
Effects of Flavonoids Extracted from Citrus aurantium on Performance, Behavior, and Rumen Gene Expression in Holstein Bulls Fed with High-Concentrate Diets in Pellet Form


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Feeding, Housing, and Experimental Design
2.2. Feed Consumption and Performance
2.3. Animal Behavior
2.4. Carcass Quality
2.5. Rumen and Liver Macroscopic Evaluation and Sample Collection
2.6. Biological and Chemical Analyses
2.7. Calculations and Statistical Analyses
3. Results
3.1. Animal Health
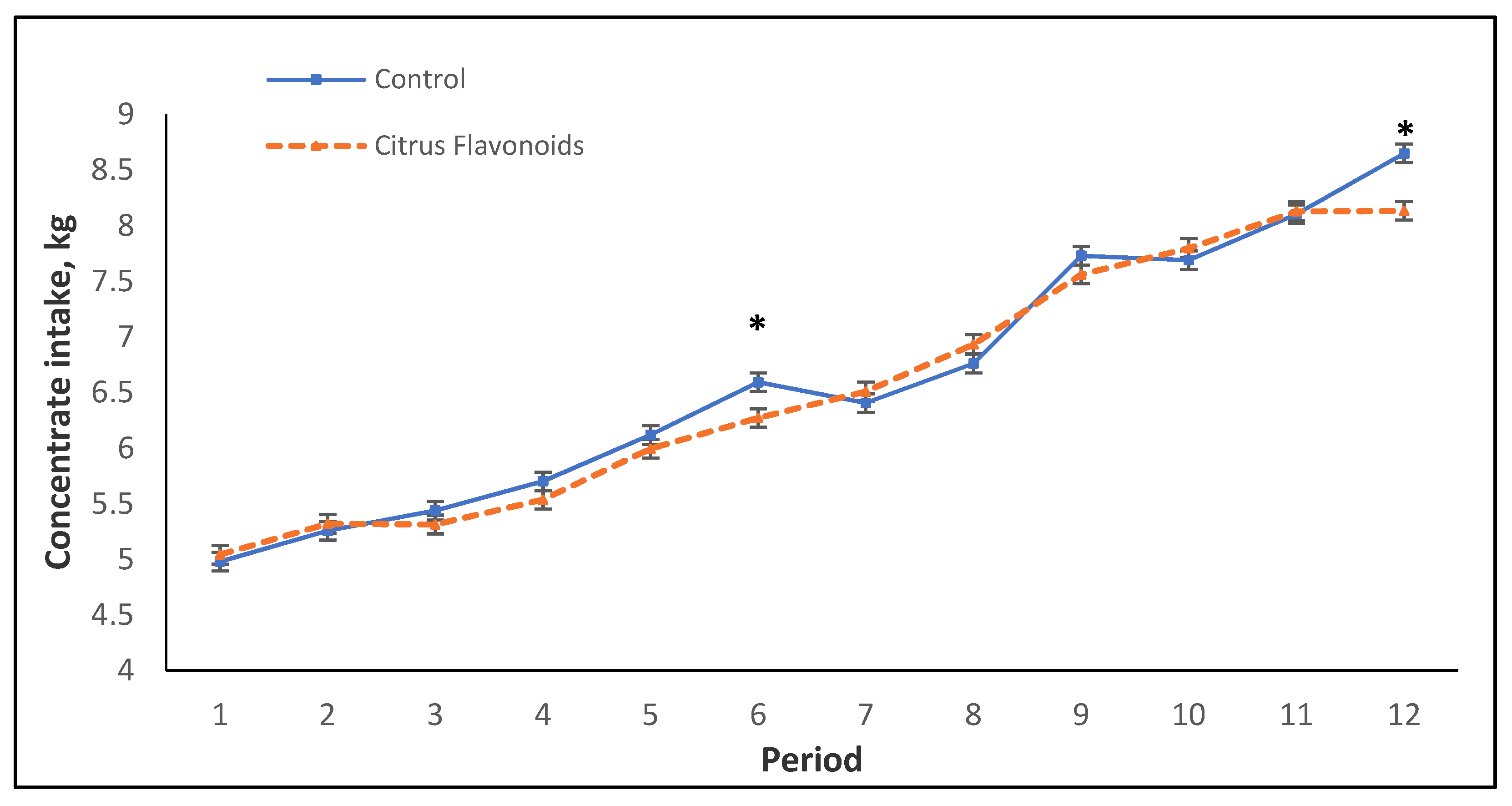
3.2. Intake, Performance, and Carcass Quality
3.3. Animal Behavior
3.3.1. General Activities
3.3.2. Active Behavior
3.4. Macroscopic Rumen Evaluation and Liver Abscesses
3.5. Rumen pH and VFA Concentration at Slaughterhouse
3.6. Expression of Genes in the Rumen Epithelium
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Durmic, Z.; Blache, D. Bioactive plants and plant products: Effects on animal function, health and welfare. Anim. Feed Sci. Technol. 2012, 176, 150–162. [Google Scholar] [CrossRef]
- Balcells, J.; Aris, A.; Serrano, A.; Seradj, A.R.; Crespo, J.; Devant, M. Effects of an extract of plant flavonoids (Bioflavex) on rumen fermentation and performance in heifers fed high-concentrate diets. J. Anim. Sci. 2012, 90, 4975–4984. [Google Scholar] [CrossRef] [PubMed]
- Seradj, A.R.; Abecia, L.; Crespo, J.; Villalba, D.; Fondevila, M.; Balcells, J. The effect of Bioflavex® and its pure flavonoid components on in vitro fermentation parameters and methane production in rumen fluid from steers given high concentrate diets. Anim. Feed Sci. Technol. 2014, 197, 85–91. [Google Scholar] [CrossRef]
- Paniagua, M.; Crespo, J.; Bach, A.; Devant, M. Effects of flavonoids extracted from Citrus aurantium on performance, eating and animal behavior, rumen health, and carcass quality in Holstein bulls fed high-concentrate diets. Anim. Feed Sci. Technol. 2018, 246, 114–126. [Google Scholar] [CrossRef]
- Paniagua, M.; Crespo, J.; Arís, A.; Devant, M. Citrus aurantium flavonoid extract improves concentrate efficiency, animal behavior, and reduces rumen inflammation of Holstein bulls fed high-concentrate diets. Anim. Feed Sci. Technol. 2019, 258, 114–304. [Google Scholar] [CrossRef]
- Devant, M.; Penner, G.B.; Marti, S.; Quintana, B.; Fábregas, F.; Bach, A.; Arís, A. Behavior and inflammation of the rumen and cecum in Holstein bulls fed high-concentrate diets with different concentrate presentation forms with or without straw supplementation. J. Anim. Sci. 2016, 94, 3902–3917. [Google Scholar] [CrossRef]
- Verdú, M.; Bach, A.; Devant, M. Effect of feeder design and concentrate presentation form on performance, carcass characteristics, and behavior of fattening Holstein bulls fed high-concentrate diets. Anim. Feed Sci. Technol. 2017, 232, 148–159. [Google Scholar] [CrossRef]
- Ferret, A.; Calsamiglia, S.; Bach, A.; Devant, M.; Fernández, C.; García-Rebollar, P. Necesidades Nutricionales Para Rumiantes de Cebo Normas FEDNA; Fundación Española para el Desarrollo de la Nutrición Animal: Madrid, Spain, 2008. [Google Scholar]
- González, J.M.; Garcia de Jalón, J.A.; Askar, A.R.; Guada, J.A.; Ferrer, L.M.; de las Heras, M. Efecto de la dieta con cebada y nucleo proteico sobre la patología ruminal en corderos. XXVI J. SEOC Junta Andal. 2001, 733–739. [Google Scholar]
- Lesmeister, K.E.; Tozer, P.R.; Heinrichs, A.J. Development and analysis of a rumen tissue sampling procedure. J. Dairy Sci. 2004, 87, 1336–1344. [Google Scholar] [CrossRef]
- Nocek, J.E.; Heald, C.W.; Polan, C.E. Influence of ration physical form and nitrogen availability on ruminal morphology of growing bull calves. J. Dairy Sci. 1984, 67, 334–343. [Google Scholar] [CrossRef]
- Brown, H.; Bing, R.F.; Grueter, H.P.; McAskill, J.W.; Cooley, C.O.; Rathmacher, R.P. Tylosin and chloretetracycline for the prevention of liver abscesses, improved weight gains and feed efficiency in feedlot cattle. J. Anim. Sci. 1975, 40, 207–213. [Google Scholar] [CrossRef]
- Jouany, J.P. Volatile fatty acids and alcohol determination in digestive contents, silage juice, bacterial cultures and anaerobic fermentor contents. Sci. Alim. 1982, 2, 131–144. [Google Scholar]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis, 17th ed.; AOAC: Arlington, VA, USA, 2005; Volume 2. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Amann, R.P.; Walker, O.A. Changes in the pituitary-gonadal axis associated with puberty in Holstein bulls. J. Anim. Sci. 1983, 57, 433–442. [Google Scholar] [CrossRef]
- Kenny, D.A.; Byrne, C.J. Review: The effect of nutrition on timing of pubertal onset and subsequent fertility in the bull. Animal 2018, 12, s36–s44. [Google Scholar] [CrossRef]
- Bergeron, R.; Badnell-Waters, A.J.; Lambton, S.; Mason, G.J. Stereotypic Animal Behaviour: Fundamentals and Applications to Welfare, 2nd ed.; Manson, G., Rishen, J., Eds.; CAB International: Wallingford, UK, 2006; pp. 19–57. ISBN 1-84593-042-8. [Google Scholar]
- Evans, J.M.; Morris, L.S.; Marchesi, J.R. The gut microbiome: The role of a virtual organ in the endocrinology of the host. J. Endocrinol. 2013, 218, R37–R47. [Google Scholar] [CrossRef]
- Haagensen, A.M.J.; Sørensen, D.B.; Sandøe, P.; Matthews, L.R.; Birck, M.M.; Fels, J.J.; Astrup, A. High Fat, Low Carbohydrate Diet Limit Fear and Aggression in Göttingen Minipigs. PLoS ONE 2014, 9, e93821. [Google Scholar] [CrossRef]
- Harborne, J.B.; Williams, C.A. Advances in flavonoid research since 1992. Phytochem 2000, 55, 481–504. [Google Scholar] [CrossRef]
- Qaisrani, S.N.; van Krimpen, M.M.; Kwakkel, R.P. Effects of dietary dilution source and dilution level on feather damage, performance, behavior, and litter condition in pullets. Poult. Sci. 2013, 92, 591–602. [Google Scholar] [CrossRef] [PubMed]
- Depoortere, I. Taste receptors of the gut: Emerging roles in health and disease. Gut 2013, 63, 179–190. [Google Scholar] [CrossRef]
- Chen, M.C.; Wu, S.V.; Reeve, J.R.; Rozengurt, E.; Monica, C.; Wu, S.V.; Reeve, J.R. Bitter stimuli induce Ca 2+ signaling and CCK release in enteroendocrine STC-1 cells: Role of L-type voltage-sensitive Ca 2+ channels. Am. J. Physiol. 2006, 1786, 726–739. [Google Scholar] [CrossRef]
- Takai, S.; Yoshida, R.; Shigemura, N.; Ninomiya, Y. Chemosensory Transduction, 1st ed.; Zufall, F., Munger, S.D., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 299–317. ISBN 978-0-12-801694-7. [Google Scholar]
- Dewnowski, A.; Ahlstrom, S.; Shore, A.B. Taste responses to naringin, a flavonoid, and the acceptance of grapefruit juice are related to genetic sensitivity to 6-n-propylthiouracil. Am. J. Clin. Nutr. 1997, 66, 391–397. [Google Scholar] [CrossRef]
- Roland, W.S.U. Intrinsic Bitterness of Flavonoids and Isoflavonoids and Masking of Their Taste Activity. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 2014. [Google Scholar]
- Bufe, B.; Hofmann, T.; Krautwurst, D.; Raguse, J.; Meyerhof, W. The human TAS2R16 receptor mediates bitter taste in response to β -glucopyranosides. Nat. Genet. 2002, 32, 397–401. [Google Scholar] [CrossRef]
- Ji, M.; Su, X.; Su, X.; Huang, W.; Zhang, J.; Li, C.; Lu, X. Identification of Novel Compounds for Human Bitter Taste Receptors. Chem. Biol. Drug Des. 2014, 84, 63–74. [Google Scholar] [CrossRef]
- Tripoli, E.; La Guardia, M.; Giammanco, S.; Di Majo, D.; Giammanco, M. Food Chemistry Citrus flavonoids: Molecular structure, biological activity and nutritional properties: A review. Food Chem. J. 2007, 104, 466–479. [Google Scholar] [CrossRef]
- Larhammar, D. Structural diversity of receptors for neuropeptide Y, peptide YY and pancreatic polypeptide. Regul. Pept. 1996, 65, 165–174. [Google Scholar] [CrossRef]
- Hassan, F.; Adeel Arhshad, M.; Li, M.; Saif-ur Rehman, M.; Loor, J.J.; Huang, J. Potencial of mulberry lead biomass and its flavonoids to improve production and health in ruminants: Mechanistics insights and prospects. Animals 2020, 10, 2076. [Google Scholar] [CrossRef] [PubMed]
- Simpson, F.; Jones, G.; Wolin, E. Anaerobic degradation of some bioflavonoids by microflora of the rumen. Can. J. Microbiol. 1969, 15, 972–974. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.J.; Krishnamurty, H.G.; Jones, G.A.; Simpson, F.J. Identification of products produced by the anaerobic degradation of naringin by Butyrivibrio sp. C3. Can. J. Microbiol. 1971, 17, 129–131. [Google Scholar] [CrossRef] [PubMed]
- Bertipaglia, L.M.A.; Fondevila, M.; Laar, H.V.; Castrillo, C. Effect of pelleting and pellet size of a concentrate for intensively reared beef cattle on in vitro fermentation by two different approaches. Anim. Feed Sci. Technol. 2010, 159, 88–95. [Google Scholar] [CrossRef]
- Hudson, B.D.; Christiansen, E.; Tikhonova, I.G.; Grundmann, M.; Kostenis, E.; Adams, D.R.; Ulven, T.; Milligan, G.; Chemically, G. Chemically engineering ligand selectivity at the free fatty acid receptor 2 based on pharmacological variation between species orthologs. FASEB J. 2012, 26, 4951–4965. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Item | ||
|---|---|---|
| Ingredient, g/kg | Growing | Finishing |
| Corn grain meal | 399.7 | 450.9 |
| Barley grain meal | 179.8 | 155.5 |
| Dried distillers grains | 179.8 | 150.2 |
| Wheat | 109.7 | 110.3 |
| Beet pulp | 73.9 | 80.2 |
| Palm oil | 20.0 | 25.0 |
| Calcium carbonate | 15.5 | 12.8 |
| Urea | 8.0 | 4.0 |
| Sodium bicarbonate | 5.0 | 4.0 |
| Dicalcium phosphate | 3.6 | 3.1 |
| Vitamin premix | 3.0 | 2.0 |
| Salt | 2.0 | 2.0 |
| Nutrient, per kg DM | ||
| ME, Mcal | 3.30 | 2.97 |
| CP, g | 157 | 123 |
| Ether extract, g | 58 | 54 |
| Ash, g | 56 | 44 |
| NDF, g | 178 | 151 |
| Non-fiber carbohydrates, g | 550 | 628 |
| Treatment 1 | p-Value 2 | |||||
|---|---|---|---|---|---|---|
| Item | Control | BF | SEM | T | Time | T × Time |
| Growing phase | ||||||
| Initial age, d | 137.3 | 137.3 | 1.81 | ns | ||
| Final age, d | 259.3 | 249.3 | 1.74 | ns | ||
| Initial BW, kg | 182.8 | 183.2 | 7.53 | ns | ||
| Final BW (112 d of study) | ||||||
| Mean, kg | 344.5 | 346.3 | 7.72 | ns | ||
| CV, % | 8.1 | 7.9 | 0.45 | ns | ||
| ADG, kg/d | ||||||
| Mean, kg/d | 1.45 | 1.45 | 0.018 | ns | *** | ns |
| CV, % | 24.6 | 26.8 | 2.18 | ns | t | ns |
| Concentrate DM intake | ||||||
| Mean, kg/d | 5.9 | 5.8 | 0.09 | ns | *** | * |
| CV, % | 13.8 | 13.2 | 0.76 | ns | *** | ns |
| FCR, kg/kg | 4.1 | 4.1 | 0.05 | ns | ** | ns |
| Finishing Phase | ||||||
| Initial age, d | 259.3 | 249.3 | 1.74 | ns | ||
| Final age, d | 306.3 | 306.3 | 1.74 | ns | ||
| Initial BW, kg | 344.6 | 346.3 | 7.72 | ns | ||
| Final BW (112 d of study), | ||||||
| Mean, kg | 433.9 | 435.9 | 7.60 | ns | ||
| CV, % | 7.9 | 7.1 | 0.59 | ns | ||
| ADG, kg/d | ||||||
| Mean, kg/d | 1.56 | 1.55 | 0.026 | ns | ns | ns |
| CV, % | 33.6 | 32.6 | 3.23 | ns | ns | ns |
| Concentrate DM intake | ||||||
| Mean, kg/d | 8.0 | 7.9 | 0.11 | ns | *** | ** |
| CV, % | 13.1 | 12.4 | 0.99 | ns | t | * |
| FCR, kg/kg | 5.2 | 5.1 | 0.12 | ns | ns | ns |
| Whole study | ||||||
| Initial age, d | 137.2 | 137.3 | 1.81 | ns | ||
| Final age, d | 306.3 | 306.3 | 1.73 | ns | ||
| Initial BW, kg | 182.8 | 183.2 | 7.53 | ns | ||
| Final BW (168 d of study), | ||||||
| Mean, kg | 433.9 | 435.9 | 7.60 | ns | ||
| CV, % | 7.9 | 7.1 | 0.59 | ns | ||
| ADG, kg/d | ns | |||||
| Mean, kg/d | 1.48 | 1.48 | 0.018 | ns | *** | ns |
| CV, % | 27.6 | 28.8 | 1.97 | ns | ** | ns |
| Concentrate DM intake | ns | |||||
| Mean, kg/d | 6.6 | 6.5 | 0.09 | ns | *** | ** |
| CV, % | 13.6 | 13.0 | 0.59 | ns | *** | ns |
| FCR, kg/kg | 4.5 | 4.5 | 0.07 | ns | *** | ns |
| Item | Treatment 1 | p-Values 2 | ||||
|---|---|---|---|---|---|---|
| Control | BF | SEM 3 | T | Time | T × Time | |
| General Activity, % | ||||||
| Standing | 78.46 | 77.79 | 0.058 | ns | *** | ns |
| Lying | 19.66 | 20.33 | 0.191 | ns | *** | ns |
| Eating concentrate | 7.64 | 9.99 | 0.060 | ** | * | ns |
| Eating straw | 11.51 | 12.81 | 0.025 | ns | *** | ns |
| Drinking | 1.55 | 2.63 | 0.003 | ns | ns | ns |
| Ruminating | 7.66 | 11.37 | 0.082 | ** | * | ns |
| Social behavior, /15 min | ||||||
| Self-grooming | 19.32 | 28.95 | 0.102 | *** | ns | ns |
| Social | 1.96 | 4.02 | 0.126 | ** | ns | ns |
| Oral non-nutritive | 0.77 | 0.59 | 0.049 | ns | ** | ns |
| Fighting | 9.16 | 4.91 | 0.346 | 0.06 | *** | ns |
| Butting | 2.30 | 1.18 | 0.098 | ** | *** | ns |
| Displacement | 0.464 | 0.214 | 0.033 | *** | ns | ns |
| Chasing | 0.70 | 0.20 | 0.071 | * | * | 0.06 |
| Chasing up | 0.16 | 0.05 | 0.029 | ns | ns | ns |
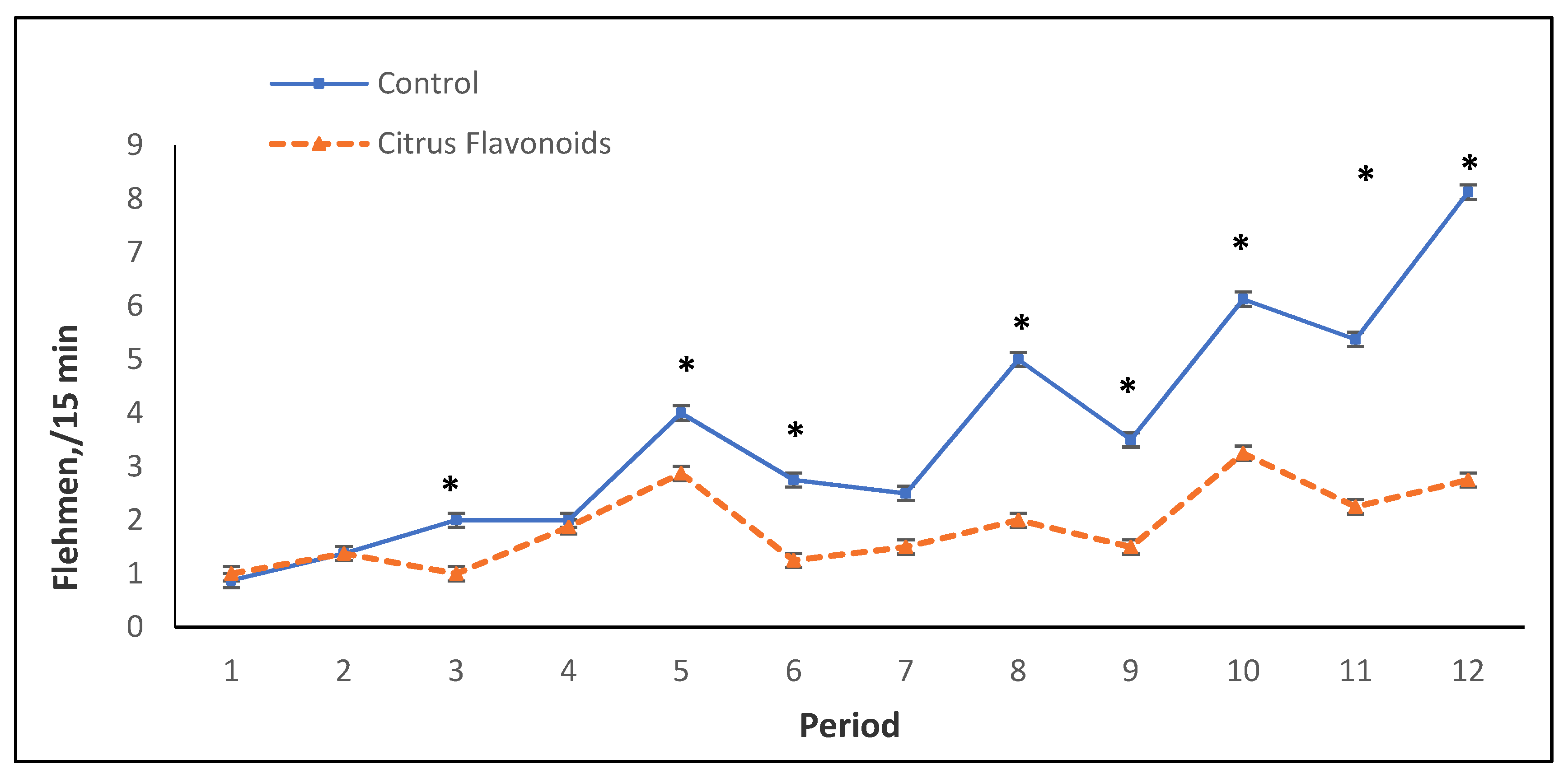
| Flehmen | 2.21 | 1.55 | 0.091 | * | ** | ns |
| Attempt to mount | 2.21 | 1.44 | 0.153 | ns | ** | ns |
| Complete mounts | 2.55 | 2.00 | 0.091 | ns | * | ns |
| Item | Treatment 1 | p-Values 2 | ||||
|---|---|---|---|---|---|---|
| Control | BF | SEM 3 | T | Time | T × Time | |
| General Activity, % | ||||||
| Standing | 67.14 | 66.41 | 0.080 | ns | ** | ns |
| Lying | 32.86 | 32.93 | 0.168 | ns | ** | ns |
| Eating concentrate | 6.27 | 8.10 | 0.060 | ns | ns | ns |
| Eating straw | 7.86 | 8.67 | 0.065 | ns | *** | ns |
| Drinking | 2.14 | 2.54 | 0.004 | ns | ns | ns |
| Ruminating | 8.12 | 14.03 | 0.072 | ** | ns | ns |
| Social behavior, /15 min | ||||||
| Self-grooming | 10.20 | 16.90 | 0.150 | *** | *** | ns |
| Social | 3.85 | 5.93 | 0.210 | * | ns | ns |
| Oral non-nutritive | 1.38 | 0.78 | 0.067 | t | ** | ns |
| Fighting | 8.85 | 3.85 | 0.256 | ** | ** | ns |
| Butting | 4.18 | 1.60 | 0.214 | * | ns | ns |
| Displacement | 1.00 | 0.15 | 0.070 | *** | ns | ns |
| Chasing | 0.56 | 0.08 | 0.050 | * | ns | ns |
| Chasing up | 0.23 | 0.05 | 0.032 | ns | ns | ns |
| Flehmen | 5.63 | 2.35 | 0.079 | *** | ** | ns |
| Attempt to mount | 1.45 | 0.10 | 0.228 | t | t | ns |
| Complete mounts | 2.80 | 1.58 | 0.165 | ns | 0.06 | ns |
| Treatment 1 | p-Value 2 | ||
|---|---|---|---|
| Item | C | BF | |
| Color of the rumen 3 | * | ||
| 2 | 2.67 | 2.86 | |
| 3 | 48.00 | 70.00 | |
| 4 | 48.00 | 24.29 | |
| 5 | 1.33 | 2.86 | |
| Papillae clumping 4 | ns | ||
| Yes | 40.00 | 44.29 | |
| No | 60.00 | 55.71 | |
| Baldness region 4 | ns | ||
| Yes | 40.00 | 49.33 | |
| No | 60.00 | 50.67 | |
| Liver abscess 5 | ns | ||
| None | 90.67 | 94.29 | |
| A | 2.67 | 1.43 | |
| A- | 4.00 | 2.86 | |
| A+ | 1.33 | - | |
| Inflammation | 1.33 | 1.43 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paniagua, M.; Crespo, F.J.; Arís, A.; Devant, M. Effects of Flavonoids Extracted from Citrus aurantium on Performance, Behavior, and Rumen Gene Expression in Holstein Bulls Fed with High-Concentrate Diets in Pellet Form. Animals 2021, 11, 1387. https://doi.org/10.3390/ani11051387
Paniagua M, Crespo FJ, Arís A, Devant M. Effects of Flavonoids Extracted from Citrus aurantium on Performance, Behavior, and Rumen Gene Expression in Holstein Bulls Fed with High-Concentrate Diets in Pellet Form. Animals. 2021; 11(5):1387. https://doi.org/10.3390/ani11051387
Chicago/Turabian StylePaniagua, Montserrat, Francisco Javier Crespo, Anna Arís, and Maria Devant. 2021. "Effects of Flavonoids Extracted from Citrus aurantium on Performance, Behavior, and Rumen Gene Expression in Holstein Bulls Fed with High-Concentrate Diets in Pellet Form" Animals 11, no. 5: 1387. https://doi.org/10.3390/ani11051387
APA StylePaniagua, M., Crespo, F. J., Arís, A., & Devant, M. (2021). Effects of Flavonoids Extracted from Citrus aurantium on Performance, Behavior, and Rumen Gene Expression in Holstein Bulls Fed with High-Concentrate Diets in Pellet Form. Animals, 11(5), 1387. https://doi.org/10.3390/ani11051387