
Comparison of Ovarian Morphology and Follicular Disturbances between Two Inbred Strains of Cotton Rats (Sigmodon hispidus)

 ,
,  , , , , and
, , , , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Ovary Processing
2.2. Histological Analysis and Histoplanimetry
2.3. Immunohistochemistry
2.4. TUNEL Assay
2.5. Electron Microscopy
2.6. Statistical Analysis
3. Results
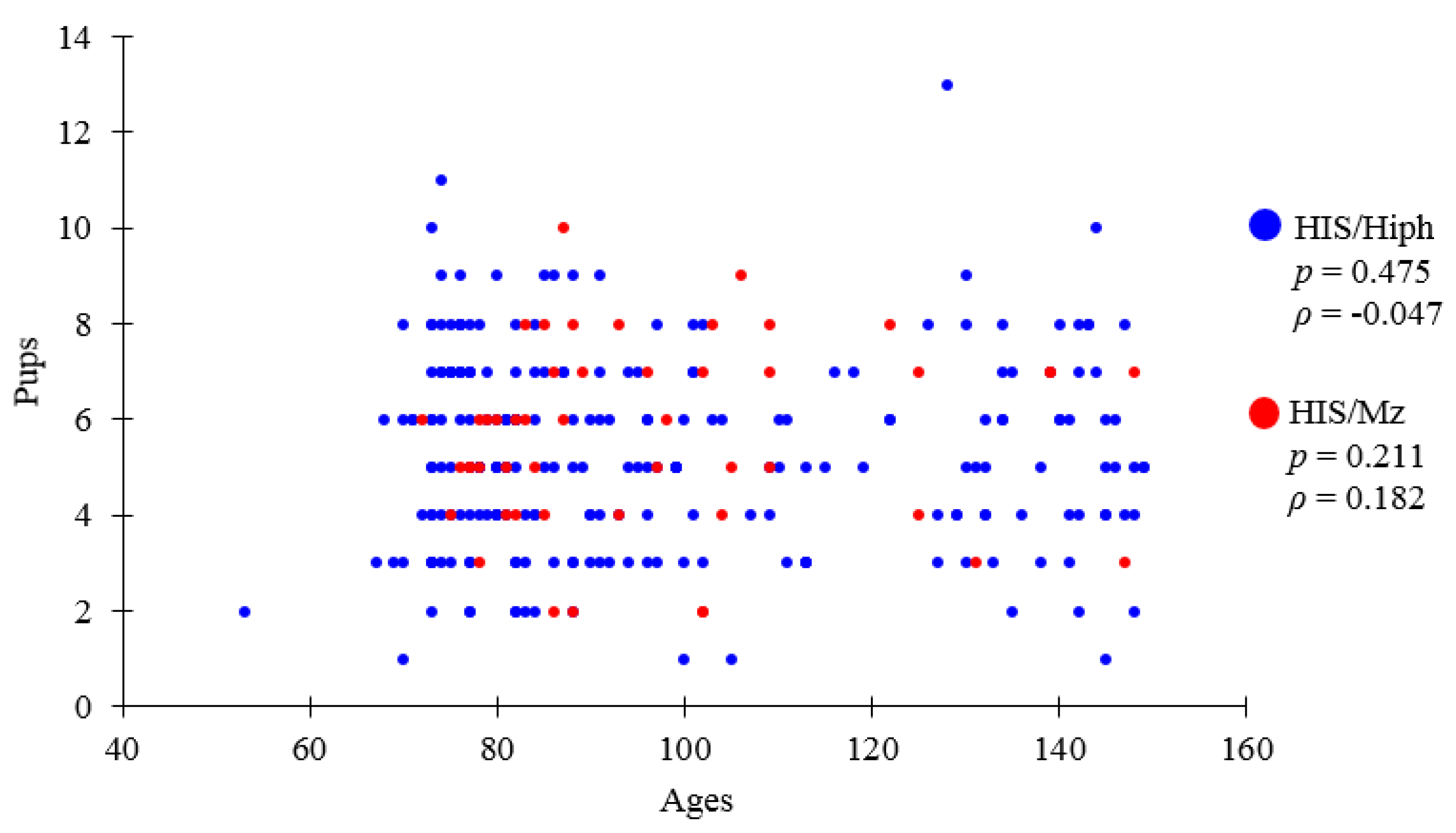
3.1. Comparative Results of Recorded Data of HIS/Hiph and HIS/Mz
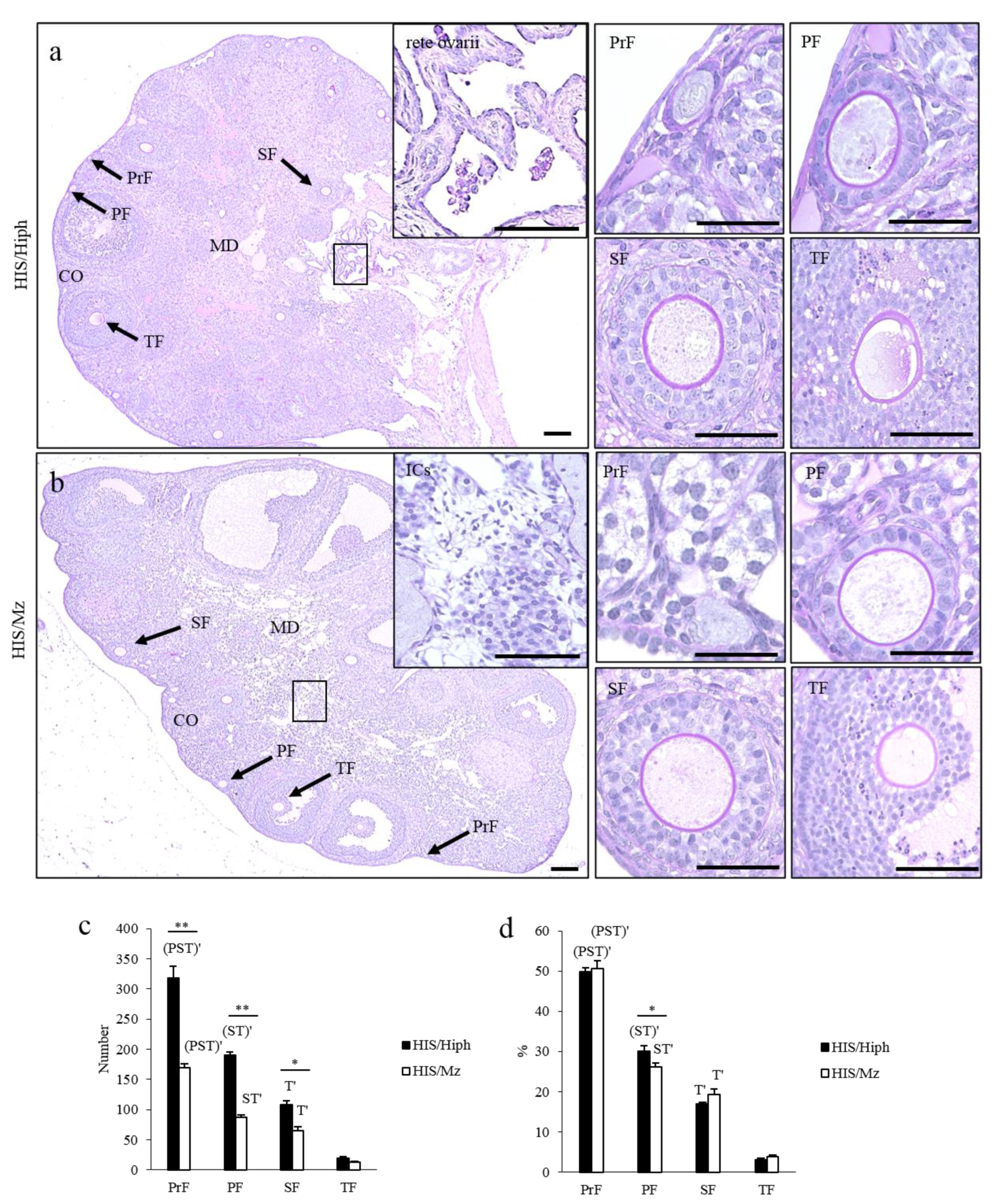
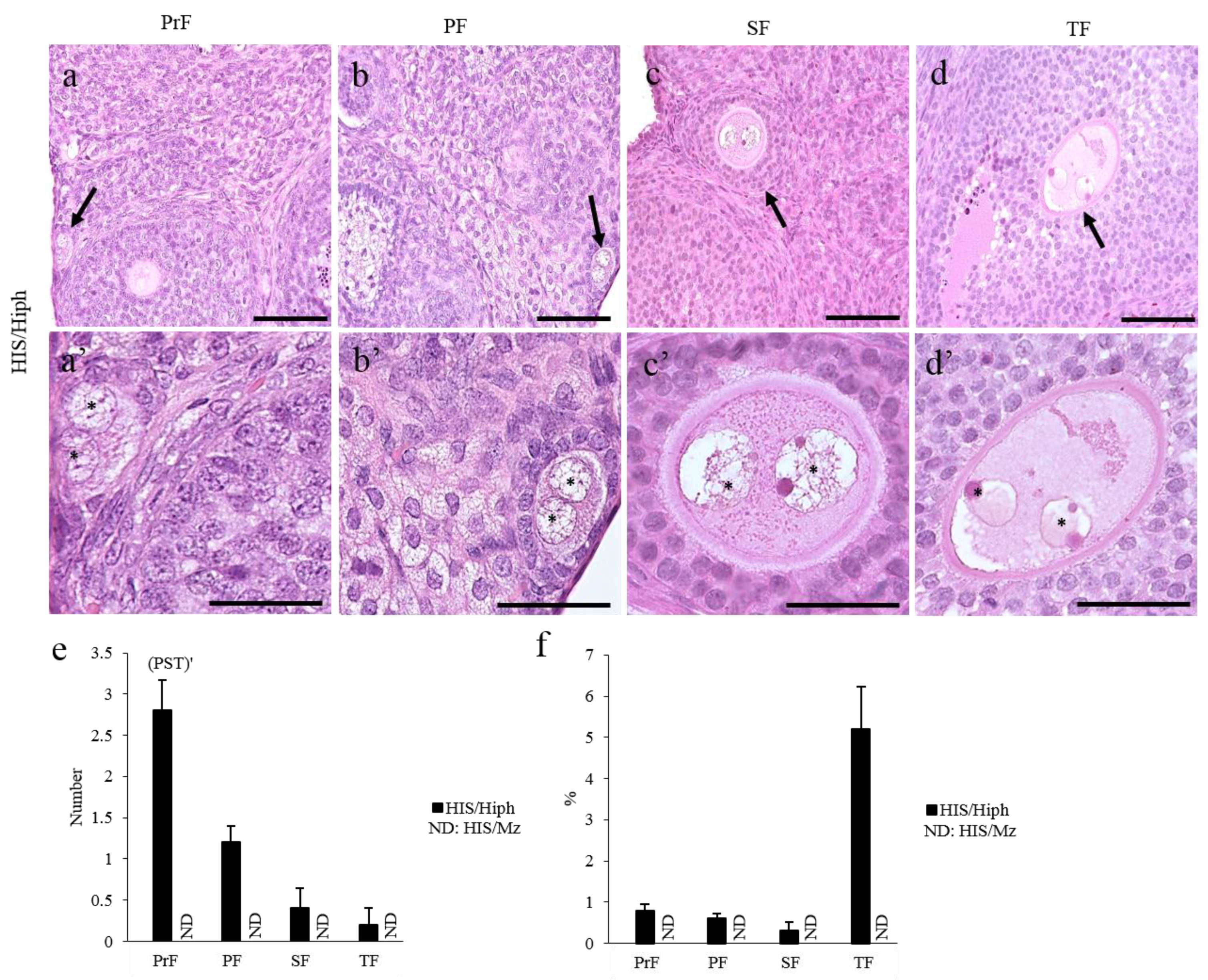
3.2. Comparative Ovarian Histology of HIS/Hiph and HIS/Mz
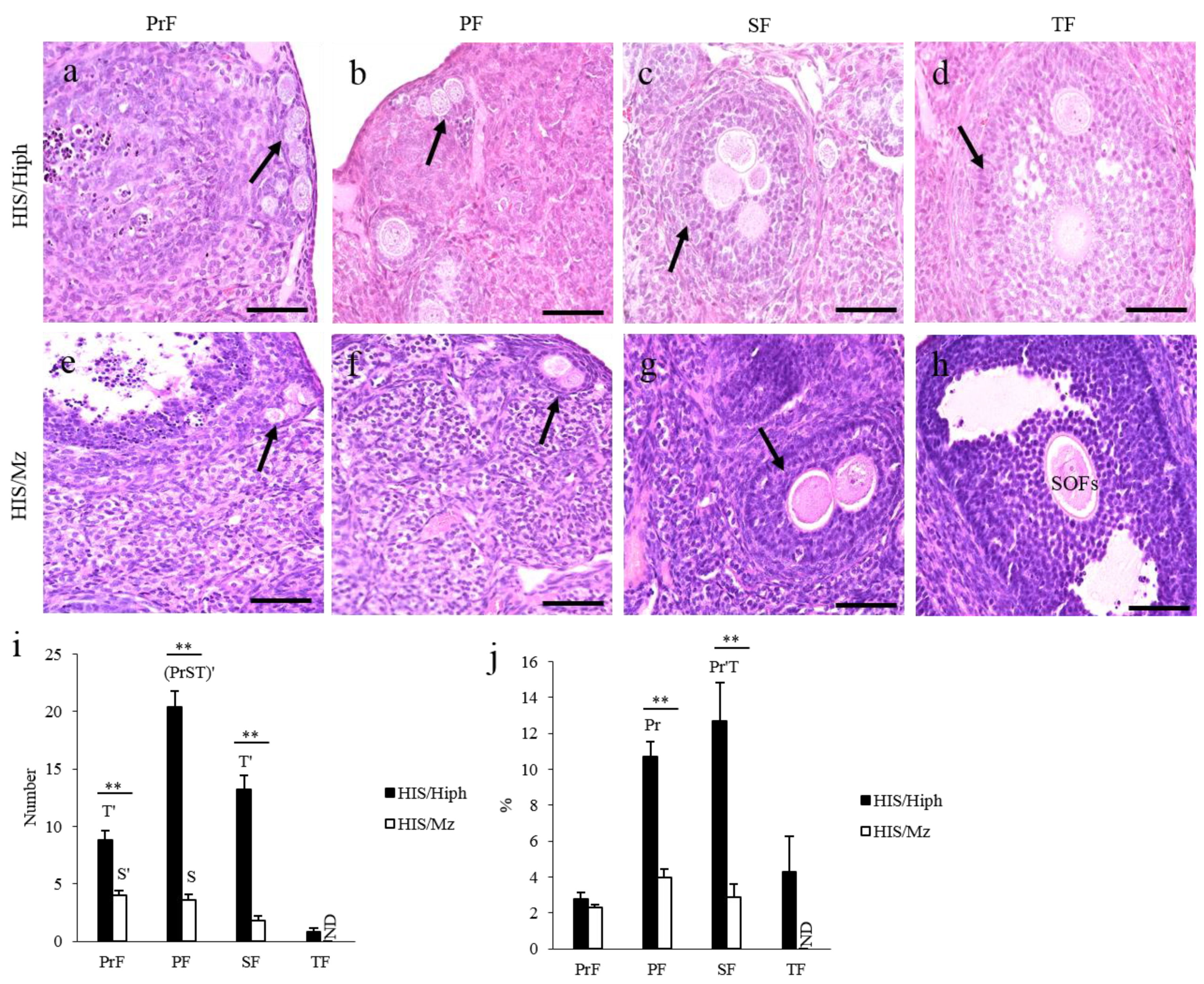
3.3. MOFs Found in the Ovaries of HIS/Hiph and HIS/Mz
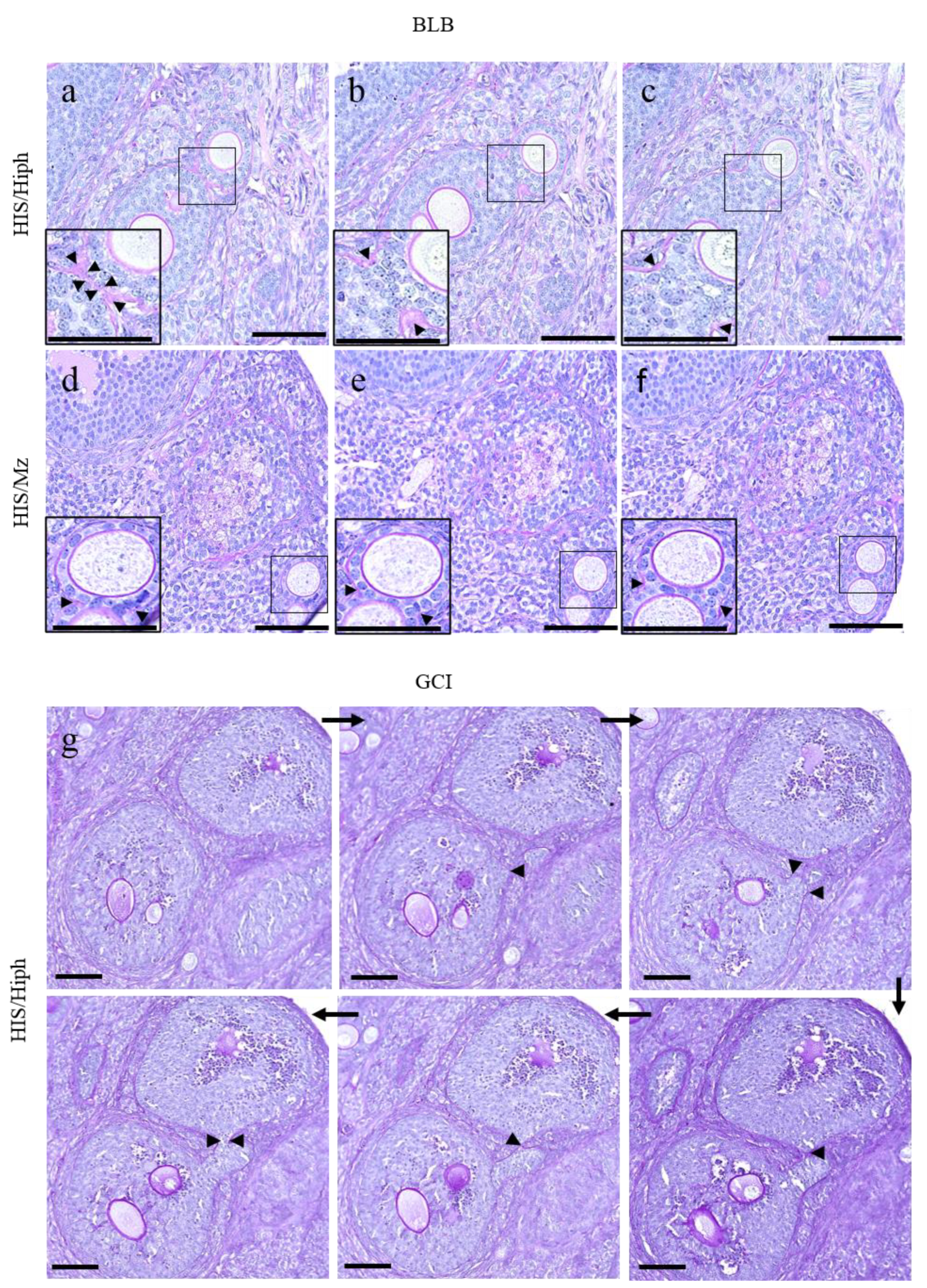
3.4. Unique Morphologies Found in the MOF of HIS/Hiph and HIS/Mz
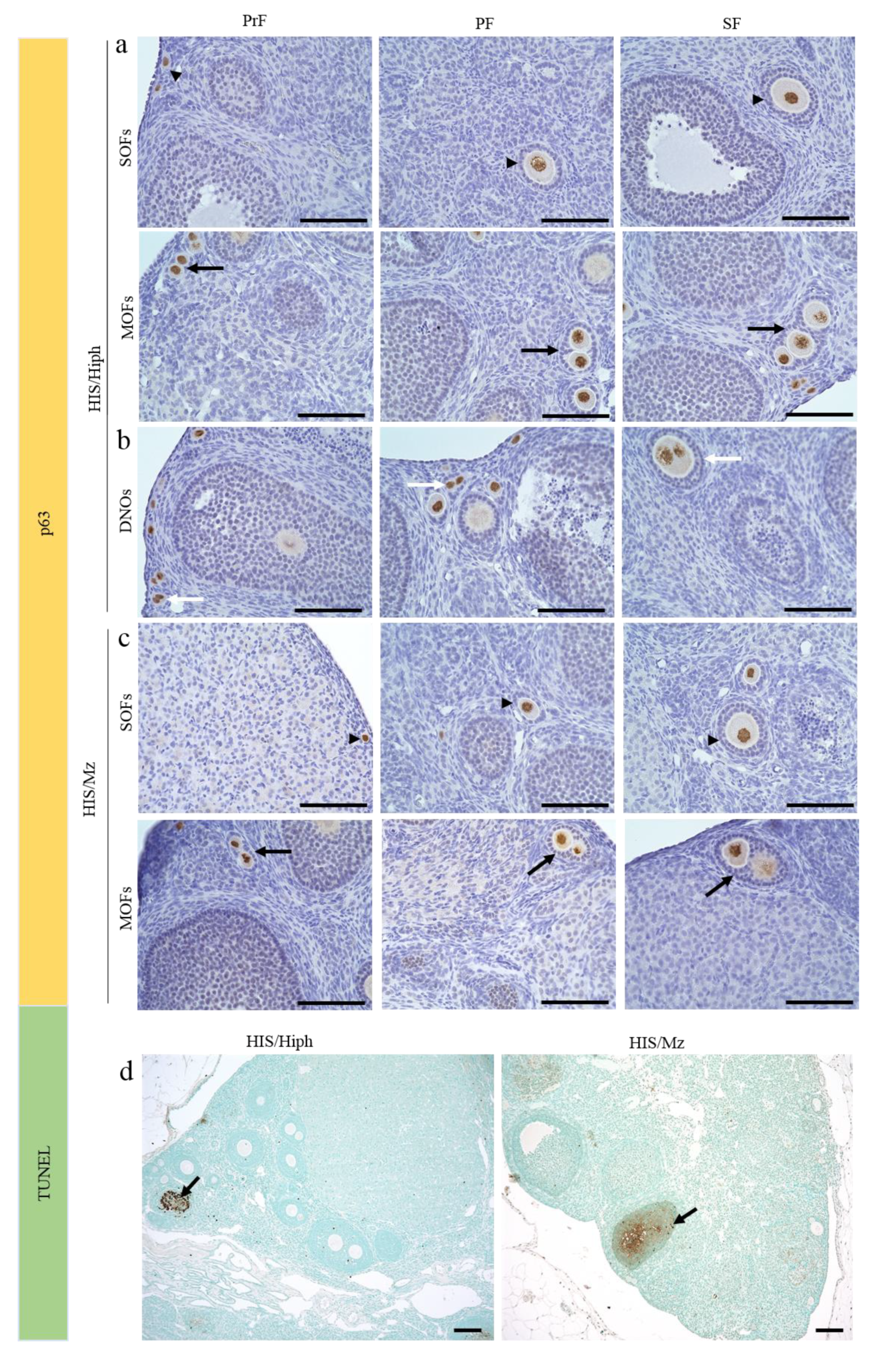
3.5. DNOs Found in the Ovaries of HIS/Hiph and HIS/Mz
3.6. Follicular Cell-Cycle Arrest and Oocyte Death in the Ovaries of HIS/Hiph and HIS/Mz
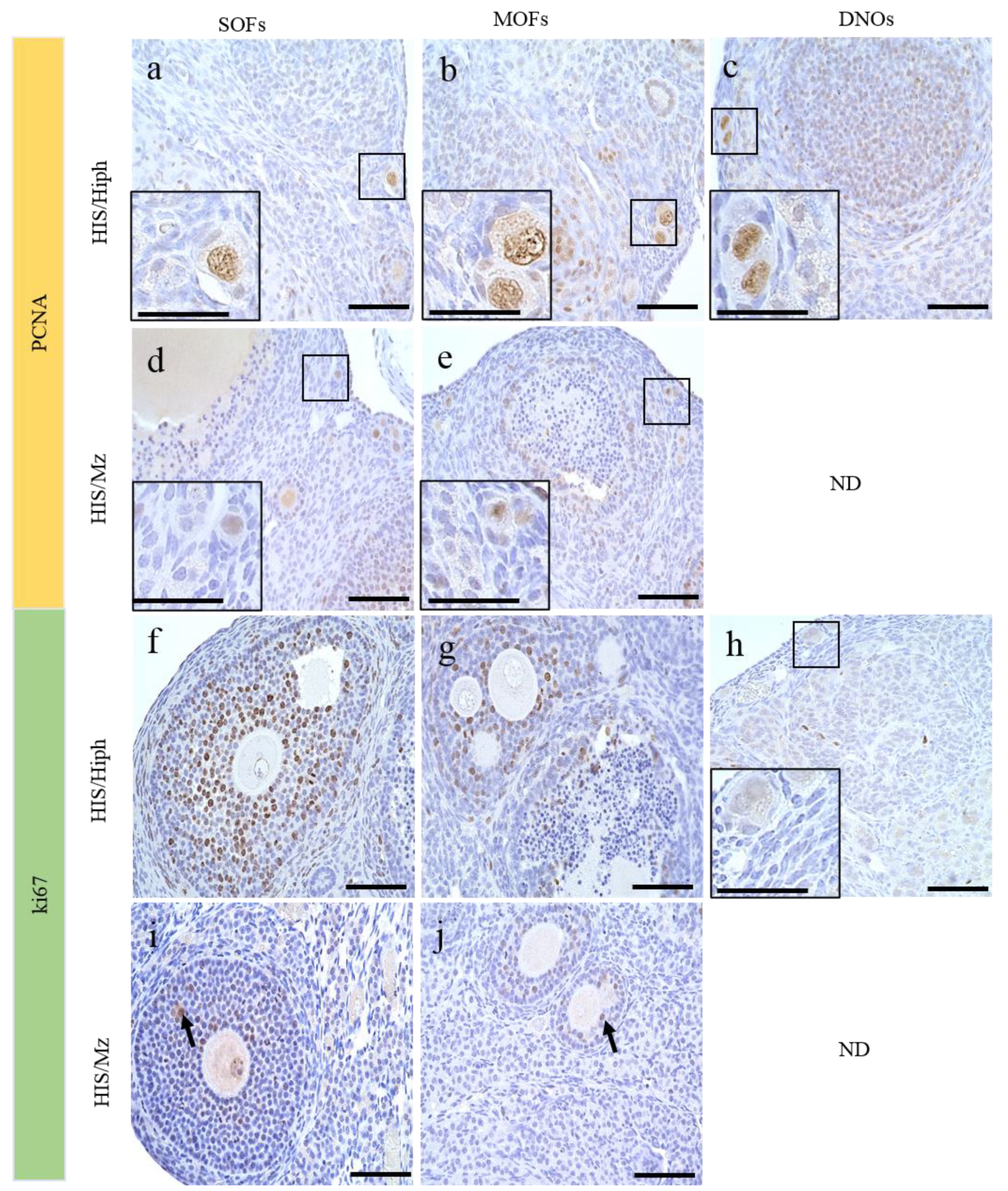
3.7. Proliferative Activity of the Oocytes in the Ovaries of HIS/Hiph and HIS/Mz
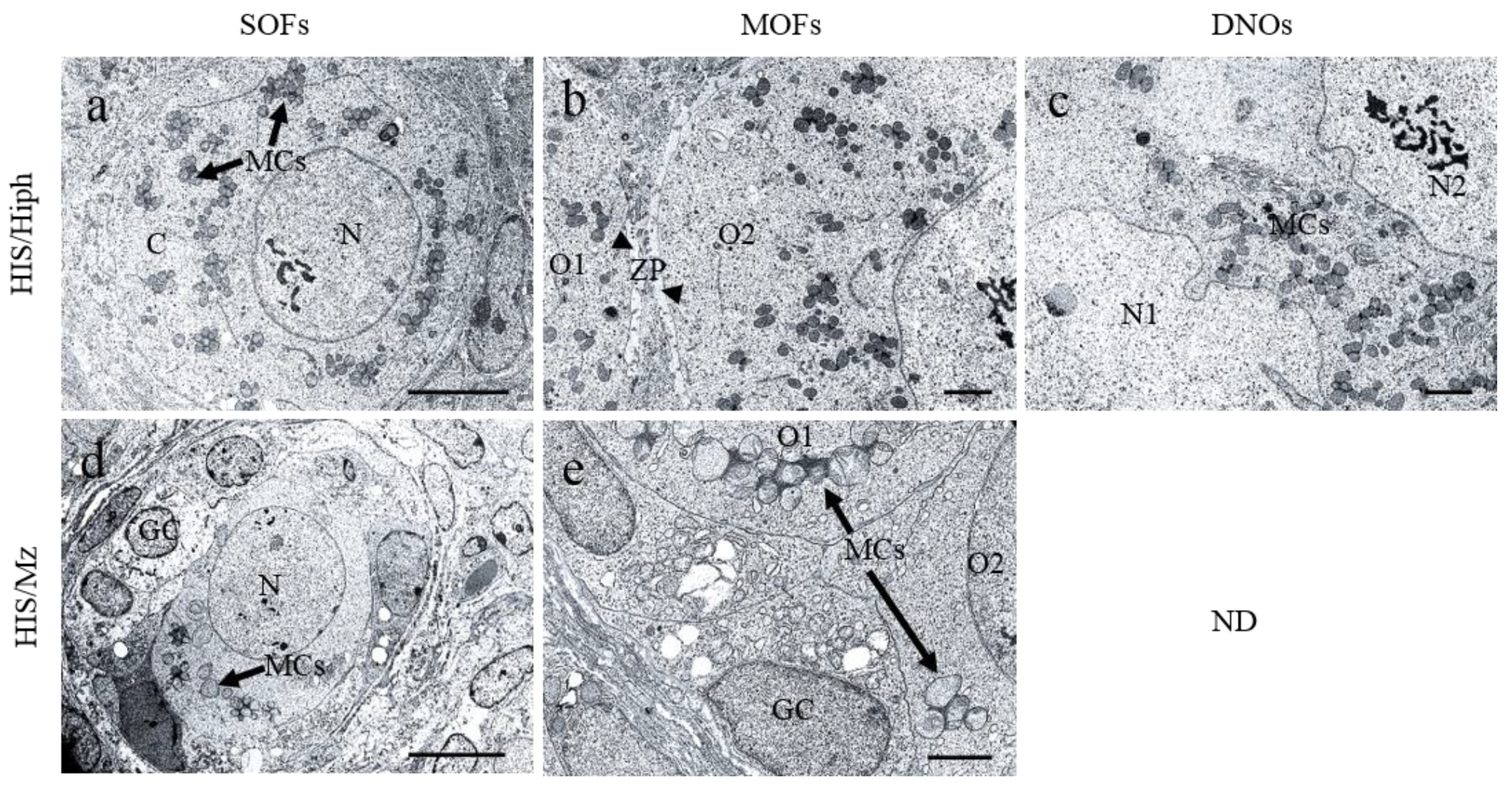
3.8. Ultrastructures of SOFs, MOFs, and DNOs in the Ovaries of HIS/Hiph and HIS/Mz
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Monniaux, D.; Clément, F.; Dalbiès-Tran, R.; Estienne, A.; Fabre, S.; Mansanet, C.; Monget, P. The ovarian reserve of primordial follicles and the dynamic reserve of antral growing follicles: What is the link. Biol. Reprod. 2014, 90, 85. [Google Scholar] [CrossRef]
- Tingen, C.; Kim, A.; Woodruff, T.K. The primordial pool of follicles and nest breakdown in mammalian ovaries. Mol. Hum. Reprod. 2009, 15, 795–803. [Google Scholar] [CrossRef]
- Islam, M.R.; Ichii, O.; Nakamura, T.; Irie, T.; Masum, M.A.; Hosotani, M.; Otani, Y.; Elewa, Y.H.A.; Kon, Y. Unique morphological characteristics in the ovary of cotton rat (Sigmodon hispidus). J. Reprod. Dev. 2020, 66, 529–538. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.R.; Ichii, O.; Nakamura, T.; Irie, T.; Masum, M.A.; Otani, Y.; Namba, T.; Chuluunbaatar, T.; Elewa, Y.H.A.; Kon, Y. Developmental Changes of the Ovary in Neonatal Cotton Rat (Sigmodon hispidus). Front. Physiol. 2021, 11, 601927. [Google Scholar] [CrossRef] [PubMed]
- Ireland, J.L.; Scheetz, D.; Jimenez-Krassel, F.; Themmen, A.P.; Ward, F.; Lonergan, P.; Smith, G.W.; Perez, G.I.; Evans, A.C.; Ireland, J.J. Antral follicle count reliably predicts number of morphologically healthy oocytes and follicles in ovaries of young adult cattle. Biol. Reprod. 2008, 79, 1219–1225. [Google Scholar] [CrossRef] [PubMed]
- Silva-Santos, K.C.; Santos, G.M.; Siloto, L.S.; Hertel, M.F.; Andrade, E.R.; Rubin, M.I.; Sturion, L.; Melo-Sterza, F.A.; Seneda, M.M. Estimate of the population of preantral follicles in the ovaries of Bos taurus indicus and Bos taurus taurus cattle. Theriogenology 2011, 76, 1051–1057. [Google Scholar] [CrossRef]
- Payan-Carreira, R.; Pires, M.A. Multioocyte follicles in domestic dogs: A survey of frequency of occurrence. Theriogenology 2008, 69, 977–982. [Google Scholar] [CrossRef] [PubMed]
- Reynaud, K.; de Lesegno, C.V.; Chebrout, M.; Thoumire, S.; Chastant-Maillard, S. Follicle population, cumulus mucification, and oocyte chromatin configuration during the periovulatory period in the female dog. Theriogenology 2009, 72, 1120–1131. [Google Scholar] [CrossRef] [PubMed]
- Greenwald, G.S.; Moor, R.M. Isolation and preliminary characterization of pig primordial follicles. J. Reprod. Fertil. 1989, 87, 561–571. [Google Scholar] [CrossRef]
- Stankiewicz, T.; Błaszczyk, B.; Udała, J. A study on the occurrence of polyovular follicles in porcine ovaries with particular reference to intrafollicular hormone concentrations, quality of oocytes and their in vitro fertilization. Anat. Histol. Embryol. 2009, 38, 233–239. [Google Scholar] [CrossRef]
- Hadek, R. Morphological and histochemical study on the ovary of the sheep. Am. J. Vet. Res. 1958, 19, 873–881. [Google Scholar]
- Oliveira, R.L.; Silva, C.B.; Silva, E.O.; Gerez, J.R.; Santos, M.M.; Sarapião, F.D.; Sartori, L.; Bracarense, A.P.F.R.L.; Seneda, M.M. Proliferative activity of multi-oocyte follicles in sheep ovaries. Small Rum. Res. 2017, 146, 58–60. [Google Scholar] [CrossRef]
- Telfer, E.; Gosden, R.G. A quantitative cytological study of polyovular follicles in mammalian ovaries with particular reference to the domestic bitch (Canis familiaris). J. Reprod. Fertil. 1987, 81, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Bristol-Gould, S.; Woodruff, T.K. Folliculogenesis in the domestic cat (Felis catus). Theriogenology 2006, 66, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Gougeon, A. Frequent occurrence of multiovular and multinuclear oocytes in the adult human ovary. Fertil. Steril. 1981, 35, 417–422. [Google Scholar] [CrossRef]
- Prince, G.A. The cotton rat in biomedical research. AWIC News 1994, 5, 3–5, ISSN: 1050-561X. [Google Scholar]
- Faith, R.E.; Montgomery, C.A.; Durfee, W.J.; Aguilar-Cordova, E.; Wyde, P.R. The cotton rat in biomedical research. Lab. Anim. Sci. 1997, 47, 337–345. [Google Scholar]
- Nakamura, T.; Ichii, O.; Irie, T.; Mizoguchi, T.; Shinohara, A.; Kouguchi, H.; Sunden, Y.; Otsuka-Kanazawa, S.; Elewa, Y.H.A.; Koshimoto, C.; et al. Cotton rats (Sigmodon hispidus) possess pharyngeal pouch remnants originating from different primordia. Histol. Histopathol. 2018, 33, 555–565. [Google Scholar] [CrossRef]
- Nakamura, T.; Ichii, O.; Irie, T.; Kouguchi, H.; Sotozaki, K.; Chihara, M.; Sunden, Y.; Nagasaki, K.I.; Tatsumi, O.; Elewa, Y.H.A.; et al. Cotton rat (Sigmodon hispidus) develops metabolic disorders associated with visceral adipose inflammation and fatty pancreas without obesity. Cell Tissue Res. 2019, 375, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Ichii, O.; Nakamura, T.; Irie, T.; Kouguchi, H.; Nakamura, D.; Nakamura, S.; Sato, S.; Yokoyama, K.; Horino, T.; Sunden, Y.; et al. Female cotton rats (Sigmodon hispidus) develop chronic anemia with renal inflammation and cystic changes. Histochem. Cell Biol. 2016, 146, 351–362. [Google Scholar] [CrossRef]
- Waldum, H.L.; Rørvik, H.; Falkmer, S.; Kawase, S. Neuroendocrine (ECL cell) differentiation of spontaneous gastric carcinomas of cotton rats (Sigmodon hispidus). Lab. Anim. Sci. 1999, 49, 241–247. [Google Scholar]
- Jefferson, W.N.; Couse, J.F.; Padilla-Banks, E.; Korach, K.S.; Newbold, R.R. Neonatal exposure to genistein induces estrogen receptor (ER)alpha expression and multioocyte follicles in the maturing mouse ovary: Evidence for ERbeta-mediated and nonestrogenic actions. Biol. Reprod. 2002, 67, 1285–1296. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Kim, H.; Kakuta, H.; Iguchi, T. Effects of 2,3-Bis(4-hydroxyphenyl)-propionitrile on induction of polyovular follicles in the mouse ovary. In Vivo 2018, 32, 19–24. [Google Scholar] [CrossRef]
- Su, W.; Guan, X.; Zhang, D.; Sun, M.; Yang, L.; Yi, F.; Hao, F.; Feng, X.; Ma, T. Occurrence of multi-oocyte follicles in aquaporin 8-deficient mice. Reprod. Biol. Endocrinol. 2013, 11, 88. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Sanz, J.; Arluzea, J.; Matorras, R.; González-Santiago, N.; Bilbao, J.; Yeh, N.; Barlas, A.; Romin, Y.; Manova-Todorova, K.; Koff, A.; et al. Increased number of multi-oocyte follicles (MOFs) in juvenile p27Kip1 mutant mice: Potential role of granulosa cells. Hum. Reprod. 2013, 4, 1023–1030. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Al-Mufti, W.; Bomsel-Helmreich, O.; Christidès, J.P. Oocyte size and intrafollicular position in polyovular follicles in rabbits. J. Reprod. Fertil. 1988, 82, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Gaytán, F.; Morales, C.; Manfredi-Lozano, M.; Tena-Sempere, M. Generation of multi-oocyte follicles in the peripubertal rat ovary: Link to the invasive capacity of granulosa cells. Fertil. Steril. 2014, 101, 1467–1476. [Google Scholar] [CrossRef] [PubMed]
- Lena, A.M.; Rossi, V.; Osterburg, S.; Smirnov, A.; Osterburg, C.; Tuppi, M.; Cappello, A.; Amelio, I.; Dötsch, V.; De Felici, M.; et al. The p63 C-terminus is essential for murine oocyte integrity. Nat. Commun. 2021, 12, 383. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, E.; Arai, K. Research on cotton rat breeding. Exp. Anim. 1967, 16, 121–126. [Google Scholar] [CrossRef][Green Version]
- Kon, Y.; Konno, A.; Hashimoto, Y.; Endoh, D. Ovarian cysts in MRL/MpJ mice originate from rete ovarii. Anat. Histol. Embryol. 2007, 36, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Lucci, C.M.; Amorim, C.A.; Rodrigues, A.P.; Figueiredo, J.R.; Báo, S.N.; Silva, J.R.; Gonçalves, P.B. Study of preantral follicle population in situ and after mechanical isolation from caprine ovaries at different reproductive stages. Anim. Reprod. Sci. 1999, 56, 223–236. [Google Scholar] [CrossRef][Green Version]
- McDougall, K.; Hay, M.A.; Goodrowe, K.L.; Gartley, C.J.; King, W.A. Changes in the number of follicles and of oocytes in ovaries of prepubertal, peripubertal and mature bitches. J. Reprod. Fertil. Suppl. 1997, 51, 25–31. [Google Scholar] [PubMed]
- Nakamuta, N.; Kobayashi, S. Expression of p63 in the mouse primordial germ cells. J. Vet. Med. Sci. 2004, 66, 1365–1370. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Oliveira, R.L.; Silva-Santos, K.C.; González, S.M.; Bizarro-Silva, C.; Machado, F.Z.; Bracarense, A.P.; Seneda, M.M. Proliferative activity of oocytes in multi-oocyte follicles of bovine ovary. Semin. Ciênc. Agrár. 2017, 38, 3591–3597. [Google Scholar] [CrossRef]
- Xu, B.; Hua, J.; Zhang, Y.; Jiang, X.; Zhang, H.; Ma, T.; Zheng, W.; Sun, R.; Shen, W.; Sha, J.; et al. Proliferating cell nuclear antigen (PCNA) regulates primordial follicle assembly by promoting apoptosis of oocytes in fetal and neonatal mouse ovaries. PLoS ONE 2011, 6, e16046. [Google Scholar] [CrossRef] [PubMed]
- Tilly, J.L. Ovarian follicle counts--not as simple as 1, 2, 3. Reprod. Biol. Endocrinol. 2003, 1, 11. [Google Scholar] [CrossRef] [PubMed]
- Uslu, B.; Dioguardi, C.C.; Haynes, M.; Miao, D.Q.; Kurus, M.; Hoffman, G.; Johnson, J. Quantifying growing versus non-growing ovarian follicles in the mouse. J. Ovarian Res. 2017, 10, 3. [Google Scholar] [CrossRef] [PubMed]
- Aoyama, M.; Shiraishi, A.; Matsubara, S.; Horie, K.; Osugi, T.; Kawada, T.; Yasuda, K.; Satake, H. Identification of a New Theca/Interstitial Cell-Specific Gene and Its Biological Role in Growth of Mouse Ovarian Follicles at the Gonadotropin-Independent Stage. Front. Endocrinol. 2019, 10, 553. [Google Scholar] [CrossRef]
- Tian, Y.; Shen, W.; Lai, Z.; Shi, L.; Yang, S.; Ding, T.; Wang, S.; Luo, A. Isolation and identification of ovarian theca-interstitial cells and granulose cells of immature female mice. Cell Biol. Int. 2015, 39, 584–590. [Google Scholar] [CrossRef]
- Hartman, C.G. Polynuclear ova and polyovular follicles in the opossum and other mammals with special reference to the problem of fecundity. Am. J. Anat. 1926, 37, 1–51. [Google Scholar] [CrossRef]
- Bodemer, C.W.; Warnick, S. Polyovular Follicles in the Immature Hamster Ovary: I. Polyovular Follicles in the Normal Intact Animal. Fertil. Steril. 1961, 12, 159–169. [Google Scholar] [CrossRef]
- Zeilmaker, G.H.; Alberda, A.T.; Gent, I.V. Fertilization and cleavage of oocytes from a binovular human ovarian follicle: A possible cause of dizygotic twinning and chimerism. Fertil. Steril. 1983, 40, 841–843. [Google Scholar] [CrossRef]
- Williams, S.A.; Stanley, P. Mouse fertility is enhanced by oocyte-specific loss of core 1-derived O-glycans. FASEB J. 2008, 22, 2273–2284. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhang, Z.; Hou, R.; Li, G.; Yu, J.; Zhang, A.; Wang, J.; He, G.; Lan, J.; Fujihara, N. Ultrastructure of “twin oocytes” in the ovaries of the giant pandas (Ailuropoda melanoleuca). J. Mamm. Ova. Res. 2001, 18, 48–51. [Google Scholar] [CrossRef][Green Version]
- Yano, K.; Hashida, N.; Kubo, T.; Ohashi, I.; Koizumi, A.; Kageura, R.; Furutani, K.; Yano, C. Repeated collection of conjoined oocytes from a patient with polycystic ovary syndrome, resulting in one successful live birth from frozen thawed blastocyst transfer: A case report. J. Assist. Reprod. Genet. 2017, 34, 1547–1552. [Google Scholar] [CrossRef] [PubMed]
- Donne, R.; Saroul-Aïnama, M.; Cordier, P.; Celton-Morizur, S.; Desdouets, C. Polyploidy in liver development, homeostasis and disease. Nat. Rev. Gastroenterol. Hepatol. 2020, 7, 391–405. [Google Scholar] [CrossRef]
- Macgregor, H.C.; Kezer, J. Gene amplification in oocytes with 8 germinal vesicles from the tailed frog Ascaphus truei Stejneger. Chromosoma 1970, 29, 189–206. [Google Scholar] [CrossRef] [PubMed]
- Robinson, E.S.; Stephenson, E.M.; Stephenson, N.G. Nuclear constitution of primary oocytes of the frog Leiopelma hochstetteri (Ascaphidae). Copeia 1973, 1, 173–176. [Google Scholar] [CrossRef]
- Muskhelishvili, L.; Wingard, S.K.; Latendresse, J.R. Proliferating cell nuclear antigen--a marker for ovarian follicle counts. Toxicol. Pathol. 2005, 33, 365–368. [Google Scholar] [CrossRef]
- Jaglarz, M.K.; Nowak, Z.; Biliński, S.M. The Balbiani body and generation of early asymmetry in the oocyte of a tiger beetle. Differentiation 2003, 71, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Marlow, F.L.; Mullins, M.C. Bucky ball functions in Balbiani body assembly and animal-vegetal polarity in the oocyte and follicle cell layer in zebrafish. Dev. Biol. 2008, 321, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Pepling, M.E.; Wilhelm, J.E.; O’Hara, A.L.; Gephardt, G.W.; Spradling, A.C. Mouse oocytes within germ cell cysts and primordial follicles contain a Balbiani body. Proc. Natl. Acad. Sci. USA 2007, 104, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Albamonte, M.I.; Albamonte, M.S.; Stella, I.; Zuccardi, L.; Vitullo, A.D. The infant and pubertal human ovary: Balbiani’s body-associated VASA expression, immunohistochemical detection of apoptosis-related BCL2 and BAX proteins, and DNA fragmentation. Hum. Reprod. 2013, 28, 698–706. [Google Scholar] [CrossRef] [PubMed]
- Sekula, M.; Tworzydlo, W.; Bilinski, S.M. Morphogenesis of the Balbiani body in developing oocytes of an orthopteran, Metrioptera brachyptera, and multiplication of female germline mitochondria. J. Morphol. 2020, 281, 1142–1151. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SH Strain | Recorded Period | Mother Age (Days) | Number of Pups | ||
|---|---|---|---|---|---|
| Total | Male | Female | |||
| HIS/Hiph | 2015–2021 | 97.8 ± 1.6 | 5.2 ± 0.1 (1–13) | 2.8 ± 0.1 (0–8) | 2.4 ± 0.2 (0–8) |
| HIS/Mz | 2019–2020 | 96.6 ± 2.7 | 5.7 ± 0.3 (1–10) | 3.0 ± 0.2 (1–7) | 2.7 ± 0.2 (0–6) |
| Follicle Type | Multi-Oocyte Follicles | |||||
|---|---|---|---|---|---|---|
| Two Oocytes (%) | Three Oocytes (%) | >Four Oocytes (%) | ||||
| HIS/Hiph | HIS/Mz | HIS/Hiph | HIS/Mz | HIS/Hiph | HIS/Mz | |
| Primordial | 52.7 | 82.0 | 43.3 | 16.0 | 4.0 | 2.0 |
| Primary | 62.5 | 94.5 | 35.0 | 5.5 | 2.5 | 0.0 |
| Secondary | 68.0 | 100.0 | 27.5 | 0.0 | 4.5 | 0.0 |
| Tertiary | 82.5 | 0.0 | 15.8 | 0.0 | 1.7 | 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Islam, M.R.; Ichii, O.; Nakamura, T.; Irie, T.; Shinohara, A.; Masum, M.A.; Otani, Y.; Namba, T.; Chuluunbaatar, T.; Elewa, Y.H.A.; et al. Comparison of Ovarian Morphology and Follicular Disturbances between Two Inbred Strains of Cotton Rats (Sigmodon hispidus). Animals 2021, 11, 1768. https://doi.org/10.3390/ani11061768
Islam MR, Ichii O, Nakamura T, Irie T, Shinohara A, Masum MA, Otani Y, Namba T, Chuluunbaatar T, Elewa YHA, et al. Comparison of Ovarian Morphology and Follicular Disturbances between Two Inbred Strains of Cotton Rats (Sigmodon hispidus). Animals. 2021; 11(6):1768. https://doi.org/10.3390/ani11061768
Chicago/Turabian StyleIslam, Md Rashedul, Osamu Ichii, Teppei Nakamura, Takao Irie, Akio Shinohara, Md Abdul Masum, Yuki Otani, Takashi Namba, Tsolmon Chuluunbaatar, Yaser Hosny Ali Elewa, and et al. 2021. "Comparison of Ovarian Morphology and Follicular Disturbances between Two Inbred Strains of Cotton Rats (Sigmodon hispidus)" Animals 11, no. 6: 1768. https://doi.org/10.3390/ani11061768
APA StyleIslam, M. R., Ichii, O., Nakamura, T., Irie, T., Shinohara, A., Masum, M. A., Otani, Y., Namba, T., Chuluunbaatar, T., Elewa, Y. H. A., & Kon, Y. (2021). Comparison of Ovarian Morphology and Follicular Disturbances between Two Inbred Strains of Cotton Rats (Sigmodon hispidus). Animals, 11(6), 1768. https://doi.org/10.3390/ani11061768