
Effect of Artemisia absinthium and Malva sylvestris on Antioxidant Parameters and Abomasal Histopathology in Lambs Experimentally Infected with Haemonchus contortus

 , , ,
, , ,  , and
, and
Abstract
Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Ethics Statement
2.2. Animals and Experimental Design
2.3. Plant Methanolic Extracts
2.4. Antioxidant Parameters
2.5. Abomasum Histology
2.6. Calculations and Statistical Analysis
3. Results
3.1. Plant Methanolic Extracts
3.2. Antioxidant Parameters
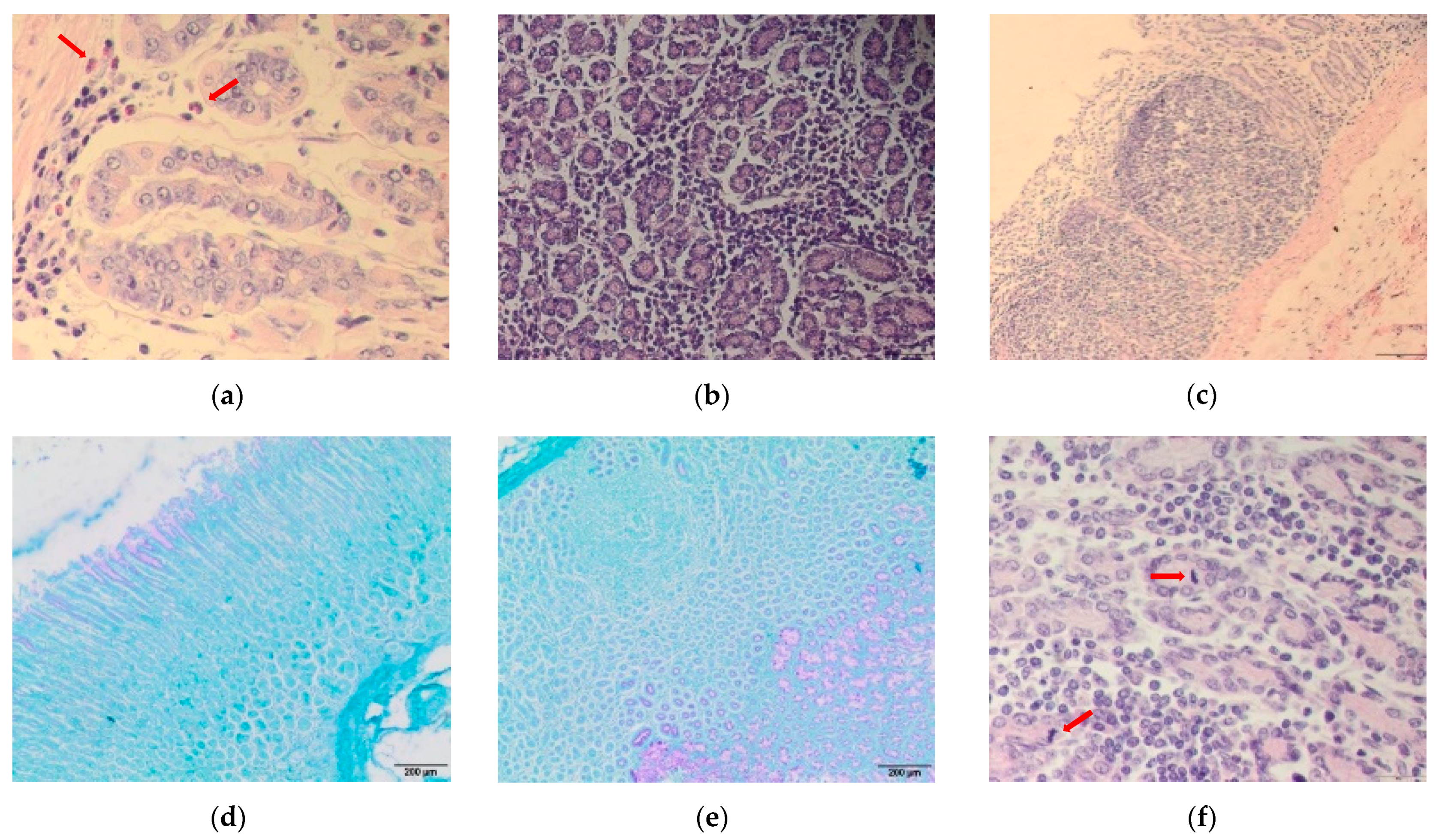
3.3. Abomasum Histology
3.4. Immune Cells in the Abomasal Mucosa
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Besier, R.B.; Kahn, L.P.; Sargison, N.D.; Van Wyk, J.A. The pathophysiology, ecology and epidemiology of Haemonchus contortus infection in small ruminants. In Haemonchus contortus and Haemonchosis—Past, Present and Future Trends, 1st ed.; Gasser, R.B., Von Sampson-Himmelstjerna, G., Eds.; Academic Press: London, UK, 2016; Volume 93, pp. 95–143. [Google Scholar] [CrossRef]
- Jackson, F.; Varady, M.; Bartley, D.J. Managing anthelmintic resistance in goats—Can we learn lessons from sheep? Small Rumin. Res. 2012, 103, 3–9. [Google Scholar] [CrossRef]
- Burke, J.M.; Miller, J.E. Sustainable approaches to parasite control in ruminant livestock. Vet. Clin. North Am. Food Anim. Pract. 2020, 36, 89–107. [Google Scholar] [CrossRef]
- Torres-Acosta, J.F.J.; Hoste, H. Alternative or improved methods to limit gastro-intestinal parasitism in grazing sheep and goats. Small Rumin. Res. 2008, 77, 159–173. [Google Scholar] [CrossRef]
- Zajíčková, M.; Nguyen, L.T.; Skálová, L.; Raisová Stuchlíková, L.; Matoušková, P. Anthelmintics in the future: Current trends in the discovery and development of new drugs against gastrointestinal nematodes. Drug Discov. Today 2020, 25, 430–437. [Google Scholar] [CrossRef]
- Githiori, J.B.; Athanasiadou, S.; Thamsborg, S.M. Use of plants in novel approaches for control of gastrointestinal helminths in livestock with emphasis on small ruminants. Vet. Parasitol. 2006, 139, 308–320. [Google Scholar] [CrossRef]
- Hoste, H.; Torres-Acosta, J.F.J.; Quijada, J.; Chan-Perez, I.; Dakheel, M.M.; Kommuru, D.S.; Mueller-Harvey, I.; Terrill, T.H. Interactions between nutrition and infections with Haemonchus contortus and related gastrointestinal nematodes in small ruminants. In Haemonchus contortus and Haemonchosis—Past, Present and Future Trends, 1st ed.; Gasser, R.B., Von Sampson-Himmelstjerna, G., Eds.; Academic Press: London, UK, 2016; Volume 93, pp. 239–351. [Google Scholar] [CrossRef]
- Spiegler, V.; Liebau, E.; Hensel, A. Medicinal plant extracts and plant-derived polyphenols with anthelmintic activity against intestinal nematodes. Nat. Prod. Rep. 2017, 34, 627–643. [Google Scholar] [CrossRef]
- Athanasiadou, S.; Kyriazakis, I. Plant secondary metabolites: Antiparasitic effects and their role in ruminant production systems. Proc. Nutr. Soc. 2004, 63, 631–639. [Google Scholar] [CrossRef]
- Acamovic, T.; Brooker, J.D. Biochemistry of plant secondary metabolites and their effects in animals. Proc. Nutr. Soc. 2005, 64, 403–412. [Google Scholar] [CrossRef]
- Wink, M. Medicinal plants: A source of anti-parasitic secondary metabolites. Molecules 2012, 17, 12771–12791. [Google Scholar] [CrossRef]
- Villalba, J.J.; Costes-Thiré, M.; Ginane, C. Phytochemicals in animal health: Diet selection and trade-offs between costs and benefits. Proc. Nutr. Soc. 2017, 76, 113–121. [Google Scholar] [CrossRef]
- Athanasiadou, S.; Kyriazakis, I.; Jackson, F.; Coop, R.L. Direct anthelmintic effects of condensed tannins towards different gastrointestinal nematodes of sheep: In vitro and in vivo studies. Vet. Parasitol. 2001, 99, 205–219. [Google Scholar] [CrossRef]
- Hoste, H.; Martinez-Ortiz-De-Montellano, C.; Manolaraki, F.; Brunet, S.; Ojeda-Robertos, N.; Fourquaux, I.; Torres-Acosta, J.F.J.; Sandoval-Castro, C.A. Direct and indirect effects of bioactive tannin-rich tropical and temperate legumes against nematode infections. Vet. Parasitol. 2012, 186, 18–27. [Google Scholar] [CrossRef]
- Liu, M.; Panda, S.K.; Luyten, W. Plant-based natural products for the discovery and development of novel anthelmintics against nematodes. Biomolecules 2020, 10, 426. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Mitre, G.F.; Olmedo-Juárez, A.; Rojo-Rubio, R.; Gonzáles-Cortázar, M.; Mendoza-de Gives, P.; Hernández-Beteta, E.E.; Reyes-Guerrero, D.E.; López-Arellano, M.E.; Vázquez-Armijo, J.F.; Ramírez-Vargas, G.; et al. Caffeoyl and coumaroyl derivates from Acacia cochliacantha exhibit ovicidal activity against Haemonchus contortus. J. Ethnopharmacol. 2017, 204, 125–131. [Google Scholar] [CrossRef]
- Delgado-Núñez, E.J.; Zamilpa, A.; Gonzáles-Cortazar, M.; Olmedo-Juárez, A.; Cardoso-Taketa, A.; Sánchez-Mendoza, E.; Tapia-Maruri, D.; Salinas-Sánchez, D.O.; Mendoza-de Gives, P. Isorhamnetin: A nematocidal flavonoid from Prosopis Leavigata leaves against Haemonchus contortus eggs and larvae. Biomolecules 2020, 10, 733. [Google Scholar] [CrossRef]
- Olmedo-Juárez, A.; Zarza-Albarran, M.A.; Rojo-Rubio, R.; Zamilpa, A.; Gonzáles-Cortazar, M.; Mondragón-Ancelmo, J.; Rivero-Péerez, N.; Mendoza-de Gives, P. Acacia farnesiana pods (plant: Fabaceae) possesses anti-parasitic compounds against Haemonchus contortus in female lambs. Exp. Parasitol. 2020, 218, 107980. [Google Scholar] [CrossRef]
- Mravčáková, D.; Komáromyová, M.; Babják, M.; Urda Dolinská, M.; Königová, A.; Petrič, D.; Čobanová, K.; Śluzarcyk, S.; Cieslak, A.; Várady, M.; et al. Anthelmintic activity of wormwood (Artemisia absinthium L.) and mallow (Malva sylvestris L.) against Haemonchus contortus in sheep. Animals 2020, 10, 219. [Google Scholar] [CrossRef] [PubMed]
- Váradyová, Z.; Kišidayová, S.; Čobanová, K.; Grešáková, L.; Babják, M.; Königová, A.; Urda Dolinská, M.; Várady, M. The impact of a mixture of medicinal herbs on ruminal fermentation, parasitological status and hematological parameters of the lambs experimentally infected with Haemonchus contortus. Small Rumin. Res. 2017, 151, 124–132. [Google Scholar] [CrossRef]
- Váradyová, Z.; Mravčáková, D.; Babják, M.; Bryszak, M.; Grešáková, L.; Čobanová, K.; Kišidayová, S.; Plachá, I.; Königová, A.; Cieslak, A.; et al. Effects of herbal nutraceuticals and/or zinc against Haemonchus contortus in lambs experimentally infected. BMC Vet. Res. 2018, 14, 78. [Google Scholar] [CrossRef]
- Mravčáková, D.; Váradyová, Z.; Kopčáková, A.; Čobanová, K.; Grešáková, L.; Kišidayová, S.; Babják, M.; Urda Dolinská, M.; Dvorožňáková, E.; Königová, A.; et al. Natural chemotherapeutic alternatives for controlling of haemonchosis in sheep. BMC Vet. Res. 2019, 15, 302. [Google Scholar] [CrossRef]
- Coop, R.L.; Kyriazakis, I. Influence of host nutrition on the development and consequences of nematode parasitism in ruminants. Trends Parasitol. 2001, 17, 325–330. [Google Scholar] [CrossRef]
- Bambou, J.C.; Archimède, H.; Arquet, R.; Mahieu, M.; Alexandre, G.; González-Garcia, E.; Mandonnet, N. Effect of dietary supplementation on resistance to experimental infection with Haemonchus contortus in Creole kids. Vet. Parasitol. 2011, 178, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Costes-Thiré, M.; Laurent, P.; Ginane, C.; Villalba, J.J. Diet selection and trade-offs between condensed tannins and nutrients in parasitized sheep. Vet. Parasitol. 2019, 271, 14–21. [Google Scholar] [CrossRef]
- Balic, A.; Bowles, V.M.; Meeusen, E.N.T. Mechanisms of immunity to Haemonchus contortus infection in sheep. Parasite Immunol. 2002, 24, 39–46. [Google Scholar] [CrossRef]
- Alba-Hurtado, F.; Muñoz-Guzmán, M.A. Immune responses associated with resistance to haemonchosis in sheep. Biomed. Res. Int. 2013, 2013, 162158. [Google Scholar] [CrossRef]
- McRae, K.M.; Stear, M.J.; Good, B.; Keane, O.M. The host immune response to gastrointestinal nematode infection in sheep. Parasite Immunol. 2015, 37, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Machado, V.; Da Silva, A.S.; Schafer, A.S.; Aires, A.R.; Tonin, A.A.; Oliveira, C.B.; Hermes, C.L.; Almeida, T.C.; Moresco, R.F.; Stefani, L.M.; et al. Relationship between oxidative stress and pathological findings in abomasum of infected lambs by Haemonchus contortus. Pathol. Res. Pract. 2014, 210, 812–817. [Google Scholar] [CrossRef]
- Gessner, D.K.; Ringseis, R.; Eder, K. Potential of plant polyphenols to combat oxidative stress and inflammatory processes in farm animals. J. Anim. Physiol. Anim. Nutr. 2017, 101, 605–628. [Google Scholar] [CrossRef]
- Poli, C.H.E.C.; Thornton-Kurth, K.J.; Legako, J.F.; Bremm, C.; Hampel, V.S.; Hall, J.; Ipharraguerre, I.R.; Villalba, J.J. Self-selection of plant bioactive compounds by sheep in response to challenge infection with Haemonchus contortus. Physiol. Behav. 2018, 194, 302–310. [Google Scholar] [CrossRef]
- Balic, A.; Bowles, V.M.; Meeusen, E.N.T. Cellular profiles in the abomasal mucosa and lymph node during primary infection with Haemonchus contortus in sheep. Vet. Immunol. Immunopathol. 2000, 75, 109–120. [Google Scholar] [CrossRef]
- Amarante, A.F.T.; Bricarello, P.A.; Huntley, J.F.; Mazzolin, L.P.; Gomes, J.C. Relationship of abomasal histology and parasite-specific immunoglobulin A with the resistance to Haemonchus contortus infection in three breeds of sheep. Vet. Parasitol. 2005, 128, 99–107. [Google Scholar] [CrossRef]
- Muñoz-Guzmán, M.A.; Cuenca-Verde, C.; Valdivia-Anda, G.; Cuéllar-Ordaz, J.A.; Alba-Hurtado, F. Differential immune response between fundic and pyloric abomasal regions upon experimental ovine infection with Haemonchus contortus. Vet. Parasitol. 2012, 185, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Čobanová, K.; Chrastinová Chrenková, M.; Polačiková, M.; Formelová, Z.; Ivanišinová, O.; Ryzner, M.; Grešáková, L. The effect of different dietary zinc sources on mineral deposition and antioxidant indices in rabbit tissues. World Rabbit Sci. 2018, 26, 241–248. [Google Scholar] [CrossRef]
- Paglia, D.E.; Valentine, W.N. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med. 1967, 70, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Benzie, I.F.F.; Strain, J.J. Ferric reducing/antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for stimultaneous measurement of total antioxidant power and ascorbic acid concentration. Methods Enzymol. 1999, 299, 15–27. [Google Scholar] [CrossRef]
- Jo, C.; Ahn, D.U. Fluorometric analysis of 2-thiobarbituric acid reactive substances in Turkey. Poult. Sci. 1998, 77, 475–480. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Barros, L.; Carvalho, A.M.; Ferreira, I.C.F.R. Leaves, flowers, immature fruits and leafy flowered stems of Malva sylvestris: A comparative study of the nutraceutical potential and composition. Food Chem. Toxicol. 2010, 48, 1466–1472. [Google Scholar] [CrossRef]
- El-Saber Batiha, G.; Olatunde, A.; El-Mleeh, A.; Hetta, H.F.; Al-Rejaie, S.; Alghamdi, S.; Zahoor, M.; Beshbishy, A.M.; Murata, T.; Zaragoza-Bastida, A.; et al. Bioactive compounds, pharmacological actions, and pharmacokinetics of wormwood (Artemisia absinthium). Antibiotics 2020, 9, 353. [Google Scholar] [CrossRef]
- Do Reo Leal, M.L.; De Camargo, E.V.; Henrique Ross, D.; Molento, M.B.; Dos Anjos Lopes, S.T.; Da Rocha, J.B.T. Effect of selenium and vitamin E on oxidative stress in lambs experimentally infected with Haemonchus contortus. Vet. Res. Commun. 2010, 34, 549–555. [Google Scholar] [CrossRef]
- Kaurinovic, B.; Vastag, D. Flavonoids and phenolic acids as potential natural antioxidants. In Antioxidants, 1st ed.; Shalaby, E., Ed.; IntechOpen: London, UK, 2019; pp. 127–146. [Google Scholar] [CrossRef]
- Celi, P. The role of oxidative stress in small ruminants’ health and production. Rev. Bras. Zootec. 2010, 39, 348–363. [Google Scholar] [CrossRef]
- Bora, K.S.; Sharma, A. Evaluation of antioxidant and free-radical scavenging potential of Artemisia absinthium. Pharm. Biol. 2011, 49, 1216–1223. [Google Scholar] [CrossRef]
- Paolini, V.; Bergeaud, J.P.; Grisez, C.; Prevot, F.; Dorchies, P.; Hoste, H. Effects of condensed tannins on goats experimentally infected with Haemonchus contortus. Vet. Parasitol. 2003, 113, 253–261. [Google Scholar] [CrossRef]
- Pérez, J.; Garcia, P.M.; Hernandez, S.; Martinez-Moreno, A.; Martin De Las Mulas, J.; Camara, S. Pathological and immunohistochemical study of the abomasum and abomasal lymph nodes in goats experimentally infected with Haemonchus contortus. Vet. Res. 2001, 32, 463–473. [Google Scholar] [CrossRef]
- Venturina, V.M.; Gossner, A.G.; Hopkins, J. The immunology and genetics of resistance of sheep to Teladorsagia circumcincta. Vet. Res. Commun. 2013, 37, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Salman, S.K.; Duncan, J.L. The abomasal histology of worm-free sheep given primary and challenge infections of Haemonchus contortus. Vet. Parasitol. 1984, 16, 43–54. [Google Scholar] [CrossRef]
- Grencis, R.K.; Humphreys, N.E.; Bancroft, A.J. Immunity to gastrointestinal nematodes: Mechanisms and myths. Immunol. Rev. 2014, 260, 183–205. [Google Scholar] [CrossRef]
- Balic, A.; Cunningham, C.P.; Meeusen, E.N.T. Eosinophil interactions with Haemonchus contortus larvae in the ovine gastrointestinal tract. Parasite Immunol. 2006, 28, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Balic, A.; Bowles, V.M.; Meeusen, E.N.T.; Bowles, M. The immunobiology of gastrointestinal nematode infections in ruminants. Adv. Parasitol. 2000, 45, 181–241. [Google Scholar] [CrossRef] [PubMed]
- Terefe, G.; Lacroux, C.; Andreoletti, O.; Grisez, C.; Prevot, F.; Bergeaud, J.P.; Penicaud, J.; Rouillon, V.; Gruner, L.; Brunel, J.C.; et al. Immune response to Haemonchus contortus infection in susceptible (INRA 401) and resistant (Barbados Black Belly) breeds of lambs. Parasite Immunol. 2007, 29, 415–424. [Google Scholar] [CrossRef]
- Krystel-Whittemore, M.; Dileepan, K.N.; Wood, J.G. Mast cell: A multi-functional master cell. Front. Immunol. 2016, 6, 620. [Google Scholar] [CrossRef]
- Galli, S.J.; Nakae, S.; Tsai, M. Mast cells in the development of adaptive immune responses. Nat. Immunol. 2005, 6, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Tzamaloukas, O.; Athanasiadou, S.; Kyriazakis, I.; Huntley, J.F.; Jackson, F. The effect of chicory (Cichorium intybus) and sulla (Hedysarum coronarium) on larval development and mucosal cell responses of growing lambs challenged with Teladorsagia circumcincta. Parasitology 2006, 132, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Robinson, N.; Piedrafita, D.; Snibson, K.; Harrison, P.; Meeusen, E.N.T. Immune cell kinetics in the ovine abomasal mucosa following hyperimmunization and challenge with Haemonchus contortus. Vet. Res. 2010, 41, 37. [Google Scholar] [CrossRef]
- Penissi, A.B.; Piezzi, R.S. Effect of dehydroleucodine on mucus production: A quantitative study. Dig. Dis. Sci. 1999, 44, 708–712. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Fraction (F) | RT (min) | Composition of Fraction | FW (mg/g) | Concentration (µg/mL) |
|---|---|---|---|---|
| A. absinthium | ||||
| F1 | 1.00–5.25 | Very polar substances, sugars, polar peptides and proteins, polar phenolic acids | 45.913 | 46,000 |
| F2 | 5.25–18.00 | Polyphenols, less polar phenolic acids, sesquiterpenes | 19.813 | 20,000 |
| F3 | 18.00–25.00 | A similar spectrum of substances as F2, but of lower polarity | 5.583 | 6000 |
| F4 | 25.00–45.00 | Nonpolar substances, lipids, resins, di- and triterpenes | 12.102 | 12,000 |
| M. sylvestris | ||||
| F1 | 1.00–5.25 | Sugars, polar phenols | 150.441 | 150,000 |
| F2 | 5.25–10.50 | Anthocyanins, other phenolic compounds | 41.818 | 42,000 |
| F3 | 10.50–17.60 | Polyphenols, flavonoids, phenolic acids | 9.403 | 9000 |
| F4 | 17.60–45.00 | Nonpolar substances, esters, fatty acids, lipids | 18.368 | 18,000 |
| Item | Treatment | |||||
|---|---|---|---|---|---|---|
| UNS | ART | MAL | ARTMAL | SEM | p-Value | |
| Abomasum | ||||||
| MDA (nmol/g protein) | 277.71 | 237.69 | 209.35 | 174.78 * | 13.67 | 0.048 |
| GPx (U/g protein) | 98.46 | 88.48 | 92.63 | 95.67 | 2.314 | 0.508 |
| TAC (µmol/g protein) | 50.98 | 51.69 | 62.94 * | 55.65 | 2.381 | 0.050 |
| Serum | ||||||
| MDA (μmol/L) | 0.188 | 0.192 | 0.185 | 0.160 | 0.008 | 0.498 |
| GPx (U/mL) | 0.106 | 0.095 | 0.102 | 0.102 | 0.007 | 0.957 |
| TAC (mmol/L) | 0.593 | 0.512 | 0.578 | 0.542 | 0.013 | 0.102 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mravčáková, D.; Sobczak-Filipiak, M.; Váradyová, Z.; Kucková, K.; Čobanová, K.; Maršík, P.; Tauchen, J.; Vadlejch, J.; Mickiewicz, M.; Kaba, J.; et al. Effect of Artemisia absinthium and Malva sylvestris on Antioxidant Parameters and Abomasal Histopathology in Lambs Experimentally Infected with Haemonchus contortus. Animals 2021, 11, 462. https://doi.org/10.3390/ani11020462
Mravčáková D, Sobczak-Filipiak M, Váradyová Z, Kucková K, Čobanová K, Maršík P, Tauchen J, Vadlejch J, Mickiewicz M, Kaba J, et al. Effect of Artemisia absinthium and Malva sylvestris on Antioxidant Parameters and Abomasal Histopathology in Lambs Experimentally Infected with Haemonchus contortus. Animals. 2021; 11(2):462. https://doi.org/10.3390/ani11020462
Chicago/Turabian StyleMravčáková, Dominika, Małgorzata Sobczak-Filipiak, Zora Váradyová, Katarína Kucková, Klaudia Čobanová, Peter Maršík, Jan Tauchen, Jaroslav Vadlejch, Marcin Mickiewicz, Jaroslaw Kaba, and et al. 2021. "Effect of Artemisia absinthium and Malva sylvestris on Antioxidant Parameters and Abomasal Histopathology in Lambs Experimentally Infected with Haemonchus contortus" Animals 11, no. 2: 462. https://doi.org/10.3390/ani11020462
APA StyleMravčáková, D., Sobczak-Filipiak, M., Váradyová, Z., Kucková, K., Čobanová, K., Maršík, P., Tauchen, J., Vadlejch, J., Mickiewicz, M., Kaba, J., & Várady, M. (2021). Effect of Artemisia absinthium and Malva sylvestris on Antioxidant Parameters and Abomasal Histopathology in Lambs Experimentally Infected with Haemonchus contortus. Animals, 11(2), 462. https://doi.org/10.3390/ani11020462