
Effects of Medicinal Plants and Organic Selenium against Ovine Haemonchosis

 , , , , , , , , and
, , , , , , , , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals, Diets, and Experimental Design
2.3. Chemical Composition of Dietary Substrates
2.4. Animal Weighing and Parasitological Analyses
2.5. Hematological Analyses
2.6. Statistical Analysis
3. Results
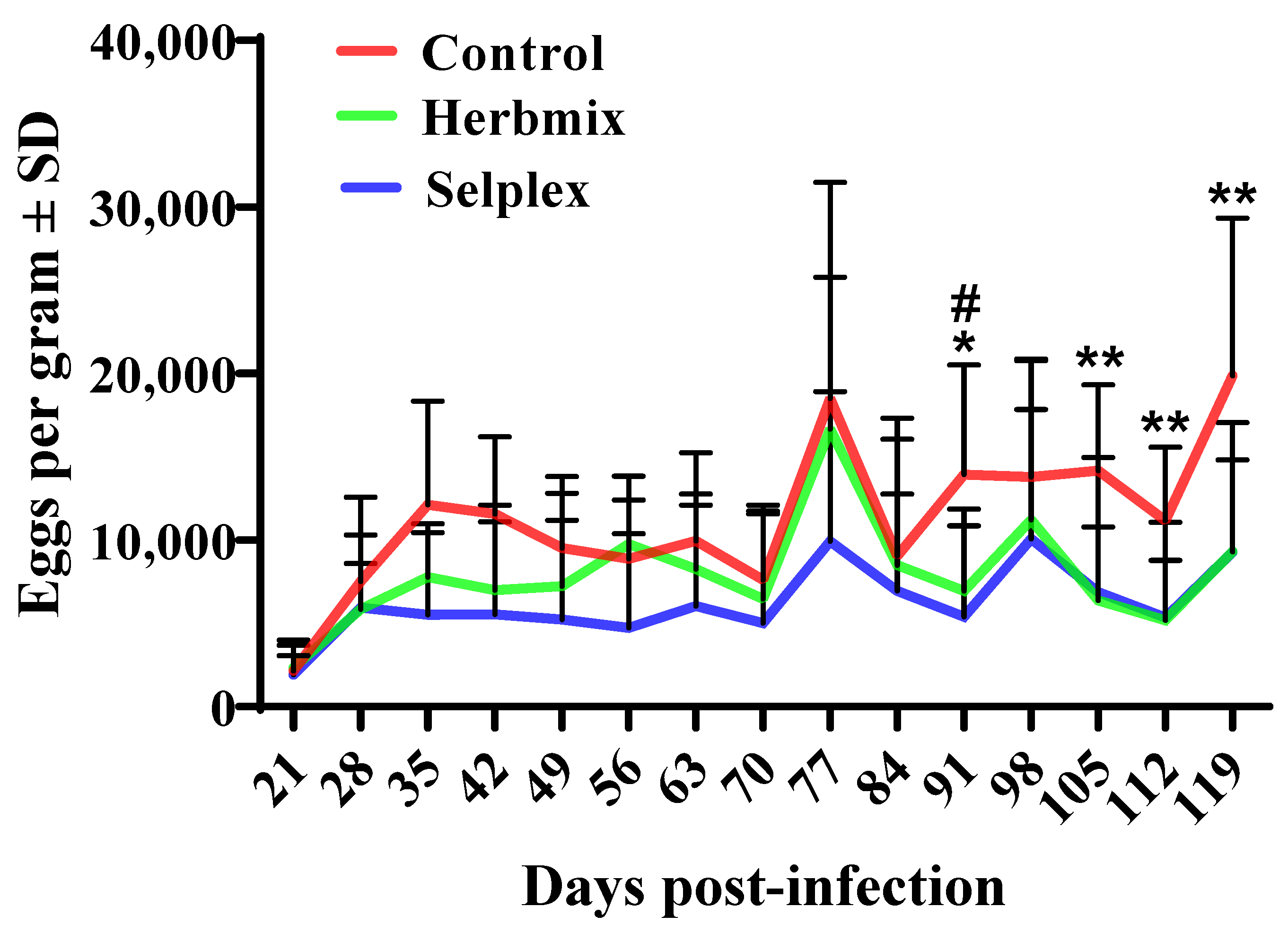
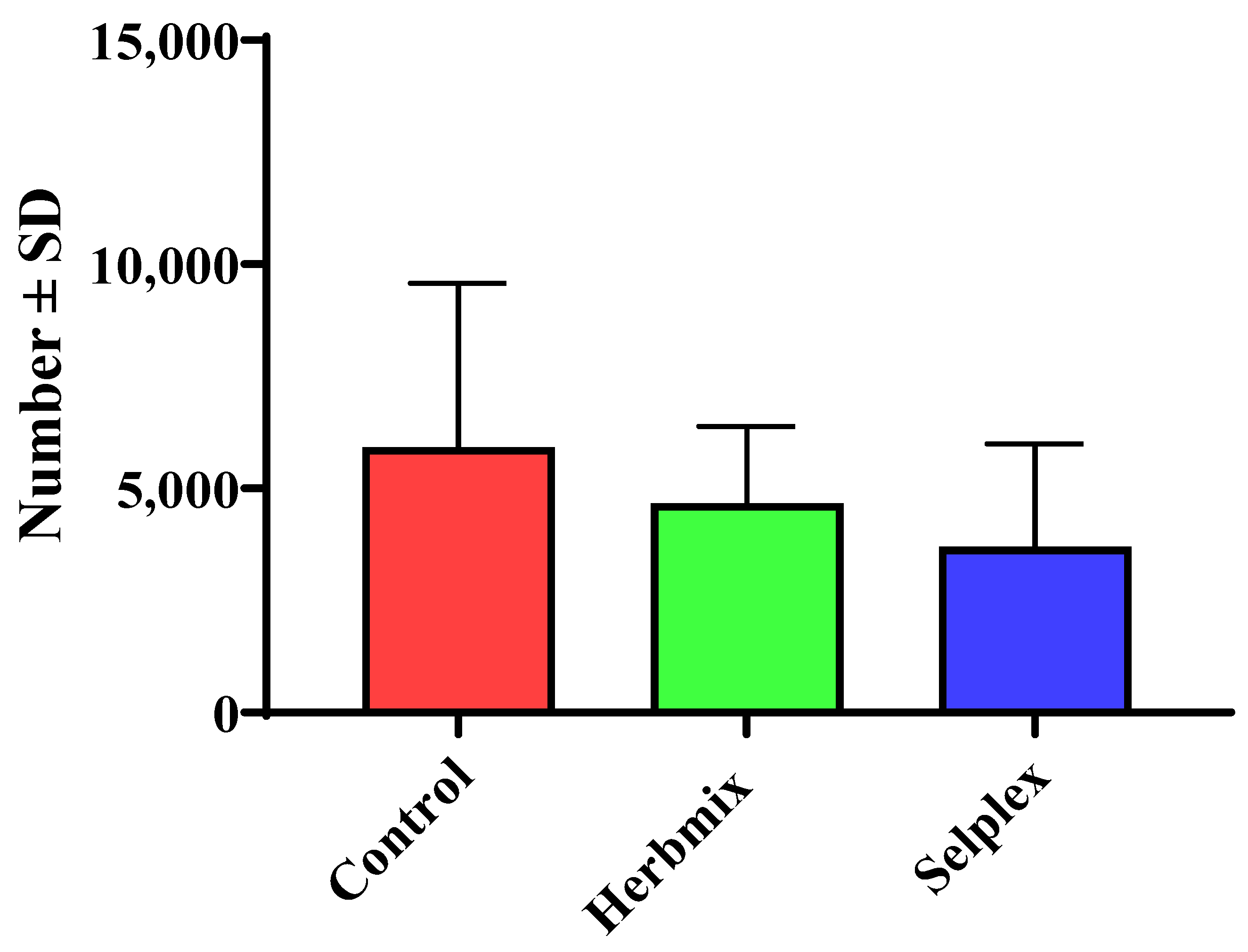
3.1. Weight and Parasitological Status
3.2. Red Blood Cell Hemograms
3.3. Vitamin E
3.4. Antioxidant Status
3.5. Triiodothyronine and Thyroxine
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wolstenholme, A.J.; Fairweather, I.; Prichard, R.; von Samson-Himmelstjerna, G.; Sangster, N.C. Drug resistance in veterinary helminths. Trends Parasitol. 2004, 20, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Waghorn, T.S.; Leathwick, D.M.; Rhodes, A.P.; Lawrence, K.E.; Jackson, R.; Pomroy, W.E.; West, D.M.; Moffat, J.R. Prevalence of anthelmintic resistance on sheep farms in New Zealand. N. Z. Vet. J. 2006, 54, 271–277. [Google Scholar] [CrossRef]
- Zajíčková, M.; Nguyen, L.T.; Skálová, L.; Raisová Stuchlíková, L.; Matoušková, P. Anthelmintics in the future: Current trends in the discovery and development of new drugs against gastrointestinal nematodes. Drug Discov. Today 2020, 25, 430–437. [Google Scholar] [CrossRef]
- Hoste, H.; Torres-Acosta, J.F.J.; Quijada, J.; Chan-Perez, I.; Dakheel, M.M.; Kommuru, D.S.; Mueller-Harvey, I.; Terrill, T.H. Interactions between nutrition and infections with Haemonchus contortus and related gastrointestinal nematodes in small ruminants. Adv. Parasitol. 2016, 93, 239–351. [Google Scholar] [CrossRef] [PubMed]
- Hoste, H.; Torres-Acosta, J.F.; Sandoval-Castro, C.A.; Mueller-Harvey, I.; Sotiraki, S.; Louvandini, H.; Thamsborg, S.M.; Terrill, T.H. Tannin containing legumes as a model for nutraceuticals against digestive parasites in livestock. Vet. Parasitol. 2015, 212, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Torres-Fajardo, R.A.; González-Pech, P.G.; Sandoval-Castro, C.A.; Torres-Acosta, J.F.J. Small ruminant production based on rangelands to optimize animal nutrition and health: Building an interdisciplinary approach to evaluate nutraceutical plants. Animals 2020, 10, 1799. [Google Scholar] [CrossRef]
- Váradyová, Z.; Kišidayová, S.; Čobanová, K.; Grešáková, Ľ.; Babják, M.; Königová, A.; Urda Dolinská, M.; Várady, M. The impact of a mixture of medicinal herbs on ruminal fermentation, parasitological status and hematological parameters of the lambs experimentally infected with Haemonchus contortus. Small Rum. Res. 2017, 151, 124–132. [Google Scholar] [CrossRef]
- Váradyová, Z.; Mravčáková, D.; Babják, M.; Bryszak, M.; Grešáková, Ľ.; Čobanová, K.; Kišidayová, S.; Plachá, I.; Königová, A.; Cieslak, A.; et al. Effects of herbal nutraceuticals and/or zinc against Haemonchus contortus in lambs experimentally infected. BMC Vet. Res. 2018, 14, 78. [Google Scholar] [CrossRef] [PubMed]
- Mravčáková, D.; Váradyová, Z.; Kopčáková, A.; Čobanová, K.; Grešáková, Ľ.; Kišidayová, S.; Babják, M.; Urda Dolinská, M.; Dvorožňáková, E.; Königová, A.; et al. Natural chemotherapeutic alternatives for controlling of haemonchosis in sheep. BMC Vet. Res. 2019, 15, 302. [Google Scholar] [CrossRef]
- Mravčáková, D.; Komáromyová, M.; Babják, M.; Urda Dolinská, M.; Königová, A.; Petrič, D.; Čobanová, K.; Ślusarczyk, S.; Cieslak, A.; Várady, M.; et al. Anthelmintic activity of wormwood (Artemisia absinthium L.) and mallow (Malva sylvestris L.) against Haemonchus contortus in sheep. Animals 2020, 10, 219. [Google Scholar] [CrossRef]
- Váradyová, Z.; Pisarčíková, J.; Babják, M.; Hodges, A.; Mravčáková, D.; Kišidayová, S.; Königová, A.; Vadlejch, J.; Várady, M. Ovicidal and larvicidal activity of extracts from medicinal-plants against Haemonchus contortus. Exp. Parasitol. 2018, 195, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Oliveira Santos, F.; Ponce Morais Cerqueira, A.; Branco, A.; José Moreira Batatinha, M.; Borges Botura, M. Anthelmintic activity of plants against gastrointestinal nematodes of goats: A review. Parasitology 2019, 146, 1233–1246. [Google Scholar] [CrossRef] [PubMed]
- Kotze, A.C. Catalase induction protects Haemonchus contortus against hydrogen peroxide in vitro. Int. J. Parasitol. 2003, 33, 393–400. [Google Scholar] [CrossRef]
- Leal, M.L.R.; Pivoto, F.L.; Fausto, G.C.; Aires, A.R.; Grado, T.H.; Roos, D.H.; Sudati, J.H.; Wagner, C.; Costa, M.M.; Molento, M.B.; et al. Copper and selenium: Auxiliary measure to control infection by Haemonchus contortus in lambs. Exp. Parasitol. 2014, 144, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeldt, F.; Wilson, M.; Lee, G.; Kure, C.; Ou, R.; Braun, L.; de Haan, J. Oxidative stress in surgery in an ageing population: Pathophysiology and therapy. Exp. Gerontol. 2013, 48, 45–54. [Google Scholar] [CrossRef]
- Pivoto, F.L.; Torbitz, V.D.; Aires, A.R.; da Rocha, J.F.X.; Severo, M.M.; Grando, T.H.; Peiter, M.; Moresco, R.N.; da Rocha, J.B.T.; Leal, M.L.R. Oxidative stress by Haemonchus contortus in lambs: Influence of treatment with zinc edetate. Res. Vet. Sci. 2015, 102, 22–24. [Google Scholar] [CrossRef] [PubMed]
- Čobanová, K.; Váradyová, Z.; Grešáková, Ľ.; Kucková, K.; Mravčáková, D.; Várady, M. Does herbal and/or zinc dietary supplementation improve the antioxidant and mineral status of lambs with parasite infection? Antioxidants 2020, 9, 1172. [Google Scholar] [CrossRef]
- Leal, M.L.R.; de Camargo, E.V.; Ross, D.H.; Molento, M.B.; Lopes, S.T.; da Rocha, J.B.T. Effect of selenium and vitamin E on oxidative stress in lambs experimentally infected with Haemonchus contortus. Vet. Res. Commun. 2010, 34, 549–555. [Google Scholar] [CrossRef]
- Čobanová, K.; Faix, Š.; Plachá, I.; Mihaliková, K.; Váradyová, Z.; Kišidayová, S.; Grešáková, Ľ. Effects of different dietary selenium sources on antioxidant status and blood phagocytic activity in sheep. Biol. Trace Elem. Res. 2017, 175, 339–346. [Google Scholar] [CrossRef]
- Grešáková, L.; Čobanová, K.; Faix, S. Selenium retention in lambs fed diets supplemented with selenium from inorganic or organic sources. Small Rum. Res. 2013, 111, 76–82. [Google Scholar] [CrossRef]
- Hendriks, S.J.; Laven, R.A. Selenium requirements in grazing dairy cows: A review. N. Z. Vet. J. 2020, 68, 13–22. [Google Scholar] [CrossRef]
- Chauhan, S.S.; Celi, P.; Ponnampalam, E.N.; Leury, B.J.; Liu, F.; Dunshea, F.R. Antioxidant dynamics in the live animal and implications for ruminant health and product (meat/milk) quality: Role of vitamin E and selenium. Anim. Prod. Sci. 2014, 54, 1525–1536. [Google Scholar] [CrossRef]
- Mehdi, Y.; Dufrasne, I. Selenium in Cattle: A Review. Molecules 2016, 21, 545. [Google Scholar] [CrossRef]
- Kurutas, E.B. The importance of antioxidants which play the role in cellular response against oxidative/nitrosative stress: Current state. Nutr. J. 2016, 15, 71. [Google Scholar] [CrossRef]
- Arthur, J.R.; Nicol, F.; Beckett, G.J. The role of selenium in thyroid hormone metabolism and effects of selenium deficiency on thyroid hormone and iodine metabolism. Biol. Trace Elem. Res. 1992, 34, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Arsenos, G.; Fortomaris, P.; Giadinis, N.; Roubies, N.; Papadopoulos, E. Serum vitamin A and vitamin E levels of growing lambs infected or not with gastrointestinal nematodes and fed a diet containing clinoptilolite. Asian-Aust. J. Anim. Sci. 2010, 23, 567–572. [Google Scholar] [CrossRef]
- Commission Implementing Regulation (EU) 2019/49. Available online: http://data.europa.eu/eli/reg_impl/2019/49/oj (accessed on 4 January 2019).
- European Commission (EC). Council Regulation (EC) 1099/2009 of 24 September 2009 on the Protection of Animals at the Time of Killing. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex:32009R1099 (accessed on 18 November 2009).
- Horwitz, W. Official Methods of AOAC International, 17th ed.; Association of Official Analytical Chemists (AOAC) International: Gaithersburg, MD, USA; Washington, DC, USA, 2000. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber neutral detergent fiber, and non-starch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Jaradat, N.; Hussen, F.; Al Ali, A. Preliminary phytochemical screening, quantitative estimation of total flavonoids, total phenols and antioxidant activity of Ephedra alata Decne. J. Mater. Environ. Sci. 2015, 6, 1771–1778. [Google Scholar]
- Coles, G.C.; Bauer, C.; Borgsteede, F.H.M.; Geerts, S.; Klei, T.R.; Taylor, M.A.; Waller, P.J. World Association for the Advancement of Veterinary Parasitology (W.A.A.V.P) methods for the detection of anthelmintic resistance in nematodes of veterinary importance. Vet. Parasitol. 1992, 44, 35–44. [Google Scholar] [CrossRef]
- Chadio, S.E.; Kotsampasi, B.M.; Menegatos, J.G.; Zervas, G.P.; Kalogiannis, D.G. Effect of selenium supplementation on thyroid hormone levels and selenoenzyme activities in growing lambs. Biol. Trace Elem. Res. 2006, 109, 145–154. [Google Scholar] [CrossRef]
- Akasha, M.; Anderson, R.R. Thyroxine and triiodothyronine in milk of cows, goats, sheep, and guinea pigs. Proc. Soc. Exp. Biol. Med. 1984, 177, 360–371. [Google Scholar] [CrossRef]
- Kassab, A.Y.; Mohammed, A.A. Ascorbic acid administration as anti-stress before transportation of sheep. Egyptian. J. Anim. Prod. 2014, 51, 19–25. [Google Scholar]
- Liu, M.; Panda, S.K.; Luyten, W. Plant-based natural products for the discovery and development of novel anthelmintics against nematodes. Biomolecules 2020, 10, 426. [Google Scholar] [CrossRef] [PubMed]
- Mayer, M.; Vogl, C.R.; Amorena, M.; Hamburger, M.; Walkenhorst, M. Treatment of organic livestock with medicinal plants: A systematic review of European ethnoveterinary research. Forsch Komplementmed. 2014, 21, 375–386. [Google Scholar] [CrossRef] [PubMed]
- Mavrot, F.; Hertzberg, H.; Torgerson, P. Effect of gastro-intestinal nematode infection on sheep performance: A systematic review and meta-analysis. Parasit Vectors 2015, 8, 557. [Google Scholar] [CrossRef] [PubMed]
- Beigh, Y.A.; Mir, D.M.; Ganai, A.M.; Ahmad, H.A.; Muzamil, S. Body biometrics correlation studies on sheep fed wormwood (Artemisia absinthium L.) herb supplemented complete diets. Indian, J. Vet. Res. 2019, 28, 8–13. [Google Scholar] [CrossRef]
- Bahrami, A.M.; Doosti, A.; Moosavi, A.B. Effect of Matricaria chamomilla L. plant extraction on experimental infected lamb with Ostertagia ostertagi parasits. Int. J. Pharmacol. 2010, 6, 712–718. [Google Scholar] [CrossRef]
- Tufarelli, V.; Casalino, E.; D’Alessandro, A.G.; Laudadio, V. Dietary phenolic compounds: Biochemistry, metabolism and significance in animal and human health. Curr. Drug Metab. 2017, 18, 905–913. [Google Scholar] [CrossRef]
- Fausto, G.C.; Pivoto, F.L.; Costa, M.M.; dos Anjos Lopes, S.T.; França, R.T.; Molento, M.B.; Minervino, A.H.; da Rocha, J.B.; Leal, M.L. Protein profile of lambs experimentally infected with Haemonchus contortus and supplemented with selenium and copper. Parasit. Vectors 2014, 7, 355. [Google Scholar] [CrossRef][Green Version]
- Ramírez-Bribiesca, J.E.; Tórtora, J.L.; Huerta, M.; Hernández, L.M.; López, R.; Crosby, M.M. Effect of selenium-vitamin E injection in selenium-deficient dairy goats and kids on the Mexican plateau. Arq. Bras. Med. Vet. Zootec. 2005, 57, 77–84. [Google Scholar] [CrossRef]
- Cériac, S.; Jayles, C.; Arquet, R.; Feuillet, D.; Félicité, Y.; Archimède, H.; Bambou, J.C. The nutritional status affects the complete blood count of goats experimentally infected with Haemonchus contortus. BMC Vet. Res. 2017, 13, 326. [Google Scholar] [CrossRef]
- Retama-Flores, C.; Torres-Acosta, J.F.J.; Sandoval-Castro, C.A.; Aguilar-Caballero, A.J.; Cámara-Sarmiento, R.; Canul-Ku, H.L. Maize supplementation of Pelibuey sheep in a silvopastoral system: Fodder selection, nutrient intake and resilience against gastrointestinal nematodes. Animal 2012, 6, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Torres-Acosta, J.F.J.; Jacobs, D.E.; Aguilar-Caballero, A.; Sandoval-Castro, C.; Cob-Galera, L.; May-Martínez, M. Improving resilience against natural gastrointestinal nematode infections in browsing kids during the dry season in tropical Mexico. Vet. Parasitol. 2006, 135, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Alam, R.T.M.; Hassanen, E.A.A.; El-Mandrawy, S.A.M. Heamonchus contortus infection in sheep and goats: Alterations in haematological, biochemical, immunological, trace element and oxidative stress markers. J. Appl. Anim. Res. 2020, 48, 357–364. [Google Scholar] [CrossRef]
- Toker, N.Y. Blood serum vitamin A and E concentrations and distribution into lipoprotein fractions of pregnant sheep and newborn lambs. Revue Méd. Vét. 2007, 158, 413–417. [Google Scholar]
- Gardiner, M.R. Pathological changes and vitamin B12 metabolism in sheep parasitised by Haemonchus contortus, Ostertagia spp. and Trichostrongylus colubriformis. J. Helminthol. 1966, 40, 63–76. [Google Scholar] [CrossRef]
- Sykes, A.R.; Greer, A.W. Effects of parasitism on the nutrient economy of sheep. Aust. J. Exp. Agric. 2003, 43, 1393–1398. [Google Scholar] [CrossRef]
- MacGlaflin, C.E.; Zajac, A.M.; Rego, K.A.; Petersson, K.H. Effect of vitamin E supplementation on naturally acquired parasitic infection in lambs. Vet. Parasitol. 2011, 175, 300–305. [Google Scholar] [CrossRef] [PubMed]
- Combs, G.F., Jr.; Scott, M.L. Nutritional interrelationships of vitamin E and selenium. BioScience 1977, 27, 467–473. [Google Scholar] [CrossRef]
- Zoidis, E.; Seremelis, I.; Kontopoulos, N.; Danezis, G.P. Selenium-dependent antioxidant enzymes: Actions and properties of selenoproteins. Antioxidants 2018, 7, 66. [Google Scholar] [CrossRef]
- Knox, M.R.; Torres-Acosta, J.F.J.; Aguilar-Caballero, A.J. Exploiting the effect of dietary supplementation of small ruminants on resilience and resistance against gastrointestinal nematodes. Vet. Parasitol. 2006, 139, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Todini, L. Thyroid hormones in small ruminants: Effects of endogenous, environmental and nutritional factors. Animal 2007, 1, 997–1008. [Google Scholar] [CrossRef] [PubMed]
- Voudouri, A.E.; Chadio, S.E.; Menegatos, J.G.; Zervas, G.P.; Nicol, F.; Arthur, J.R. Selenoenzyme activities in selenium- and odine-deficient sheep. Biol. Trace Elem. Res. 2003, 94, 213–224. [Google Scholar] [CrossRef]
- Ventura, M.; Melo, M.; Carrilho, F. Selenium and thyroid disease: From pathophysiology to treatment. Int. J. Endocrinol. 2017, 2017, 1297658. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Dietary Substrate | DM (g/kg) | NDF (g/kg DM) | ADF (g/kg DM) | CP (g/kg DM) | N (g/kg DM) | Ash (g/kg DM) |
|---|---|---|---|---|---|---|
| Meadow hay | 917 | 675 | 445 | 74 | 12 | 68 |
| Concentrate | 899 | 263 | 145 | 261 | 42 | 124 |
| Selplex | 898 | 238 | 137 | 262 | 42 | 146 |
| Herbmix | 908 | 346 | 326 | 161 | 26 | 116 |
| Plant Species | Tannins | Saponins | Alkaloids | Terpenoids | Flavonoids | Steroids |
|---|---|---|---|---|---|---|
| Althaea officinalis | − | − | − | + | + | − |
| Petasites hybridus | − | + | − | − | − | + |
| Inula helenium | − | + | − | + | − | + |
| Plantago lanceolate | + | − | − | + | − | + |
| Rosmarinus officinalis | + | − | + | − | + | + |
| Foeniculum vulgare | − | − | − | + | − | + |
| Solidago virgaurea | + | + | − | − | − | + |
| Fumaria officinalis | + | − | − | + | + | − |
| Hyssopus officinalis | − | − | + | − | − | − |
| Herbmix | + | + | + | + | + | + |
| Day | Control | Herbmix | Selplex | SD | Significance of Effects (p-Value) | |||
|---|---|---|---|---|---|---|---|---|
| T | Time | T × Time | ||||||
| BW (kg) | 0 | 18.7 | 18.1 | 18.8 | 2.26 | 0.559 | 0.001 | 0.999 |
| 35 | 22.7 | 22.3 | 22.4 | 2.74 | ||||
| 63 | 25.5 | 24.9 | 24.7 | 2.96 | ||||
| 98 | 27.4 | 26.9 | 26.5 | 3.15 | ||||
| LWG (kg) | 35 | 4.09 | 4.21 | 3.59 | 0.792 | 0.613 | 0.001 | 0.494 |
| 63 | 2.81 | 2.60 | 2.34 | 0.742 | ||||
| 98 | 1.86 | 1.93 | 1.73 | 1.075 | ||||
| Item | Day | Control | Herbmix | Selplex | SD | Significance of Effects (p-Value) | ||
|---|---|---|---|---|---|---|---|---|
| T | Time | T × Time | ||||||
| RBC | 0 | 12.1 | 12.3 | 12.5 | 0.68 | 0.003 | 0.001 | 1.000 |
| (T/L) | 15 | 11.4 | 11.4 | 12.0 | 0.88 | |||
| 35 | 9.18 | 9.17 | 10.2 | 1.31 | ||||
| 49 | 8.57 | 8.38 | 9.53 | 1.32 | ||||
| 63 | 8.24 | 8.40 | 9.43 | 1.41 | ||||
| 77 | 7.83 | 8.08 | 8.93 | 1.41 | ||||
| 98 | 7.04 | 6.98 | 7.78 | 1.84 | ||||
| 112 | 6.91 | 7.14 | 7.62 | 1.91 | ||||
| HGB | 0 | 125 | 120 | 120 | 8.47 | 0.088 | 0.001 | 0.977 |
| (g/L) | 15 | 110 | 109 | 113 | 7.01 | |||
| 35 | 90.3 | 92.9 | 95.8 | 9.92 | ||||
| 49 | 87.9 | 88.1 | 89.5 | 11.7 | ||||
| 63 | 80.8 | 86.1 | 87.2 | 14.9 | ||||
| 77 | 79.1 | 83.5 | 85.2 | 14.3 | ||||
| 98 | 69.3 | 71.4 | 74.5 | 20.7 | ||||
| 112 | 63.6 | 70.5 | 73.6 | 22.3 | ||||
| HCT | 0 | 0.255 | 0.262 | 0.247 | 0.013 | 0.197 | 0.001 | 1.000 |
| (g/L) | 15 | 0.240 | 0.243 | 0.237 | 0.011 | |||
| 35 | 0.216 | 0.226 | 0.216 | 0.018 | ||||
| 49 | 0.213 | 0.216 | 0.206 | 0.023 | ||||
| 63 | 0.198 | 0.214 | 0.204 | 0.030 | ||||
| 77 | 0.197 | 0.210 | 0.204 | 0.027 | ||||
| 98 | 0.173 | 0.181 | 0.178 | 0.043 | ||||
| 112 | 0.162 | 0.181 | 0.178 | 0.046 | ||||
| MCV | 0 | 21.0 | 21.3 | 19.8 | 1.30 | 0.001 | 0.001 | 0.932 |
| (fL) | 15 | 23.7 | 21.5 | 19.8 | 1.32 | |||
| 35 | 25.1 | 24.9 | 21.6 | 2.43 | ||||
| 49 | 25.1 | 26.0 | 21.9 | 2.59 | ||||
| 63 | 24.1 | 25.7 | 21.8 | 2.39 | ||||
| 77 | 25.2 | 26.2 | 23.2 | 2.61 | ||||
| 98 | 24.4 | 26.3 | 23.0 | 2.83 | ||||
| 112 | 23.3 | 25.5 | 23.5 | 2.53 | ||||
| Item | Day | Control | Herbmix | Selplex | SD | Significance of Effects (p-Value) | ||
|---|---|---|---|---|---|---|---|---|
| T | Time | T × Time | ||||||
| α-Tocopherol (μg/mL) | 0 | 1.031 | 1.286 | 1.164 | 0.0041 | 0.001 | 0.001 | 0.002 |
| 35 | 1.035 | 1.388 | 1.178 | 0.6016 | ||||
| 63 | 1.079 | 1.211 | 1.221 | 0.2820 | ||||
| 98 | 1.417 | 1.508 | 1.273 | 0.4155 | ||||
| Item | Day | Control | Herbmix | Selplex | SD | Significance of Effects (p-Value) | ||
|---|---|---|---|---|---|---|---|---|
| T | Time | T × Time | ||||||
| GPx | 0 | 412.3 | 440.6 | 400.7 | 66.26 | 0.001 | 0.001 | 0.001 |
| (U/g Hb) | 35 | 420.3 | 465.9 | 742.7 | 159.8 | |||
| 63 | 349.4 | 384.0 | 813.2 | 228.3 | ||||
| 98 | 259.6 | 289.3 | 795.2 | 256.4 | ||||
| MDA | 0 | 0.256 | 0.275 | 0.273 | 0.037 | 0.132 | 0.001 | 0.666 |
| (µmol/L) | 35 | 0.306 | 0.271 | 0.301 | 0.047 | |||
| 63 | 0.230 | 0.205 | 0.239 | 0.039 | ||||
| 98 | 0.243 | 0.207 | 0.225 | 0.039 | ||||
| TAC | 0 | 0.663 | 0.678 | 0.690 | 0.030 | 0.115 | 0.001 | 0.980 |
| (mmol/L) | 35 | 0.667 | 0.659 | 0.681 | 0.036 | |||
| 63 | 0.433 | 0.429 | 0.453 | 0.040 | ||||
| 98 | 0.439 | 0.434 | 0.449 | 0.039 | ||||
| Day | Control | Herbmix | Selplex | SD | Significance of Effects (p-Value) | |||
|---|---|---|---|---|---|---|---|---|
| T | Time | T × Time | ||||||
| T3 (nmol/L) | 35 | 0.604 | 1.44 | 1.57 | 0.849 | |||
| 63 | 0.529 | 1.04 | 1.27 | 0.784 | 0.001 | 0.144 | 0.021 | |
| 98 | 0.201 | 0.074 | 1.96 | 1.12 | ||||
| T4 (nmol/L) | 35 | 62.9 | 72.1 | 79.7 | 20.2 | |||
| 63 | 60.2 | 59.0 | 69.7 | 19.0 | 0.047 | 0.020 | 0.849 | |
| 98 | 51.9 | 46.5 | 65.4 | 18.3 | ||||
| T3:T4 | 35 | 0.01 | 0.02 | 0.02 | 0.010 | |||
| 63 | 0.009 | 0.018 | 0.018 | 0.011 | 0.001 | 0.230 | 0.004 | |
| 98 | 0.004 | 0.002 | 0.03 | 0.015 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Komáromyová, M.; Mravčáková, D.; Petrič, D.; Kucková, K.; Babják, M.; Dolinská, M.U.; Königová, A.; Maďarová, M.; Pruszyńska-Oszmałek, E.; Cieslak, A.; et al. Effects of Medicinal Plants and Organic Selenium against Ovine Haemonchosis. Animals 2021, 11, 1319. https://doi.org/10.3390/ani11051319
Komáromyová M, Mravčáková D, Petrič D, Kucková K, Babják M, Dolinská MU, Königová A, Maďarová M, Pruszyńska-Oszmałek E, Cieslak A, et al. Effects of Medicinal Plants and Organic Selenium against Ovine Haemonchosis. Animals. 2021; 11(5):1319. https://doi.org/10.3390/ani11051319
Chicago/Turabian StyleKomáromyová, Michaela, Dominika Mravčáková, Daniel Petrič, Katarína Kucková, Michal Babják, Michaela Urda Dolinská, Alžbeta Königová, Michaela Maďarová, Ewa Pruszyńska-Oszmałek, Adam Cieslak, and et al. 2021. "Effects of Medicinal Plants and Organic Selenium against Ovine Haemonchosis" Animals 11, no. 5: 1319. https://doi.org/10.3390/ani11051319
APA StyleKomáromyová, M., Mravčáková, D., Petrič, D., Kucková, K., Babják, M., Dolinská, M. U., Königová, A., Maďarová, M., Pruszyńska-Oszmałek, E., Cieslak, A., Čobanová, K., Váradyová, Z., & Várady, M. (2021). Effects of Medicinal Plants and Organic Selenium against Ovine Haemonchosis. Animals, 11(5), 1319. https://doi.org/10.3390/ani11051319