
Successful Use of Geochemical Tools to Trace the Geographic Origin of Long-Snouted Seahorse Hippocampus guttulatus Raised in Captivity


 ,
,  , ,
, ,  ,
,  and
and 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Biological Material
2.2. Samples Preparation
2.3. Elements Extraction and ICP-MS Analysis
2.4. Statistical Analysis
3. Results
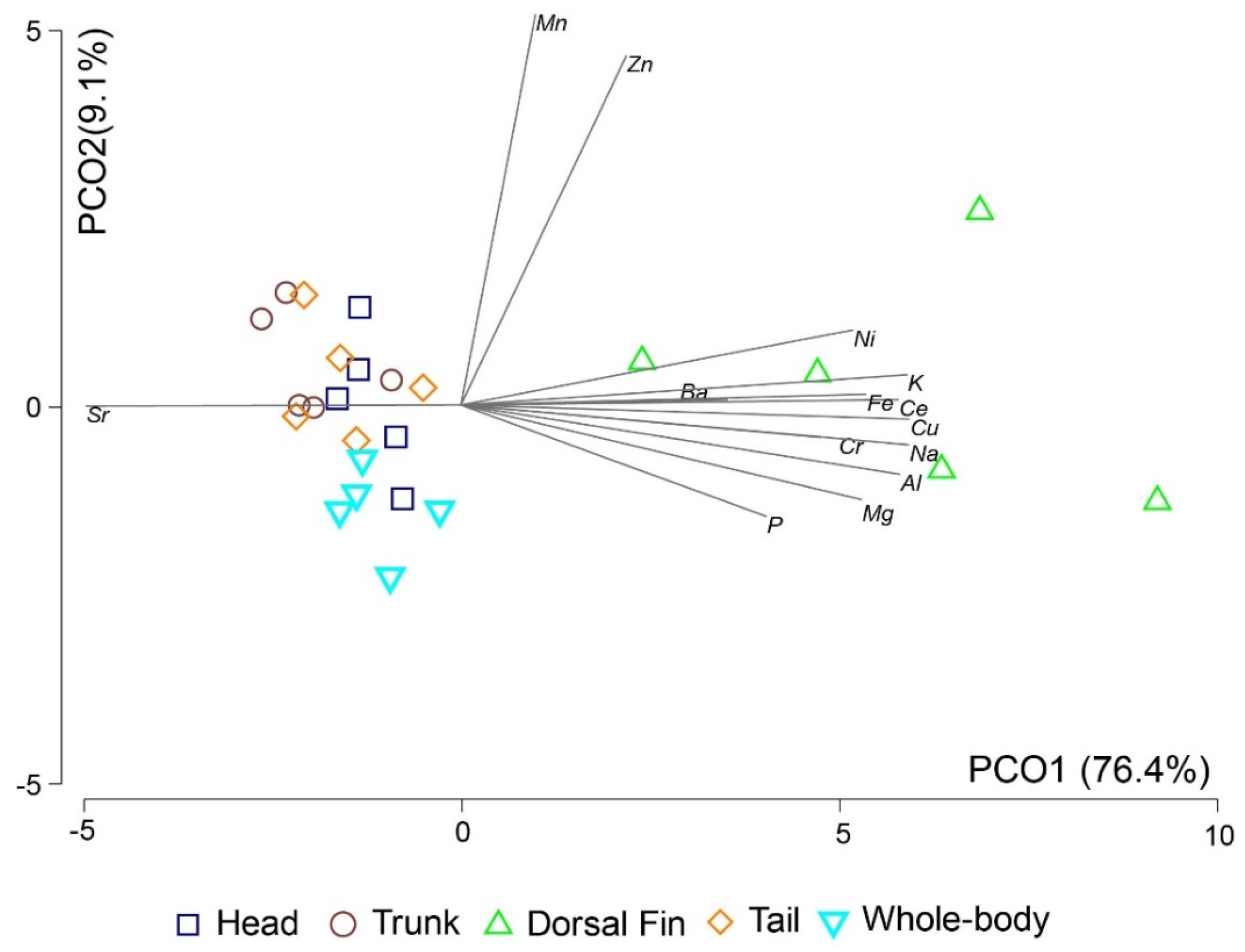
3.1. Assessment of a Proxy to the Whole Body
3.2. Traceability of the Geographic Origin
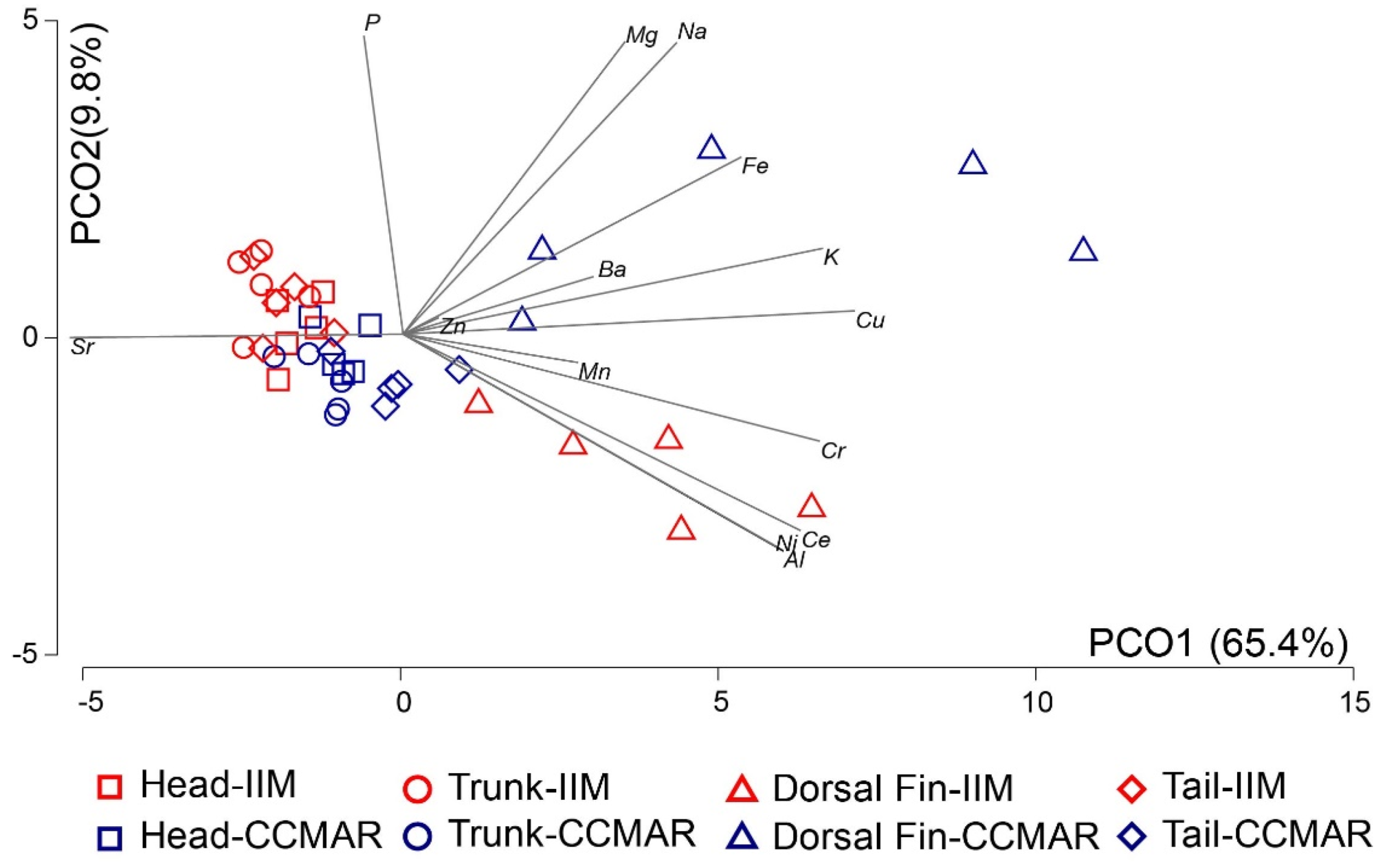
3.2.1. Elemental Composition of Seahorses from IIM and CCMAR
3.2.2. Body Parts as Blind Samples
3.2.3. Validation of the Trunk Elemental Fingerprint as a Model to Trace the Geographic Origin of Seahorses
3.2.4. Elemental Composition of Seahorse Body Portions
3.2.5. Seahorse Body Portions as Blind Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.Y.; Ryu, B.M.; Qian, Z.J. A review-biology, aquaculture and medical use of seahorse, Hippocampus spp. Annu. Res. Rev. Biol. 2017, 1–12. [Google Scholar] [CrossRef]
- Curtis, J.M.R.; Santos, S.V.; Nadeau, J.L.; Gunn, B.; Bigney Wilner, K.; Balasubramanian, H.; Overington, S.; Lesage, C.M.; D’Entremont, J.; Wieckowski, K. Life history and ecology of the elusive European short-snouted seahorse Hippocampus hippocampus. J. Biol. 2017, 91, 1603–1622. [Google Scholar] [CrossRef] [PubMed]
- Lourie, S. Measuring Seahorses; University of British Columbia Fisheries Centre: Vancouver, BC, Canada, 2003; p. 15. [Google Scholar]
- Porter, M.M.; Novitskaya, E.; Castro-Ceseña, A.B.; Meyers, M.A.; McKittrick, J. Highly deformable bones: Unusual deformation mechanisms of seahorse armor. Acta Biomater. 2013, 9, 6763–6770. [Google Scholar] [CrossRef] [PubMed]
- Lall, S.P.; Lewis-McCrea, L.M. Role of nutrients in skeletal metabolism and pathology in fish—An overview. Aquaculture 2007, 267, 3–19. [Google Scholar] [CrossRef]
- Cohen, F.P.; Valenti, W.C.; Planas, M.; Calado, R. Seahorse aquaculture, biology and conservation: Knowledge gaps and research opportunities. Rev. Fish. Sci. Aquac. 2017, 25, 100–111. [Google Scholar] [CrossRef]
- Faleiro, F.; Baptista, M.; Santos, C.; Aurélio, M.L.; Pimentel, M.; Pegado, M.R.; Paula, J.R.; Calado, R.; Repolho, T.; Rosa, R. Seahorses under a changing ocean: The impact of warming and acidification on the behaviour and physiology of a poor-swimming bony-armoured fish. Conserv. Physiol. 2015, 3, cov009. [Google Scholar] [CrossRef]
- Kuo, T.C.; Vincent, A. Assessing the changes in international trade of marine fishes under CITES regulations—A case study of seahorses. Mar. Policy 2018, 88, 48–57. [Google Scholar] [CrossRef]
- Aylesworth, L.; Phoonsawat, R.; Vincent, A.C. Effects of indiscriminate fisheries on a group of small data-poor species in Thailand. ICES J. Mar. Sci. 2018, 75, 642–652. [Google Scholar] [CrossRef]
- Vincent, A.C.; Foster, S.J.; Koldewey, H.J. Conservation and management of seahorses and other Syngnathidae. J. Fish. Biol. 2011, 78, 1681–1724. [Google Scholar] [CrossRef]
- Foster, S.; Wiswedel, S.; Vincent, A. Opportunities and challenges for analysis of wildlife trade using CITES data—Seahorses as a case study. Aquat. Conserv. 2016, 26, 154–172. [Google Scholar] [CrossRef]
- Cohen, F.P.; Valenti, W.C.; Calado, R. Traceability issues in the trade of marine ornamental species. Rev. Fish. Sci. Aquac. 2013, 21, 98–111. [Google Scholar] [CrossRef]
- Cohen, F.P.; Pimentel, T.; Valenti, W.C.; Calado, R. First insights on the bacterial fingerprints of live seahorse skin mucus and its relevance for traceability. Aquaculture 2018, 492, 259–264. [Google Scholar] [CrossRef]
- Leal, M.C.; Pimentel, T.; Ricardo, F.; Rosa, R.; Calado, R. Seafood traceability: Current needs, available tools, and biotechnological challenges for origin certification. Trends Biotechnol. 2015, 33, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Thorrold, S.R.; Latkoczy, C.; Swart, P.K.; Jones, C.M. Natal homing in a marine fish metapopulation. Science 2001, 291, 297–299. [Google Scholar] [CrossRef] [PubMed]
- Campana, S.E.; Chouinard, G.A.; Hanson, J.M.; Frechet, A.; Brattey, J. Otolith elemental fingerprints as biological tracers of fish stocks. Fish. Res. 2000, 46, 343–357. [Google Scholar] [CrossRef]
- Gillanders, B.M. Trace metals in four structures of fish and their use for estimates of stock structure. Fish. Bull. 2001, 99, 410. [Google Scholar]
- Luque, P.L.; Zhang, S.; Rooker, J.R.; Bidegain, G.; Rodríguez-Marín, E. Dorsal fin spines as a non-invasive alternative calcified structure for microelemental studies in Atlantic bluefin tuna. J. Exp. Mar. Biol. Ecol. 2017, 486, 127–133. [Google Scholar] [CrossRef]
- Ricardo, F.; Mamede, R.; Bispo, R.; Santos, A.; da Silva, E.F.; Patinha, C.; Calado, R. Cost-efficiency improvement of bivalves shells preparation when tracing their geographic origin through ICP-MS analysis of elemental fingerprints. Food Control 2020, 118, 107383. [Google Scholar] [CrossRef]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E Ltd.: Plymouth, UK, 2008. [Google Scholar]
- Ricardo, F.; Pimentel, T.; Génio, L.; Calado, R. Spatio-temporal variability of trace elements fingerprints in cockle (Cerastoderma edule) shells and its relevance for tracing geographic origin. Sci. Rep. 2017, 7, 3475. [Google Scholar] [CrossRef] [PubMed]
- Arechavala-Lopez, P.; Milošević-González, M.; Sanchez-Jerez, P. Using trace elements in otoliths to discriminate between wild and farmed European sea bass (Dicentrarchus labrax L.) and Gilthead sea bream (Sparus aurata L.). Int. Aquat. Res. 2016, 8, 263–273. [Google Scholar] [CrossRef]
- Novelli, B.; Otero-Ferrer, F.; Socorro, J.A.; Caballero, M.J.; Segade-Botella, A.; Domínguez, L.M. Development of short-snouted seahorse (Hippocampus hippocampus, L. 1758): Osteological and morphological aspects. Fish Physiol. Biochem. 2017, 43, 833–848. [Google Scholar] [CrossRef] [PubMed]
- Bruner, E.; Bartolino, V. Morphological Variation in the Seahorse Vertebral System. Int. J. Morphol. 2008, 26, 247–262. [Google Scholar] [CrossRef]
- Neutens, C.; De Dobbelaer, B.; Claes, P.; Adriaens, D. Prehensile and non-prehensile tails among syngnathid fishes: What’s the difference? Zoology 2017, 120, 62–72. [Google Scholar] [CrossRef]
- Praet, T.; Adriaens, D.; Cauter, S.V.; Masschaele, B.; Beule, M.D.; Verhegghe, B. Inspiration from nature: Dynamic modelling of the musculoskeletal structure of the seahorse tail. Int. J. Numer. Method Biomed. Eng. 2012, 28, 1028–1042. [Google Scholar] [CrossRef] [PubMed]
- Avigliano, E.; de Carvalho, B.M.; Miller, N.; Gironde, S.C.; Tombari, A.; Limburg, K.; Volpedo, A.V. Fin spine chemistry as a non-lethal alternative to otoliths for stock discrimination in an endangered catfish. Mar. Ecol. Prog. Ser. 2019, 614, 147–157. [Google Scholar] [CrossRef]
- Ramirez, M.D.; Miller, J.A.; Parks, E.; Avens, L.; Goshe, L.R.; Seminoff, J.A.; Snover, M.L.; Heppell, S.S. Reconstructing sea turtle ontogenetic habitat shifts through trace element analysis of bone tissue. Mar. Ecol. Prog. Ser. 2019, 608, 247–262. [Google Scholar] [CrossRef]
- Raubenheimer, E.J.; Brown, J.M.M.; Rama, D.B.K.; Dreyer, M.J.; Smith, P.D.; Dauth, J. Geographic variations in the composition of ivory of: The African elephant (Loxodonta africana). Arch. Oral Biol. 1998, 43, 641–647. [Google Scholar] [CrossRef]
- Avigliano, E.; Miller, N.; de Carvalho, B.M.; Gironde, S.C.; Tombari, A.; Volpedo, A.V. Fin spine metals by LA-ICP-MS as a method for fish stock discrimination of Genidens barbus in anthropized estuaries. Fish. Res. 2020, 230, 105625. [Google Scholar] [CrossRef]
- Kerr, L.A.; Campana, S.E. Chemical composition of fish hard parts as a natural marker of fish stocks. In Stock Identification Methods, 2nd ed.; Cadrin, S.X., Kerr, L.A., Mariani, S., Eds.; Academic Press: San Diego, CA, USA, 2014; pp. 205–234. [Google Scholar]
- Longmore, C.; Trueman, C.N.; Neat, F.; O’Gorman, E.J.; Milton, J.A.; Mariani, S. Otolith geochemistry indicates life-long spatial population structuring in a deep-sea fish, Coryphaenoides rupestris. Mar. Ecol. Prog. Ser. 2011, 435, 209–224. [Google Scholar] [CrossRef][Green Version]
- Clarke, L.M.; Thorrold, S.R.; Conover, D.O. Population differences in otolith chemistry have a genetic basis in Menidia menidia. Can. J. Fish. Aquat. Sci. 2011, 68, 105–114. [Google Scholar] [CrossRef]
- Campana, S.E. Otolith elemental composition as a natural marker of fish stocks. In Stock Identification Methods; Cadrin, S.X., Friedland, K.D., Waldman, J.R., Eds.; Elsevier Academic Press: Cambridge, MA, USA, 2005; pp. 227–245. [Google Scholar]
- Gillanders, B.M.; Kingsford, M.J. Spatial variation in elemental composition of otoliths of three species of fish (family Sparidae). Estuar. Coast. Shelf Sci. 2003, 57, 1049–1064. [Google Scholar] [CrossRef]
- Thorrold, S.R.; Jones, G.P.; Hellberg, M.E.; Burton, R.S.; Swearer, S.E.; Neigel, J.E.; Morgan, S.G.; Warner, R.R. Quantifying larval retention and connectivity in marine populations with artificial and natural markers. Bull. Mar. Sci. 2002, 70, 291–308. [Google Scholar]
- Aurélio, M.; Faleiro, F.; Lopes, V.M.; Pires, V.; Lopes, A.R.; Pimentel, M.S.; Repolho, T.; Baptista, M.; Narciso, L.; Rosa, R. Physiological and behavioral responses of temperate seahorses (Hippocampus guttulatus) to environmental warming. Mar. Biol. 2013, 160, 2663–2670. [Google Scholar] [CrossRef]
- Lourie, S. Fin-clipping procedure for seahorses. Proj. Seahorse Tech. Bull. 2003, 3, 1–4. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Pair-Wise Comparisons | Permanova | |
|---|---|---|
| t | p | |
| Head vs. Whole-body | 1.710 | 0.046 |
| Trunk vs. Whole-body | 2.486 | 0.004 |
| Dorsal fin vs. Whole-body | 4.163 | 0.001 |
| Tail vs. Whole-body | 2.030 | 0.013 |
| Pair-Wise Comparisons | Permanova | |
|---|---|---|
| t | p | |
| Head IIM vs. Head CCMAR | 1.440 | 0.130 |
| Trunk IIM vs. Trunk CCMAR | 3.565 | <0.001 |
| Dorsal Fin IIM vs. Dorsal Fin CCMAR | 1.863 | 0.039 |
| Tail IIM vs. Tail CCMAR | 2.656 | 0.002 |
| Origin of Sample | IIM (%) | CCMAR (%) |
|---|---|---|
| IIM-Head | 100 | 0 |
| IIM-Trunk | 100 | 0 |
| IIM-Dorsal Fin | 0 | 100 |
| IIM-Tail | 100 | 0 |
| CCMAR-Head | 60 | 40 |
| CCMAR-Trunk | 0 | 100 |
| CCMAR-Dorsal Fin | 0 | 100 |
| CCMAR-Tail | 40 | 60 |
| Pair-Wise Comparions | Permanova | |
|---|---|---|
| t | p | |
| H + T + DF IIM vs. H + T + DF CCMAR | 2.507 | 0.003 |
| T + DF IIM vs. T + DF CCMAR | 2.488 | 0.004 |
| T + TL IIM vs. T + TL CCMAR | 2.664 | 0.003 |
| T + DF + TL IIM vs. T + DF + TL CCMAR | 2.694 | 0.002 |
| H + T IIM vs. H + T CCMAR | 2.368 | 0.004 |
| H + T + TL IIM vs. H + T + TL CCMAR | 2.400 | 0.006 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cabral, A.E.; Ricardo, F.; Patinha, C.; Silva, E.F.d.; Correia, M.; Palma, J.; Planas, M.; Calado, R. Successful Use of Geochemical Tools to Trace the Geographic Origin of Long-Snouted Seahorse Hippocampus guttulatus Raised in Captivity. Animals 2021, 11, 1534. https://doi.org/10.3390/ani11061534
Cabral AE, Ricardo F, Patinha C, Silva EFd, Correia M, Palma J, Planas M, Calado R. Successful Use of Geochemical Tools to Trace the Geographic Origin of Long-Snouted Seahorse Hippocampus guttulatus Raised in Captivity. Animals. 2021; 11(6):1534. https://doi.org/10.3390/ani11061534
Chicago/Turabian StyleCabral, Ana Elisa, Fernando Ricardo, Carla Patinha, Eduardo Ferreira da Silva, Miguel Correia, Jorge Palma, Miquel Planas, and Ricardo Calado. 2021. "Successful Use of Geochemical Tools to Trace the Geographic Origin of Long-Snouted Seahorse Hippocampus guttulatus Raised in Captivity" Animals 11, no. 6: 1534. https://doi.org/10.3390/ani11061534
APA StyleCabral, A. E., Ricardo, F., Patinha, C., Silva, E. F. d., Correia, M., Palma, J., Planas, M., & Calado, R. (2021). Successful Use of Geochemical Tools to Trace the Geographic Origin of Long-Snouted Seahorse Hippocampus guttulatus Raised in Captivity. Animals, 11(6), 1534. https://doi.org/10.3390/ani11061534