrophage RAW264. 7 cells. Biomolecules 2019, 9, 677. [Google Scholar] [CrossRef] [Green Version]
- Andersson, D.I.; Hughes, D.; Kubicek-Sutherland, J.Z. Mechanisms and consequences of bacterial resistance to antimicrobial peptides. Drug. Resist. Update 2016, 26, 43–57. [Google Scholar] [CrossRef]
- Joo, H.-S.; Fu, C.-I.; Otto, M. Bacterial strategies of resistance to antimicrobial peptides. Philos. Trans. R. Soc. B. 2016, 371, 20150292. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Xiang, Q.; Zhang, Q.; Huang, Y.; Su, Z. Overview on the recent study of antimicrobial peptides: Origins, functions, relative mechanisms and application. Peptides 2012, 37, 207–215. [Google Scholar] [CrossRef]
- Olsen, I. Biofilm-specific antibiotic tolerance and resistance. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 877–886. [Google Scholar] [CrossRef]
- Rodríguez-Rojas, A.; Makarova, O.; Rolff, J. Antimicrobials, stress and mutagenesis. PLoS Pathog 2014, 10, e1004445. [Google Scholar] [CrossRef] [Green Version]
- Nadeem, S.F.; Gohar, U.F.; Tahir, S.F.; Mukhtar, H.; Pornpukdeewattana, S.; Nukthamna, P.; Moula Ali, A.M.; Bavisetty, S.C.B.; Massa, S. Antimicrobial resistance: More than 70 years of war between humans and bacteria. Crit. Rev. Microbiol. 2020, 46, 578–599. [Google Scholar] [CrossRef] [PubMed]
- Peschel, A.; Vuong, C.; Otto, M.; Götz, F. The d-Alanine Residues ofStaphylococcus aureus Teichoic Acids Alter the Susceptibility to Vancomycin and the Activity of Autolytic Enzymes. Antimicrob. Agents Chemother. 2000, 44, 2845–2847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenfeld, Y.; Shai, Y. Lipopolysaccharide (Endotoxin)-host defense antibacterial peptides interactions: Role in bacterial resistance and prevention of sepsis. Biochim. Biophys. Acta 2006, 1758, 1513–1522. [Google Scholar] [CrossRef] [Green Version]
- McCoy, A.J.; Liu, H.; Falla, T.J.; Gunn, J.S. Identification of Proteus mirabilisMutants with increased sensitivity to antimicrobial peptides. Antimicrob. Agents Chemother. 2001, 45, 2030–2037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, G.D. Bacterial resistance to antibiotics: Enzymatic degradation and modification. Adv. Drug Del. Rev. 2005, 57, 1451–1470. [Google Scholar] [CrossRef]
- Jin, T.; Bokarewa, M.; Foster, T.; Mitchell, J.; Higgins, J.; Tarkowski, A. Staphylococcus aureus Resists Human Defensins by Production of Staphylokinase, a Novel Bacterial Evasion Mechanism. J. Immunol. 2004, 172, 1169–1176. [Google Scholar] [CrossRef] [Green Version]
- Guina, T.; Yi, E.C.; Wang, H.; Hackett, M.; Miller, S.I. A PhoP-Regulated Outer Membrane Protease of Salmonella enterica Serovar Typhimurium Promotes Resistance to Alpha-Helical Antimicrobial Peptides. J. Bacteriol. 2000, 182, 4077–4086. [Google Scholar] [CrossRef] [Green Version]
- Starr, C.G.; Wimley, W.C. Antimicrobial peptides are degraded by the cytosolic proteases of human erythrocytes. Biochim. Biophys. Acta 2017, 1859, 2319–2326. [Google Scholar] [CrossRef]
- Chernysh, S.; Gordya, N.; Suborova, T. Insect Antimicrobial Peptide Complexes Prevent Resistance Development in Bacteria. PLoS ONE 2015, 10, e0130788. [Google Scholar] [CrossRef] [Green Version]
- Thomas, S.; Karnik, S.; Barai, R.S.; Jayaraman, V.K.; Idicula-Thomas, S. CAMP: A useful resource for research on antimicrobial peptides. Nucleic Acids Res. 2010, 38, 774–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moretta, A.; Salvia, R.; Scieuzo, C.; Di Somma, A.; Vogel, H.; Pucci, P.; Sgambato, A.; Wolff, M.; Falabella, P. A bioinformatic study of antimicrobial peptides identified in the Black Soldier Fly (BSF) Hermetia illucens (Diptera: Stratiomyidae). Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef]
- Barragan-Fonseca, K.B.; Dicke, M.; van Loon, J.J. Nutritional value of the black soldier fly (Hermetia illucens L.) and its suitability as animal feed–a review. J. Insects. Food. Feed 2017, 3, 105–120. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Tomberlin, J.K.; Vanlaerhoven, S. Ability of black soldier fly (Diptera: Stratiomyidae) larvae to recycle food waste. Environ. Entomol. 2015, 44, 406–410. [Google Scholar] [CrossRef]
- Newton, G.; Sheppard, D.; Watson, D.; Burtle, G.; Dove, C.; Tomberlin, J.; Thelen, E. The black soldier fly, Hermetia illucens, as a manure management/resource recovery tool. In Proceedings of the Symposium on the state of the science of Animal Manure and Waste Management, San Antonio, TX, USA, 5–7 January 2005; pp. 5–7. [Google Scholar]
- St-Hilaire, S.; Sheppard, C.; Tomberlin, J.K.; Irving, S.; Newton, L.; McGuire, M.A.; Mosley, E.E.; Hardy, R.W.; Sealey, W. Fly prepupae as a feedstuff for rainbow trout, Oncorhynchus mykiss. J. World Aquacult. Soc. 2007, 38, 59–67. [Google Scholar] [CrossRef]
- Newton, L.; Sheppard, C.; Watson, D.W.; Burtle, G.; Dove, R. Using the Black Soldier Fly, Hermetia illucens, as a Value-Added Tool for the Management of Swine Manure; Animal and Poultry Waste Management Center, North Carolina State University: Raleigh, NC, USA, 2005; p. 17. [Google Scholar]
- Oonincx, D.; Van Huis, A.; Van Loon, J. Nutrient utilisation by black soldier flies fed with chicken, pig, or cow manure. J. Insects. Food. Feed 2015, 1, 131–139. [Google Scholar] [CrossRef]
- Rachmawati, R.; Buchori, D.; Hidayat, P.; Hem, S.; Fahmi, M.R. Perkembangan dan kandungan nutrisi larva Hermetia illucens (Linnaeus)(Diptera: Stratiomyidae) pada bungkil Kelapa Sawit. J. Entomol. Res. 2015, 7, 28–41. [Google Scholar] [CrossRef] [Green Version]
- Newton, G.; Booram, C.; Barker, R.; Hale, O. Dried Hermetia illucens larvae meal as a supplement for swine. J. Anim. Sci. 1977, 44, 395–400. [Google Scholar] [CrossRef]
- Newton, G.; Sheppard, D.; Burtle, G. Research Briefs: Black Soldier Fly Prepupae—A Compelling Alternative to Fish Meal and Fish Oil. Aquaculture 2008, 24, 103–109. [Google Scholar]
- Kroeckel, S.; Harjes, A.-G.; Roth, I.; Katz, H.; Wuertz, S.; Susenbeth, A.; Schulz, C. When a turbot catches a fly: Evaluation of a pre-pupae meal of the Black Soldier Fly (Hermetia illucens) as fish meal substitute—Growth performance and chitin degradation in juvenile turbot (Psetta maxima). Aquaculture 2012, 364, 345–352. [Google Scholar] [CrossRef]
- Mesonero-Escuredo, S.; Strutzberg-Minder, K.; Casanovas, C.; Segalés, J. Viral and bacterial investigations on the aetiology of recurrent pig neonatal diarrhoea cases in Spain. Porcine Health Manag. 2018, 4, 1–6. [Google Scholar] [CrossRef]
- Zaatout, N.; Ayachi, A.; Kecha, M.; Kadlec, K. Identification of staphylococci causing mastitis in dairy cattle from Algeria and characterization of Staphylococcus aureus. J. Appl. Microbiol. 2019, 127, 1305–1314. [Google Scholar] [CrossRef]
- Al-Quraishy, S.; Qasem, M.A.; Al-Shaebi, E.M.; Murshed, M.; Mares, M.M.; Dkhil, M.A. Rumex nervosus changed the oxidative status of chicken caecum infected with Eimeria tenella. J. King Saud Univ Sci. 2020, 32, 2207–2211. [Google Scholar] [CrossRef]
- Driemeyer, H. Evaluation of Black Soldier Fly (Hermetia illucens) Larvae as an Alternative Protein Source in Pig Creep Diets in Relation to Production, Blood and Manure Microbiology Parameters; Stellenbosch University: Stellenbosch, South Africa, 2016. [Google Scholar]
- Biasato, I.; Renna, M.; Gai, F.; Dabbou, S.; Meneguz, M.; Perona, G.; Martinez, S.; Lajusticia, A.C.B.; Bergagna, S.; Sardi, L. Partially defatted black soldier fly larva meal inclusion in piglet diets: Effects on the growth performance, nutrient digestibility, blood profile, gut morphology and histological features. J. Anim. Sci. Biotechno. 2019, 10, 12. [Google Scholar] [CrossRef]
- Spranghers, T.; Michiels, J.; Vrancx, J.; Ovyn, A.; Eeckhout, M.; De Clercq, P.; De Smet, S. Gut antimicrobial effects and nutritional value of black soldier fly (Hermetia illucens L.) prepupae for weaned piglets. Anim. Feed Sci. Technol. 2017, 235, 33–42. [Google Scholar] [CrossRef]
- Liu, C.; Yao, H.; Chapman, S.J.; Su, J.; Wang, C. Changes in gut bacterial communities and the incidence of antibiotic resistance genes during degradation of antibiotics by black soldier fly larvae. Environ. Int. 2020, 142, 105834. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Yao, H.; Wang, C. Black Soldier Fly Larvae Can Effectively Degrade Oxytetracycline Bacterial Residue by Means of the Gut Bacterial Community. Front. Microbiol. 2021, 12, e663972. [Google Scholar] [CrossRef]
- Dyar, O.J.; Zhang, T.; Peng, Y.; Sun, M.; Sun, C.; Yin, J.; Ding, L.; Sun, C.; Wang, Y.; Sun, Q. Knowledge, attitudes and practices relating to antibiotic use and antibiotic resistance among backyard pig farmers in rural Shandong province, China. Prev. Vet. Med. 2020, 175, 104858. [Google Scholar] [CrossRef]
- Khan, A.; Aziz, H.; Khan, N.; Hasan, M.; Ahmed, S.; Farooqi, I.; Dhingra, A.; Vambol, V.; Changani, F.; Yousefi, M. Impact, disease outbreak and the eco-hazards associated with pharmaceutical residues: A Critical review. Int. J. Environ. Sci. Technol. 2021, 1–12. [Google Scholar] [CrossRef]
- Huang, F.; An, Z.; Moran, M.J.; Liu, F. Recognition of typical antibiotic residues in environmental media related to groundwater in China (2009−2019). J. Hazard. Mater. 2020, 399, 122813. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.E. Peptide antibiotics. Lancet 1997, 349, 418–422. [Google Scholar] [CrossRef]
- Andrä, J.; Berninghausen, O.; Leippe, M. Cecropins, antibacterial peptides from insects and mammals, are potently fungicidal against Candida albicans. Med. Microbiol. Immunol. 2001, 189, 169–173. [Google Scholar] [CrossRef]
- Chaturvedi, P.; Bhat, R.A.H.; Pande, A. Antimicrobial peptides of fish: Innocuous alternatives to antibiotics. Rev. Aquacult. 2020, 12, 85–106. [Google Scholar] [CrossRef]
- Leni, G.; Tedeschi, T.; Faccini, A.; Pratesi, F.; Folli, C.; Puxeddu, I.; Migliorini, P.; Gianotten, N.; Jacobs, J.; Depraetere, S. Shotgun proteomics, in-silico evaluation and immunoblotting assays for allergenicity assessment of lesser mealworm, black soldier fly and their protein hydrolysates. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Wu, N.; Wang, X.; Xu, X.; Cai, R.; Xie, S. Effects of heavy metals on the bioaccumulation, excretion and gut microbiome of black soldier fly larvae (Hermetia illucens). Ecotoxicol. Environ. Saf. 2020, 192, 110323. [Google Scholar] [CrossRef] [PubMed]
- Lalander, C.; Senecal, J.; Calvo, M.G.; Ahrens, L.; Josefsson, S.; Wiberg, K.; Vinnerås, B. Fate of pharmaceuticals and pesticides in fly larvae composting. Sci. Total Environ. 2016, 565, 279–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Q.; Deng, W.; Gao, Z.; Li, M.; Liu, W.; Wang, X.; Zhu, F. Effect of sulfonamide pollution on the growth of manure management candidate Hermetia illucens. PLoS ONE 2019, 14, e0216086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
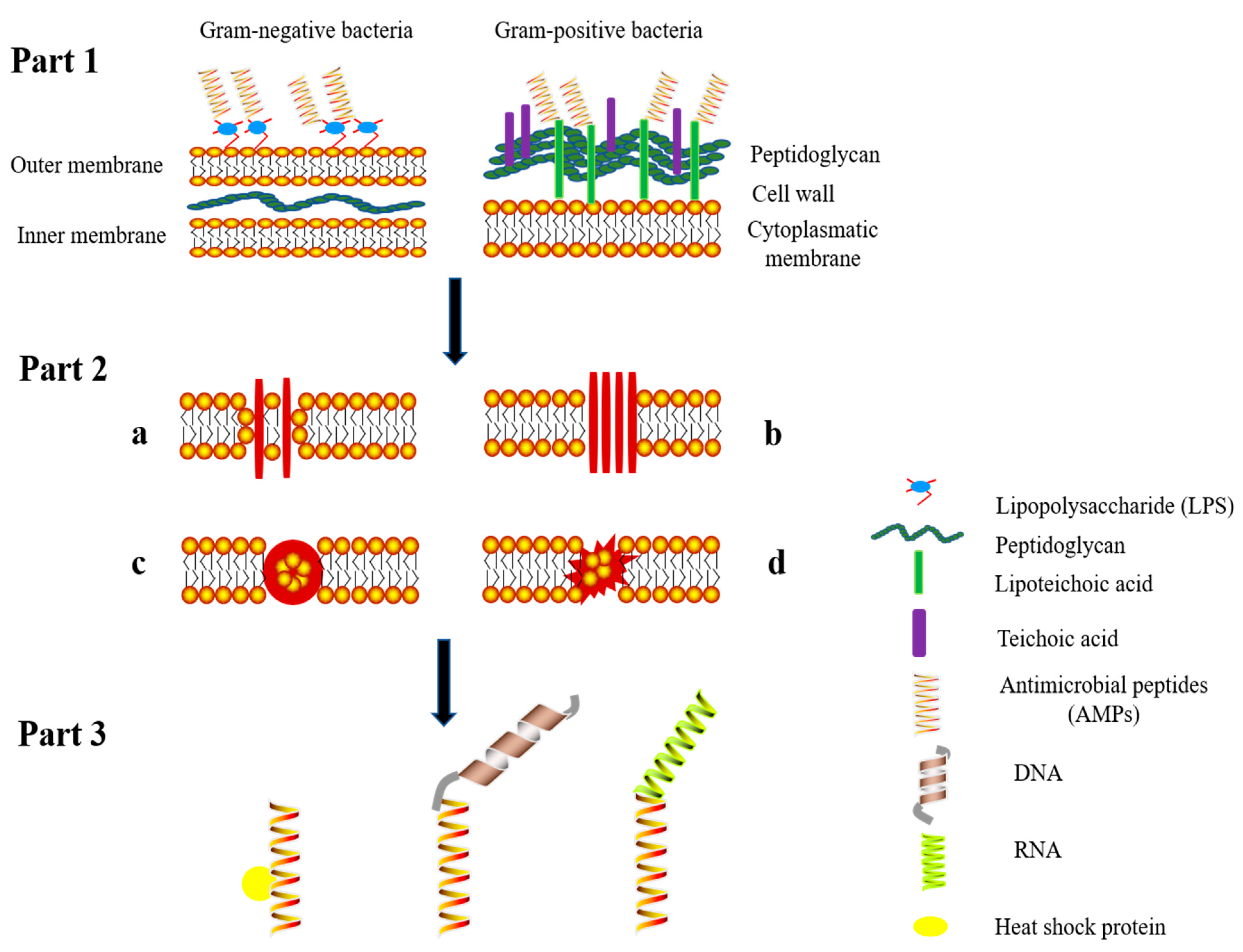

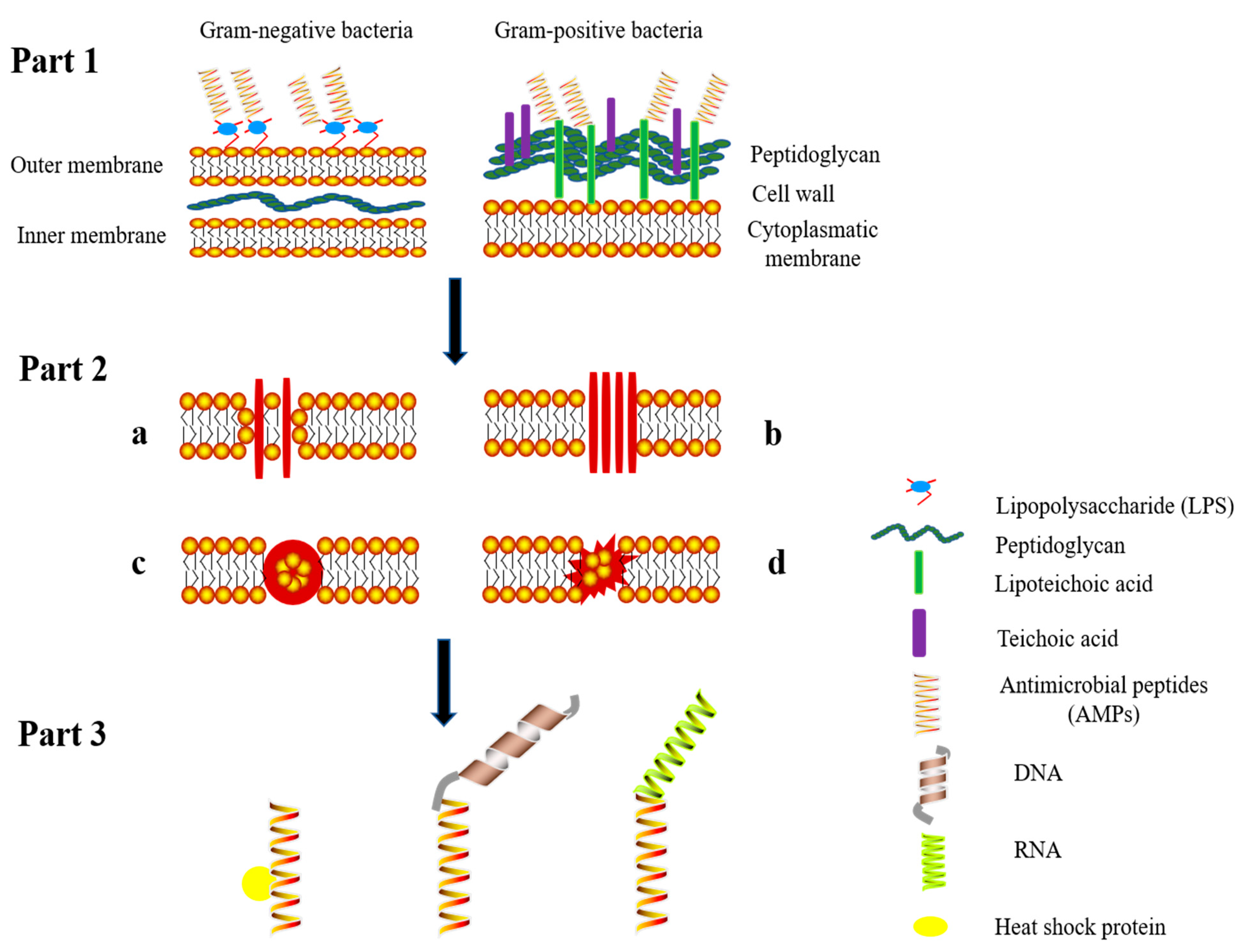{kind=link}
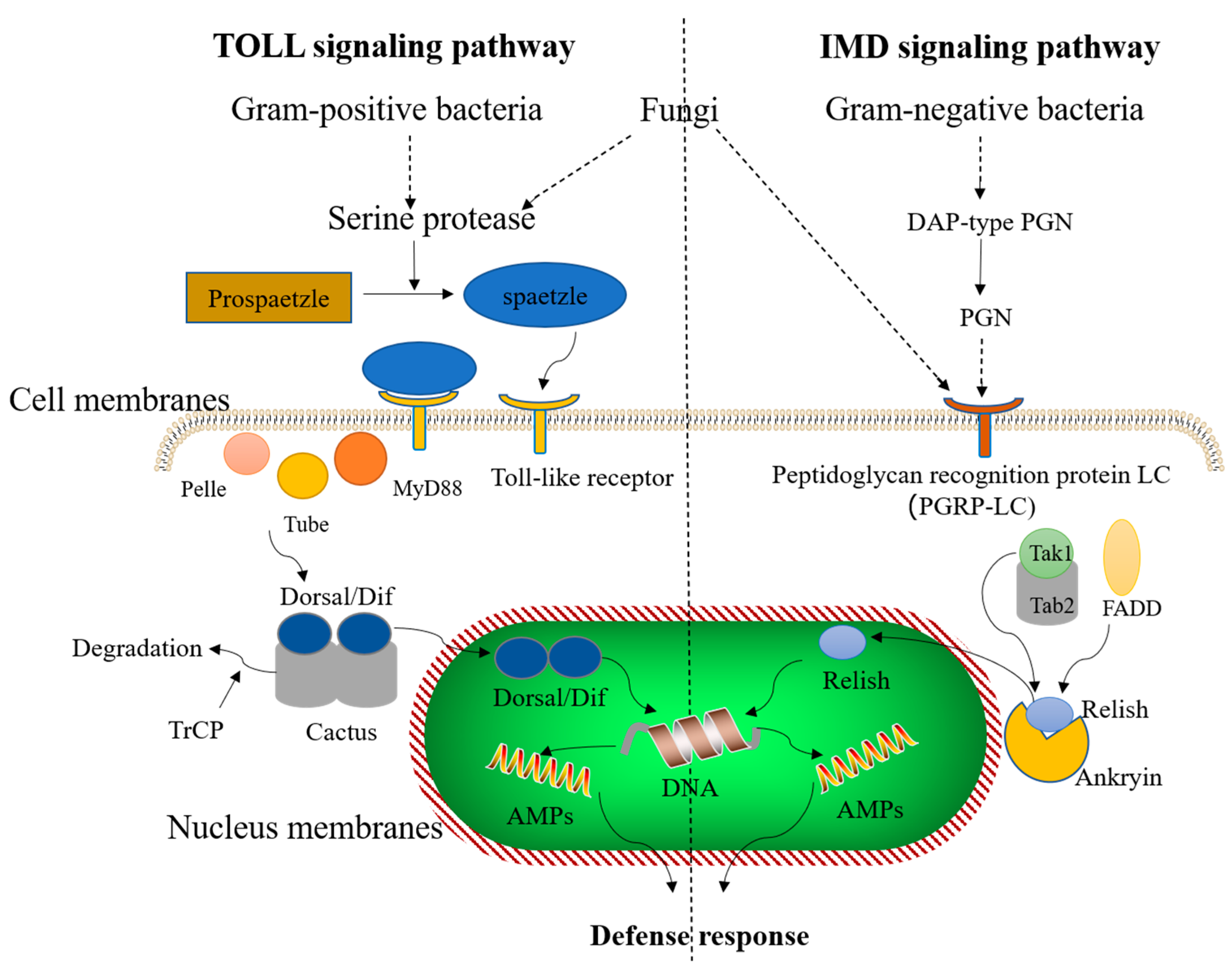{kind=link}
| Peptide | Amino Acid Sequence | Immune-Induced Strains | Reference |
|---|---|---|---|
| Defensin | |||
| Defensin-like peptide 1 (DLP1) | MRSVLVLGLIVAAFAVYTSAQPYQLQYEEDGLDQAVELPIEEEQLPSQVVEQHYRAKRATCDLLSPFKVGHAACALHCIALGRRGGWCDGRAVCNCRR | Staphylococcus aureus KCCM 40881 | [59] |
| Defensin-like peptide 2 (DLP2) | MRSILVLGLIVAAFAVYTSAQPYQLQYEEDGPGYALELPSEEEGLPSQVVEQHYRAKRATCDLLSPFKVGHAACALHCIAMGRRGGWCDGRAVCNCRR | Staphylococcus aureus KCCM 40881 | [59] |
| Defensin-like peptide 3 (DLP3) | MRSILVLGLIVAVFGVYTSAQPYQLQYEEDGPEYALVLPIEEEELPSQVVEQHYRAKRATCDLLSPFGVGHAACAVHCIAMGRRGGWCDDRAVCNCRR | Staphylococcus aureus KCCM 40881 | [56,59] |
| Defensin-like peptide 4 (DLP4) | MVHCQPFQLETEGDQQLEPVVAEVDDVVDLVAIPEHTREKRATCDLLSPFKVGHAACAAHCIARGKRGGWCDKRAVCNCRK | Staphylococcus aureus KCCM 40881 | [59] |
| Defensin 1 (HiDef1) | unknown | Lactobacillus casei | [57] |
| Cecropin | |||
| CecropinZ1 | GWLKKIGKMKFILGTTLAIVIAIFGQCQAATWSYNPNGGATVTWTANVAATAR | Escherichia. coli and Staphylococcus aureus | [29] |
| Cecropin 1 (Hicec1) | unknown | Lactobacillus casei | [57] |
| Cecropin-like peptide 1 (CLP1) | MNFTKLFVVFAVVLVAFAGQSEAGWRKRVFKPVEKFGQRVRDAGVQGIAIAQQGANVLATARGGPPQQG | Staphylococcus aureus KCCM 40881 | [55] |
| Cecropin-like peptide 2 (CLP2) | MNFAKLFVVFAIVLVAFSGQSEAGWWKRVFKPVEKLGQRVRDAGIQGLEIAQQGANVLATARGGPPQQG | Staphylococcus aureus KCCM 40881 | [55] |
| Cecropin-like peptide 3 (CLP3) | MNFTKLFVVFAVVLIAFSGQSEAGWWKRVFKPVERLGQRVRDAGIQGLEIAQQGANVLATVRGGPPQQG | Staphylococcus aureus KCCM 40881 | [55] |
| Attacin | |||
| Hermetia illucens-attacin | MASKFLGNPNHNIGGGVFAAGNTRSNTPSLGAFGTLNLKDHSLGVSHTITPGVSDTFSQNARLNILKTPDHRVDANVFNSHTRLNNGFAFDKRGGSLDYTHRAGHGLSLGASHIPKFGTTAELTGKANLWRSPSGLSTFDLTGSASRTFGGPMAGRNNFGAGLGFSHRF | Escherichia. coli KCCM 11234 | [58] |
| Sarcotoxin | |||
| Sarcotoxin1 | GWLKRKIGMKFILGTTLAIVVAIFGQCQAATWSYNPNGGATVTWTANVAATAR | Escherichia. coli and Staphylococcus aureus | [29] |
| Sarcotoxin (2a) | GWLKRKIGKKFILGTTLAIVVAIFGQCQAATWSYNPNGGATVTWTANVAATAR | Escherichia. coli and Staphylococcus aureus | [29] |
| Sarcotoxin (2b) | GWLKRKIGKKFILGTTLAIAVAIFGQCQAATWSYNPNGGATVTWTANVAATAR | Escherichia. coli and Staphylococcus aureus | [29] |
| Sarcotoxin3 | GWLKRKIGMMMKNSNFNSTEEREAAKKNYKRKYVPWFSGANVAATAR | Escherichia. coli and Staphylococcus aureus | [29] |
| Stomoxyn | |||
| StomoxynZH1 (a) | RGFRKHFNNLPICVEGLAGDIGSILLGVGSDIGALAGAIANLALIAGECAAQGEAGAAVVAAT | Escherichia. coli and Staphylococcus aureus | [29] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xia, J.; Ge, C.; Yao, H. Antimicrobial Peptides from Black Soldier Fly (Hermetia illucens) as Potential Antimicrobial Factors Representing an Alternative to Antibiotics in Livestock Farming. Animals 2021, 11, 1937. https://doi.org/10.3390/ani11071937
Xia J, Ge C, Yao H. Antimicrobial Peptides from Black Soldier Fly (Hermetia illucens) as Potential Antimicrobial Factors Representing an Alternative to Antibiotics in Livestock Farming. Animals. 2021; 11(7):1937. https://doi.org/10.3390/ani11071937
Chicago/Turabian StyleXia, Jing, Chaorong Ge, and Huaiying Yao. 2021. "Antimicrobial Peptides from Black Soldier Fly (Hermetia illucens) as Potential Antimicrobial Factors Representing an Alternative to Antibiotics in Livestock Farming" Animals 11, no. 7: 1937. https://doi.org/10.3390/ani11071937
APA StyleXia, J., Ge, C., & Yao, H. (2021). Antimicrobial Peptides from Black Soldier Fly (Hermetia illucens) as Potential Antimicrobial Factors Representing an Alternative to Antibiotics in Livestock Farming. Animals, 11(7), 1937. https://doi.org/10.3390/ani11071937