
Biochemistry and Immune Biomarkers Indicate Interacting Effects of Pre- and Postnatal Stressors in Pigs across Sexes

 ,
,  ,
,  , ,
, ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Experiments
2.2. Biochemistry Profiling
2.3. Immunological Profiling
2.4. Statistical Analysis
3. Results
3.1. Effects of Pre- and Postnatal Stressors and Sex on Serum Chemistry Analytes
3.2. Effects of Pre- and Postnatal Stressors and Sex on Serum Cytokines
4. Discussion
4.1. Effects of Pre- and Post-Natal Stressors and Sex on Serum Metabolic Parameters
4.2. Effects of Pre- and Postnatal Stressors and Sex on Serum Immune Parameters
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fair, T. The contribution of the maternal immune system to the establishment of pregnancy in cattle. Front. Immunol. 2015, 6, 7. [Google Scholar] [CrossRef] [PubMed]
- Boulanger-Bertolus, J.; Pancaro, C.; Mashour, G.A. Increasing role of maternal immune activation in neurodevelopmental disorders. Front Behav. Neurosci. 2018, 12. [Google Scholar] [CrossRef]
- Antonson, A.M.; Radlowski, E.C.; Lawson, M.A.; Rytych, J.L.; Johnson, R.W. Maternal viral infection during pregnancy elicits anti-social behavior in neonatal piglet offspring independent of postnatal microglial cell activation. Brain Behav. Immun. 2017, 59, 300–312. [Google Scholar] [CrossRef]
- Yee, N.; Ribic, A.; de Roo, C.C.; Fuchs, E. Differential effects of maternal immune activation and juvenile stress on anxiety-like behaviour and physiology in adult rats: No evidence for the “double-hit hypothesis”. Behav. Brain Res. 2011, 224, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Markham, J.A.; Koenig, J.I. Prenatal stress: Role in psychotic and depressive diseases. Psychopharmacology 2011, 214, 89–106. [Google Scholar] [CrossRef]
- Garay, P.A.; Hsiao, E.Y.; Patterson, P.H.; McAllister, A.K. Maternal immune activation causes age- and region-specific changes in brain cytokines in offspring throughout development. BrainBehav. Immun. 2013, 31, 54–68. [Google Scholar] [CrossRef] [PubMed]
- Burdick Sanchez, N.C.; Carroll, J.A.; Arthingon, J.D.; Lancaster, P.A. Exposure to lipopolysaccharide in utero alters the postnatal metabolic response in heifers1,2,3. J. Anim. Sci. 2017, 95, 5176–5183. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pileri, E.; Mateu, E. Review on the transmission porcine reproductive and respiratory syndrome virus between pigs and farms and impact on vaccination. Vet. Res. 2016, 47, 108. [Google Scholar] [CrossRef] [PubMed]
- Nathues, H.; Alarcon, P.; Rushton, J.; Jolie, R.; Fiebig, K.; Jimenez, M.; Geurts, V.; Nathues, C. Cost of porcine reproductive and respiratory syndrome virus at individual farm level—An economic disease model. Prev. Vet. Med. 2017, 142, 16–29. [Google Scholar] [CrossRef]
- Leibler, J.H.; Otte, J.; Roland-Holst, D.; Pfeiffer, D.U.; Soares Magalhaes, R.; Rushton, J.; Graham, J.P.; Silbergeld, E.K. Industrial food animal production and global health risks: Exploring the ecosystems and economics of avian influenza. EcoHealth 2009, 6, 58–70. [Google Scholar] [CrossRef]
- Adda, J. Economic activity and the spread of viral diseases: Evidence from high frequency data. Inst. Study Labor 2015, 131, 891–941. [Google Scholar] [CrossRef]
- Walker, A.K.; Nakamura, T.; Byrne, R.J.; Naicker, S.; Tynan, R.J.; Hunter, M.; Hodgson, D.M. Neonatal lipopolysaccharide and adult stress exposure predisposes rats to anxiety-like behaviour and blunted corticosterone responses: Implications for the double-hit hypothesis. Psychoneuroendocrinology 2009, 34, 1515–1525. [Google Scholar] [CrossRef]
- Maynard, T.M.; Sikich, L.; Lieberman, J.A.; LaMantia, A.-S. Neural development, cell-cell signaling, and the “two-hit” hypothesis of schizophrenia. Schizophr. Bull. 2001, 27, 457–476. [Google Scholar] [CrossRef] [PubMed]
- Imanaka, A.; Morinobu, S.; Toki, S.; Yamawaki, S. Importance of early environment in the development of post-traumatic stress disorder-like behaviors. Behav. Brain Res. 2006, 173, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Giovanoli, S.; Engler, H.; Engler, A.; Richetto, J.; Voget, M.; Willi, R.; Winter, C.; Riva, M.A.; Mortensen, P.B.; Feldon, J.; et al. Stress in puberty unmasks latent neuropathological consequences of prenatal immune activation in mice. Science 2013, 339, 1095–1099. [Google Scholar] [CrossRef] [PubMed]
- Bayer, T.A.; Falkai, P.; Maier, W. Genetic and non-genetic vulnerability factors in schizophrenia: The basis of the “two hit hypothesis”. J. Psychiatr. Res. 1999, 33, 543–548. [Google Scholar] [CrossRef]
- Rymut, H.E.; Bolt, C.R.; Caputo, M.P.; Houser, A.K.; Antonson, A.M.; Zimmerman, J.D.; Villamil, M.B.; Southey, B.R.; Rund, L.A.; Johnson, R.W.; et al. Long-lasting impact of maternal immune activation and interaction with a second immune challenge on pig behavior. Front. Vet. Sci. 2020, 9. [Google Scholar] [CrossRef]
- Cameron, J.L. Metabolic cues for the onset of puberty. Horm. Res. Paediatr. 1991, 36, 97–103. [Google Scholar] [CrossRef]
- Castillo, C.; Abuelo, A.; Hernández, J. Usefulness of metabolic profiling in the assessment of the flock’s health status and productive performance. Small Rumin. Res. 2016, 142, 28–30. [Google Scholar] [CrossRef]
- Keever, M.R.; Zhang, P.; Bolt, C.R.; Antonson, A.M.; Rymut, H.E.; Caputo, M.P.; Houser, A.K.; Hernandez, A.G.; Southey, B.R.; Rund, L.A.; et al. Lasting and Sex-Dependent Impact of Maternal Immune Activation on Molecular Pathways of the Amygdala. Front. Neurosci. 2020, 14. [Google Scholar] [CrossRef]
- Vanderhaeghe, C.; Dewulf, J.; Jourquin, J.; De Kruif, A.; Maes, D. Incidence and prevention of early parturition in sows. Reprod. Domest. Anim. Zuchthyg. 2011, 46, 428–433. [Google Scholar] [CrossRef] [PubMed]
- Tospitakkul, P.; Kraomkaew, K.; Thammasin, K.; Uttarak, P.; Nuntapaitoon, M.; De Rensis, F.; Tummaruk, P. Induction of parturition by double administration of prostaglandin F2α in sows reduces the variation of gestation length without affecting the colostrum yield and piglet performance. J. Vet. Med. Sci. 2019, 81, 1334–1340. [Google Scholar] [CrossRef] [PubMed]
- Daş, G.; Vernunft, A.; Görs, S.; Kanitz, E.; Weitzel, J.M.; Brüssow, K.P.; Metges, C.C. Effects of general anesthesia with ketamine in combination with the neuroleptic sedatives xylazine or azaperone on plasma metabolites and hormones in pigs12. J. Anim. Sci. 2016, 94, 3229–3239. [Google Scholar] [CrossRef] [PubMed]
- Meyer, U.; Feldon, J. To poly(I:C) or not to poly(I:C): Advancing preclinical schizophrenia research through the use of prenatal immune activation models. Neuropharmacology 2012, 62, 1308–1321. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Huang, J.; Yang, S.; Zhao, Y.; Xiang, A.; Cao, J.; Fan, B.; Wu, Z.; Zhao, J.; Zhao, S.; et al. Whole blood transcriptome comparison of pigs with extreme production of in vivo dsRNA-induced serum IFN-a. Dev. Comp. Immunol. 2014, 44, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Gainer, J.H.; Guarnieri, J. Effects of poly I:C in porcine iron deficient neutropenia. Cornell Vet. 1985, 75, 454–465. [Google Scholar]
- Kaneko, J.; Harvey, J.; Bruss, M. Clinical Biochemistry of Domestic Animals; Academic Press: San Diego, CA, USA, 1997; Volume 22, ISBN 9780123704917. [Google Scholar]
- Vap, L.M.; Weiser, M.G. Field chemistry analysis. Vet. Clin. North Am. Food Anim. Pract. 2007, 23, 427–442. [Google Scholar] [CrossRef]
- Radostits, O.; Gay, C.; Hinchcliff, K.; Constable, P. Veterinary Medicine, A Textbook of the Diseases of Cattle, Horses, Sheep, Pigs and Goats, 10th ed.; Elsevier Saunders: Philadelphia, PA, USA, 2006; p. 2065. [Google Scholar]
- Harris, G.; Chen, W. Profiling of Cytokine and Chemokine Responses Using Multiplex Bead Array Technology. In Immunoproteomics: Methods and Protocols; Fulton, K.M., Twine, S.M., Eds.; Springer: New York, NY, USA, 2019; pp. 79–94. [Google Scholar]
- Staples, E.; Ingram, R.J.M.; Atherton, J.C.; Robinson, K. Optimising the quantification of cytokines present at low concentrations in small human mucosal tissue samples using Luminex assays. J. Immunol. Methods 2013, 394, 1–9. [Google Scholar] [CrossRef]
- Williams, A.; Greene, N.; Kimbro, K. Increased circulating cytokine levels in African American women with obesity and elevated HbA1c. Cytokine 2020, 128, 154989. [Google Scholar] [CrossRef]
- Yu, K.; Canalias, F.; Solà-Oriol, D.; Arroyo, L.; Pato, R.; Saco, Y.; Terré, M.; Bassols, A. Age-Related Serum Biochemical Reference Intervals Established for Unweaned Calves and Piglets in the Post-weaning Period. Front. Vet. Sci. 2019, 6. [Google Scholar] [CrossRef] [PubMed]
- Kälsch, J.; Bechmann, L.P.; Heider, D.; Best, J.; Manka, P.; Kälsch, H.; Sowa, J.-P.; Moebus, S.; Slomiany, U.; Jöckel, K.-H.; et al. Normal liver enzymes are correlated with severity of metabolic syndrome in a large population based cohort. Sci. Rep. 2015, 5, 13058. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, K.; Misra, P.; Singh, A.; Mukherjee, B.; Ambade, V.N. Study of blood urea nitrogen (bun), serum creatinine in diabetic and non-diabetic patients in a tertiary care hospital. Int. J. Med Biomed. Stud. 2019, 3. [Google Scholar] [CrossRef]
- Drochner, W.; Schollenberger, M.; Götz, S.; Lauber, U.; Tafaj, M.; Piepho, H.-P. Subacute effects of moderate feed loads of isolated Fusarium toxin deoxynivalenol on selected parameters of metabolism in weaned growing piglets. J. Anim. Physiol. Anim. Nutr. 2006, 90, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Nozad, S.; Ramin, A.-G.; Moghadam, G.; Asri-Rezaei, S.; Babapour, A.; Ramin, S. Relationship between blood urea, protein, creatinine, triglycerides and macro-mineral concentrations with the quality and quantity of milk in dairy Holstein cows. Vet. Res. Forum 2012, 3, 55–59. [Google Scholar]
- Ramin, A.G.; Aghazadeh, A.; Karamian, T.; Ramin, S. Correlations of Dietary Crude Protein and Gross Energy on Blood Glucose and Urea, Milk Urea and Lactose Concentrations in Lactating Ewes. Acta Vet. 2010, 79, 369–375. [Google Scholar] [CrossRef][Green Version]
- Vuillermot, S.; Luan, W.; Meyer, U.; Eyles, D. Vitamin D treatment during pregnancy prevents autism-related phenotypes in a mouse model of maternal immune activation. Mol. Autism 2017, 8, 9. [Google Scholar] [CrossRef]
- Xiao, Y.-P.; Wu, T.-X.; Hong, Q.-H.; Sun, J.-M.; Chen, A.-G.; Yang, C.-M.; Li, X.-Y. Response to weaning and dietary L-glutamine supplementation: Metabolomic analysis in piglets by gas chromatography/mass spectrometry. J. Zhejiang Univ. Sci. B 2012, 13, 567–578. [Google Scholar] [CrossRef]
- Liao, B.; Zhang, K.; Ding, X.; Xu, Z.; Bai, S. Effects of dietary supplementation with 25-OH-D3 on antibody and cytokine levels in serum and intestinal contents of weanling pigs with or without rotavirus challenge. Chin. J. Anim. Nutr. 2011, 23, 34–42. [Google Scholar]
- Kang, K.P.; Lee, J.E.; Lee, A.S.; Jung, Y.J.; Kim, D.; Lee, S.; Hwang, H.P.; Kim, W.; Park, S.K. Effect of gender differences on the regulation of renal ischemia-reperfusion-induced inflammation in mice. Mol. Med. Rep. 2014, 9, 2061–2068. [Google Scholar] [CrossRef]
- Yeom, S.-C.; Cho, S.-Y.; Park, C.-G.; Lee, W.-J. Analysis of reference interval and age-related changes in serum biochemistry and hematology in the specific pathogen free miniature pig. Lab. Anim. Res. 2012, 28, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Wilson, G.D.A.; Harvey, D.G.; Snook, C.R. A Review of Factors Affecting Blood Biochemistry in the Pig. Br. Vet. J. 1972, 128, 596–610. [Google Scholar] [CrossRef]
- Pang, V.F.; Swanson, S.P.; Beasley, V.R.; Buck, W.B.; Haschek, W.M. The toxicity of T-2 toxin in swine following topical application: I. Clinical signs, pathology, and residue concentrations. Fundam. Appl. Toxicol. 1987, 9, 41–49. [Google Scholar] [CrossRef]
- Kornegay, E.T.; Miller, E.R.; Brent, B.E.; Long, C.H.; Ullrey, D.E.; Hoefer, J.A. Effect of fasting and refeeding on body weight, rectal temperature. J. Nutr. 1964, 84, 295–304. [Google Scholar] [CrossRef]
- Calloway, D.H.; Hilf, R.; Munson, A.H. Effects of chronic food restriction in swine. J. Nutr. 1962, 76, 365–374. [Google Scholar] [CrossRef]
- Gupta, A.; Houston, B. A comprehensive review of the bioenergetics of fatty acid and glucose metabolism in the healthy and failing heart in nondiabetic condition. Heart Fail. Rev. 2017, 22, 825–842. [Google Scholar] [CrossRef]
- Hyder, M.A.; Hasan, M.; Mohieldein, A.H. Comparative Levels of ALT, AST, ALP and GGT in Liver associated Diseases. Semant. Sch. 2013, 3, 280–284. [Google Scholar]
- Koenig, G.; Seneff, S. Gamma-Glutamyltransferase: A Predictive Biomarker of Cellular Antioxidant Inadequacy and Disease Risk. Dis. Markers 2015, 2015, 818570. [Google Scholar] [CrossRef]
- Elefsiniotisa, I.S.; Brokalakia, H.; Argyropoulosa, E.; Magaziotou, I.; Derdemezib, A.; Mihasa, C.; Tsoumakasb, K. Evaluation of liver enzymes in asymptomatic chronic hepatitis B virus infected pregnant women. Ann. Gastroenterol. 2013, 26, 59–65. [Google Scholar]
- Luo, Z.; Zhu, W.; Guo, Q.; Luo, W.; Zhang, J.; Xu, W.; Xu, J. Weaning Induced Hepatic Oxidative Stress, Apoptosis, and Aminotransferases through MAPK Signaling Pathways in Piglets. Oxidative Med. Cell. Longev. 2016, 2016, 4768541. [Google Scholar] [CrossRef]
- Nakamura, H.; Kouda, K.; Wenying, F.; Watanabe, T.; Takeuchi, H. Suppressive Effects on Allergic Contact Dermatitis by Short-Term Fasting. Toxicol. Pathol. 2001, 29, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Felsenfeld, A.J.; Jara, A.; Avedian, G.; Kleeman, C.R. Effects of fasting, feeding, and bisphosphonate administration on serum calcitriol levels in phosphate-deprived rats. Kidney Int. 2000, 58, 1016–1022. [Google Scholar] [CrossRef] [PubMed]
- Moe, S.M. Disorders involving calcium, phosphorus, and magnesium. Prim. Care 2008, 35, 215. [Google Scholar] [CrossRef] [PubMed]
- Oster, M.; Just, F.; Büsing, K.; Wolf, P.; Polley, C.; Vollmar, B.; Muráni, E.; Ponsuksili, S.; Wimmers, K. Toward improved phosphorus efficiency in monogastrics-interplay of serum, minerals, bone, and immune system after divergent dietary phosphorus supply in swine. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 310, R917–R925. [Google Scholar] [CrossRef]
- Pitorri, F.; Carmichael, N.; Massei, G.; Coats, J.; Cowan, D.; Brash, M.; Williams, T. Comparison of serum biochemistry and haematology values in captive and free-ranging wild boar (Sus scrofa). Comp. Clin. Pathol. 2015, 24, 281–286. [Google Scholar] [CrossRef]
- Tau, G.; Rothman, P. Biologic functions of the IFN-gamma receptors. Allergy 1999, 54, 1233–1251. [Google Scholar] [CrossRef]
- Onore, C.E.; Schwartzer, J.J.; Careaga, M.; Berman, R.F.; Ashwood, P. Maternal immune activation leads to activated inflammatory macrophages in offspring. Brain Behav. Immun. 2014, 38, 220–226. [Google Scholar] [CrossRef]
- Garcia-Valtanen, P.; van Diermen, B.A.; Lakhan, N.; Lousberg, E.L.; Robertson, S.A.; Hayball, J.D.; Diener, K.R. Maternal host responses to poly(I:C) during pregnancy leads to both dysfunctional immune profiles and altered behaviour in the offspring. Am. J. Reprod. Immunol. 2020, e13260. [Google Scholar] [CrossRef]
- Malagrino, P.A.; Venturini, G.; Yogi, P.S.; Dariolli, R.; Padilha, K.; Kiers, B.; Gois, T.C.; Cardozo, K.H.M.; Carvalho, V.M.; Salgueiro, J.S.; et al. Proteome analysis of acute kidney injury—Discovery of new predominantly renal candidates for biomarker of kidney disease. J. Proteom. 2017, 151, 66–73. [Google Scholar] [CrossRef]
- Solek, C.M.; Farooqi, N.; Verly, M.; Lim, T.K.; Ruthazer, E.S. Maternal immune activation in neurodevelopmental disorders. Dev. Dyn. 2018, 247, 588–619. [Google Scholar] [CrossRef]
- Woodward, E.A.; Prêle, C.M.; Nicholson, S.E.; Kolesnik, T.B.; Hart, P.H. The anti-inflammatory effects of interleukin-4 are not mediated by suppressor of cytokine signalling-1 (SOCS1). Immunology 2010, 131, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, P.; Contasta, I.; Del Beato, T.; Ciccone, F.; Berghella, A.M. Gender-specific cytokine pathways, targets, and biomarkers for the switch from health to adenoma and colorectal cancer. Clin. Dev. Immunol. 2011, 2011, 819724. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Analyte 1 | Prenatal Stress | Sex | Post-Natal Stress | Prenatal by Sex | Pre- by Postnatal | Postnatal by Sex | Pre- by Postnatal by Sex |
|---|---|---|---|---|---|---|---|
| AG Ratio | 0.080 | 0.114 | 0.137 | 0.445 | 0.819 | 0.467 | 0.826 |
| Albumin | 0.479 | 0.037 | 0.372 | 0.778 | 0.450 | 0.904 | 0.416 |
| ALP | 0.342 | 0.086 | 0.570 | 0.096 | 0.457 | 0.735 | 0.100 |
| Anion Gap | 0.626 | 0.622 | 0.179 | 0.810 | 0.099 | 0.304 | 0.153 |
| AST | 0.234 | 0.917 | 0.028 | 0.656 | 0.322 | 0.090 | 0.298 |
| Bicarbonate | 0.288 | 0.261 | 0.010 | 0.926 | 0.297 | 0.718 | 0.659 |
| Bilirubin | 0.698 | 0.078 | 0.001 | 0.922 | 0.676 | 0.217 | 0.058 |
| BUN | 0.843 | 0.521 | 0.001 | 0.695 | 0.537 | 0.658 | 0.002 |
| Calcium | 0.282 | 0.153 | 0.005 | 0.985 | 0.427 | 0.959 | 0.001 |
| Chloride | 0.447 | 0.192 | 0.668 | 0.356 | 0.765 | 0.496 | 0.180 |
| Cholesterol | 0.632 | 0.117 | 0.005 | 0.751 | 0.810 | 0.503 | 0.008 |
| CPK | 0.820 | 0.004 | 0.023 | 0.922 | 0.676 | 0.217 | 0.058 |
| Creatinine | 0.100 | 0.053 | 0.008 | 0.875 | 0.509 | 0.832 | 0.301 |
| GGT | 0.042 | 0.489 | 0.120 | 0.331 | 0.755 | 0.542 | 0.029 |
| GLDH | 0.285 | 0.473 | 0.107 | 0.953 | 0.067 | 0.796 | 0.348 |
| Globulin | 0.430 | 0.581 | 0.098 | 0.694 | 0.630 | 0.305 | 0.667 |
| Glucose | 0.023 | 0.270 | 0.056 | 0.742 | 0.733 | 0.508 | 0.096 |
| NaK Ratio | 0.236 | 0.452 | 0.860 | 0.787 | 0.495 | 0.438 | 0.800 |
| Phosphorus | 0.566 | 0.121 | 0.005 | 0.741 | 0.480 | 0.969 | 0.001 |
| Potassium | 0.108 | 0.278 | 0.159 | 0.825 | 0.455 | 0.686 | 0.732 |
| Protein | 0.095 | 0.001 | 0.298 | 0.916 | 0.553 | 0.250 | 0.097 |
| Sodium | 0.144 | 0.154 | 0.410 | 0.249 | 0.277 | 0.595 | 0.040 |
| Triglycerides | 0.689 | 0.460 | 0.001 | 0.723 | 0.841 | 0.552 | 0.015 |
| Body Weight | 0.168 | 0.083 | 0.040 | 0.131 | 0.025 | 0.418 | 0.051 |
| Analyte 1 | Control Female | PRRSV Female | Control Male | PRRSV Male | Control | PRRSV | Female | Male |
|---|---|---|---|---|---|---|---|---|
| AG Ratio | 0.200 | 0.982 | 0.739 | 0.902 | 0.529 | 0.101 | 0.416 | 0.638 |
| Albumin | 0.692 | 0.560 | 0.698 | 0.536 | 0.077 | 0.052 | 0.871 | 0.425 |
| ALP | 0.427 | 0.945 | 0.122 | 0.226 | 0.692 | 0.010 | 0.860 | 0.251 |
| Anion Gap | 0.764 | 0.176 | 0.152 | 0.021 | 0.746 | 0.572 | 0.118 | 0.026 |
| AST | 0.322 | 0.056 | 0.187 | 0.801 | 0.697 | 0.799 | 0.033 | 0.291 |
| Bicarbonate | 0.242 | 0.623 | 0.961 | 0.187 | 0.751 | 0.737 | 0.571 | 0.009 |
| Bilirubin | 0.145 | 0.584 | 0.062 | 0.074 | 0.070 | 0.146 | 0.180 | 0.006 |
| BUN | 0.027 | 0.020 | 0.003 | 0.041 | 0.688 | 0.866 | 0.011 | 0.001 |
| Calcium | 0.001 | 0.118 | 0.007 | 0.002 | 0.322 | 0.357 | 0.001 | 0.001 |
| Chloride | 0.442 | 0.998 | 0.043 | 0.223 | 0.084 | 0.832 | 0.603 | 0.017 |
| Cholesterol | 0.132 | 0.103 | 0.012 | 0.003 | 0.468 | 0.279 | 0.014 | 0.001 |
| CPK | 0.145 | 0.584 | 0.062 | 0.074 | 0.070 | 0.146 | 0.180 | 0.006 |
| Creatinine | 0.179 | 0.297 | 0.097 | 0.783 | 0.317 | 0.440 | 0.442 | 0.151 |
| GGT | 0.075 | 0.839 | 0.099 | 0.009 | 0.803 | 0.249 | 0.284 | 0.008 |
| GLDH | 0.036 | 0.742 | 0.239 | 0.914 | 0.328 | 0.388 | 0.264 | 0.614 |
| Globulin | 0.247 | 0.857 | 0.459 | 0.373 | 0.628 | 0.915 | 0.531 | 0.193 |
| Glucose | 0.099 | 0.289 | 0.051 | 0.092 | 0.483 | 0.200 | 0.227 | 0.010 |
| NaK Ratio | 0.111 | 0.856 | 0.864 | 0.611 | 0.504 | 0.789 | 0.525 | 0.608 |
| Phosphorus | 0.009 | 0.006 | 0.021 | 0.011 | 0.066 | 0.190 | 0.012 | 0.002 |
| Potassium | 0.070 | 0.868 | 0.986 | 0.802 | 0.393 | 0.634 | 0.671 | 0.983 |
| Protein | 0.803 | 0.479 | 0.259 | 0.122 | 0.030 | 0.076 | 0.708 | 0.031 |
| Sodium | 0.034 | 0.939 | 0.032 | 0.226 | 0.042 | 0.815 | 0.030 | 0.004 |
| Triglycerides | 0.108 | 0.058 | 0.013 | 0.043 | 0.925 | 0.602 | 0.017 | 0.001 |
| Body Weight | 0.019 | 0.029 | 0.234 | 0.280 | 0.339 | 0.115 | 0.096 | 0.259 |
| Analyte 1 | F 2 Sal | F Poly | F Fast | M Sal | M Poly | M Fast | Sal | Poly | Fast | M | F |
|---|---|---|---|---|---|---|---|---|---|---|---|
| AG Ratio | 0.783 | 0.170 | 0.568 | 0.294 | 0.350 | 0.473 | 0.442 | 0.234 | 0.395 | 0.220 | 0.469 |
| Albumin | 0.706 | 0.300 | 0.659 | 0.500 | 0.390 | 0.823 | 0.530 | 0.274 | 0.926 | 0.466 | 0.654 |
| ALP | 0.785 | 0.198 | 0.127 | 0.083 | 0.381 | 0.680 | 0.703 | 0.426 | 0.180 | 0.594 | 0.081 |
| Anion Gap | 0.499 | 0.700 | 0.239 | 0.774 | 0.607 | 0.199 | 0.835 | 0.498 | 0.033 | 0.877 | 0.691 |
| AST | 0.424 | 0.332 | 0.366 | 0.775 | 0.728 | 0.193 | 0.707 | 0.382 | 0.128 | 0.284 | 0.598 |
| Bicarbonate | 0.683 | 0.285 | 0.178 | 0.451 | 0.302 | 0.316 | 0.870 | 0.995 | 0.052 | 0.359 | 0.519 |
| Bilirubin | 0.543 | 0.988 | 0.357 | 0.699 | 0.960 | 0.616 | 0.932 | 0.942 | 0.435 | 0.752 | 0.839 |
| BUN | 0.107 | 0.156 | 0.823 | 0.685 | 0.277 | 0.874 | 0.180 | 0.960 | 0.987 | 0.386 | 0.815 |
| Calcium | 0.955 | 0.129 | 0.745 | 0.905 | 0.380 | 0.417 | 0.948 | 0.112 | 0.733 | 0.390 | 0.438 |
| Chloride | 0.441 | 0.990 | 0.303 | 0.489 | 0.245 | 0.973 | 0.287 | 0.247 | 0.357 | 0.289 | 0.319 |
| Cholesterol | 0.966 | 0.911 | 0.690 | 0.973 | 0.552 | 0.902 | 0.918 | 0.634 | 0.743 | 0.825 | 0.853 |
| CPK | 0.543 | 0.988 | 0.357 | 0.699 | 0.960 | 0.616 | 0.932 | 0.942 | 0.435 | 0.752 | 0.839 |
| Creatinine | 0.642 | 0.136 | 0.239 | 0.176 | 0.735 | 0.770 | 0.255 | 0.594 | 0.539 | 0.523 | 0.732 |
| GGT | 0.079 | 0.549 | 0.909 | 0.270 | 0.066 | 0.080 | 0.098 | 0.379 | 0.208 | 0.032 | 0.434 |
| GLDH | 0.607 | 0.518 | 0.073 | 0.274 | 0.656 | 0.363 | 0.280 | 0.508 | 0.089 | 0.400 | 0.394 |
| Globulin | 0.956 | 0.269 | 0.320 | 0.337 | 0.363 | 0.319 | 0.641 | 0.294 | 0.272 | 0.294 | 0.438 |
| Glucose | 0.292 | 0.072 | 0.773 | 0.668 | 0.140 | 0.875 | 0.312 | 0.884 | 0.941 | 0.590 | 0.953 |
| NaK Ratio | 0.861 | 0.077 | 0.619 | 0.674 | 0.951 | 0.930 | 0.827 | 0.218 | 0.698 | 0.770 | 0.535 |
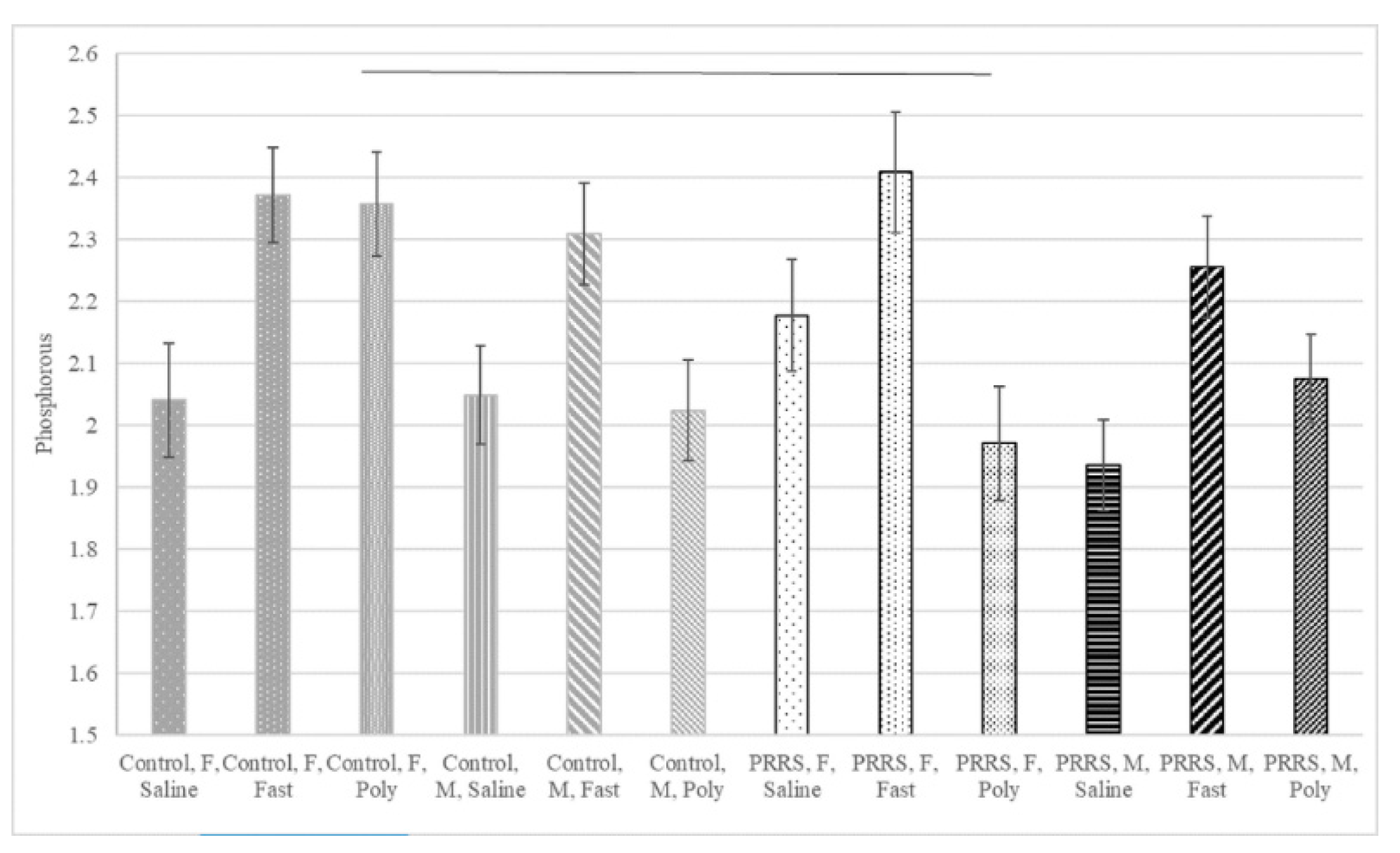
| Phosphorus | 0.290 | 0.004 | 0.764 | 0.306 | 0.635 | 0.641 | 0.899 | 0.158 | 0.924 | 0.655 | 0.430 |
| Potassium | 0.696 | 0.060 | 0.667 | 0.877 | 0.873 | 0.809 | 0.986 | 0.300 | 0.727 | 0.890 | 0.774 |
| Protein | 0.856 | 0.535 | 0.106 | 0.424 | 0.382 | 0.223 | 0.508 | 0.336 | 0.078 | 0.184 | 0.252 |
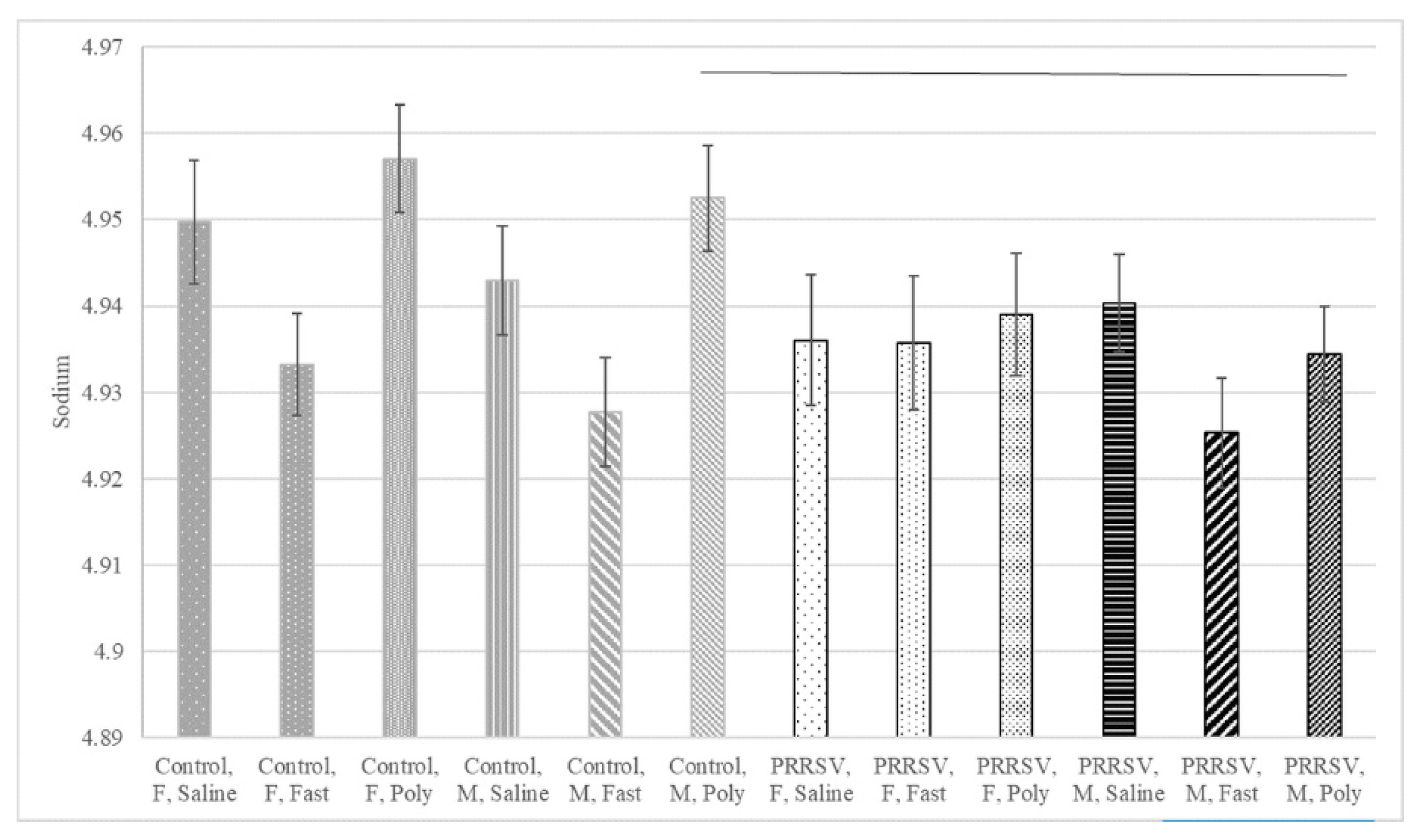
| Sodium | 0.201 | 0.072 | 0.798 | 0.760 | 0.037 | 0.785 | 0.088 | 0.097 | 0.898 | 0.263 | 0.049 |
| Triglycerides | 0.646 | 0.207 | 0.993 | 0.536 | 0.700 | 0.409 | 0.957 | 0.422 | 0.766 | 0.862 | 0.544 |
| Weight | 0.749 | 0.659 | 0.657 | 0.807 | 0.545 | 0.832 | 0.026 | 0.275 | 0.691 | 0.096 | 0.671 |
| Female | Male | |||||||
|---|---|---|---|---|---|---|---|---|
| Analyte 1 | Unit(loge) | Saline 2 | Poly(I:C) | Fasting | Saline | Poly(I:C) | Fasting | SEM |
| AG Ratio | 0.36 | 0.60 | 0.48 | 0.49 | 0.47 | 0.38 | 0.12 | |
| Albumin | g/dL | 1.11 | 1.16 | 1.10 | 1.02 | 1.08 | 1.05 | 0.05 |
| ALP | U/L | 5.40 | 5.55 | 5.56 | 5.37 | 5.61 | 5.58 | 0.10 |
| Anion Gap | mmol/L | 2.72 | 2.72 | 2.78 | 2.65 | 2.78 | 2.78 | 0.06 |
| AST | U/L | 3.57 | 3.89 | 3.38 | 3.77 | 3.68 | 3.21 | 0.24 |
| Bicarbonate | mmol/L | 3.30 | 3.19 | 3.33 | 3.30 | 3.32 | 3.30 | 0.06 |
| Bilirubin | mg/dL | 7.78 | 7.97 | 7.11 | 7.49 | 7.23 | 6.40 | 0.40 |
| BUN | mg/dL | 1.87 a | 2.08 a | 2.52 a,b | 1.90 a | 1.78 a | 2.65 b | 0.17 |
| Calcium | mg/dL | 2.37 b | 2.44 b | 2.25 a | 2.34 a,b | 2.40 b | 2.24 a | 0.03 |
| Chloride | mmol/L | 4.77 a,b | 4.62 a | 4.81 b | 4.62 a | 4.63 a,b | 4.60 a | 0.06 |
| Cholesterol | mg/dL | 4.40 a,b | 4.41 a,b | 4.60 a,b | 4.29 a | 4.36 a,b | 4.63 b | 0.08 |
| CPK | U/L | 7.78 | 7.97 | 7.11 | 7.49 | 7.23 | 6.40 | 0.40 |
| Creatinine | mg/dL | −0.02 a,b | 0.23 b | 0.07 a,b | −0.14 a | 0.01 a,b | 0.12 a,b | 0.09 |
| GGT | U/L | 3.50 | 3.76 | 3.74 | 3.52 | 3.66 | 3.76 | 0.08 |
| GLDH | U/L | −0.18 | 0.41 | −0.42 | 0.01 | 0.47 | 0.04 | 0.28 |
| Globulin | g/dL | 0.73 | 0.56 | 0.63 | 0.55 | 0.60 | 0.68 | 0.09 |
| Glucose | mg/dL | 4.84 | 4.31 | 4.50 | 4.78 | 4.76 | 4.33 | 0.16 |
| NaK Ratio | 3.48 | 3.27 | 3.51 | 3.49 | 3.50 | 3.44 | 0.08 | |
| Phosphorus | mg/dL | 2.04 a | 2.36 a,b | 2.37 b | 2.05 a | 2.02 a | 2.31 a,b | 0.08 |
| Potassium | mmol/L | 1.42 a | 1.68 b | 1.42 a | 1.42 a | 1.42 a | 1.44 a,b | 0.09 |
| Protein | g/dL | 1.64 | 1.61 | 1.61 | 1.52 | 1.56 | 1.59 | 0.04 |
| Sodium | mmol/L | 4.95 | 4.96 | 4.93 | 4.94 | 4.95 | 4.93 | 0.01 |
| Triglycerides | mg/dL | 3.82 a | 4.11 a,b | 5.05 a,b | 3.85 a | 3.79 a | 5.48 b | 0.44 |
| Body Weight | kg | 25.66 | 25.95 | 24.30 | 25.70 | 25.38 | 24.71 | 1.07 |
| Female | Male | |||||||
|---|---|---|---|---|---|---|---|---|
| Analyte 1 | Unit(loge) | Saline 2 | Poly(I:C) | Fasting | Saline | Poly(I:C) | Fasting | SEM |
| AG Ratio | 0.30 | 0.31 | 0.34 | 0.26 | 0.28 | 0.21 | 0.18 | |
| Albumin | g/dL | 1.07 | 1.06 | 1.14 | 0.97 | 1.00 | 1.03 | 0.07 |
| ALP | U/L | 5.35 | 5.34 | 5.30 | 5.63 | 5.73 | 5.52 | 0.11 |
| Anion Gap | mmol/L | 2.79 | 2.68 | 2.90 | 2.63 | 2.74 | 2.90 | 0.07 |
| AST | U/L | 3.24 | 4.27 | 3.72 | 3.86 | 3.81 | 3.67 | 0.24 |
| Bicarbonate | mmol/L | 3.25 | 3.30 | 3.18 | 3.36 | 3.24 | 3.21 | 0.07 |
| Bilirubin | mg/dL | 7.34 a,b | 7.96 b | 7.61 b | 7.72 b | 7.25 a,b | 6.66 a | 0.36 |
| BUN | mg/dL | 2.32 a,b | 1.67 a | 2.58 b | 2.00 a,b | 2.05 a,b | 2.61 b | 0.19 |
| Calcium | mg/dL | 2.37 | 2.36 | 2.26 | 2.34 | 2.36 | 2.20 | 0.03 |
| Chloride | mmol/L | 4.62 | 4.62 | 4.61 | 4.61 | 4.61 | 4.60 | 0.01 |
| Cholesterol | mg/dL | 4.41 a | 4.39 a | 4.65 b | 4.29 a | 4.30 a | 4.64 b | 0.08 |
| CPK | U/L | 7.34 a,b | 7.96 b | 7.61 b | 7.72 b | 7.25 a,b | 6.66 a | 0.36 |
| Creatinine | mg/dL | 0.04 | 0.02 | 0.21 | 0.02 | 0.06 | 0.09 | 0.07 |
| GGT | U/L | 3.75 a,b | 3.69 a | 3.75 a,b | 3.65 a | 3.87 a,b | 3.98 b | 0.08 |
| GLDH | U/L | 0.06 | 0.16 | 0.33 | 0.44 | 0.32 | 0.41 | 0.23 |
| Globulin | g/dL | 0.73 | 0.74 | 0.80 | 0.70 | 0.73 | 0.84 | 0.12 |
| Glucose | mg/dL | 4.63 | 4.84 | 4.44 | 4.71 | 4.37 | 4.36 | 0.15 |
| NaK Ratio | 3.46 | 3.52 | 3.45 | 3.53 | 3.51 | 3.43 | 0.08 | |
| Phosphorus | mg/dL | 2.19 | 1.97 | 2.41 | 1.94 | 2.08 | 2.26 | 0.08 |
| Potassium | mmol/L | 1.47 | 1.41 | 1.48 | 1.40 | 1.40 | 1.47 | 0.09 |
| Protein | g/dL | 1.65 | 1.65 | 1.71 | 1.56 | 1.61 | 1.66 | 0.04 |
| Sodium | mmol/L | 4.94 | 4.94 | 4.94 | 4.94 | 4.93 | 4.93 | 0.01 |
| Triglycerides | mg/dL | 4.13 b | 3.25 a | 5.04 c | 3.49 a,b | 4.01 a,b | 4.97 b,c | 0.43 |
| Body Weight | kg | 25.94 | 25.6 | 23.84 | 25.89 | 25.88 | 24.9 | 1.08 |
| Cytokine | Prenatal Stress | Sex | Postnatal Stress | Prenatal × Sex | Prenatal × Postnatal | Sex × Postnatal | Pre × Postnatal × Sex |
|---|---|---|---|---|---|---|---|
| IFN-γ 1 | 0.420 | 0.461 | 0.249 | 0.189 | 0.009 | 0.008 | 0.028 |
| IL-1α | 0.190 | 0.140 | 0.226 | 0.361 | 0.158 | 0.559 | 0.481 |
| IL-10 | 0.070 | 0.675 | 0.167 | 0.903 | 0.151 | 0.904 | 0.468 |
| IL-12 | 0.838 | 0.964 | 0.027 | 0.652 | 0.789 | 0.981 | 0.543 |
| IL-18 | 0.015 | 0.402 | 0.133 | 0.672 | 0.333 | 0.845 | 0.543 |
| IL-1β | 0.526 | 0.353 | 0.521 | 0.379 | 0.480 | 0.519 | 0.530 |
| IL-2 | 0.030 | 0.589 | 0.310 | 0.854 | 0.188 | 0.711 | 0.832 |
| IL-4 | 0.008 | 0.861 | 0.156 | 0.588 | 0.144 | 0.800 | 0.698 |
| TNF-α | 0.102 | 0.673 | 0.001 | 0.038 | 0.962 | 0.640 | 0.048 |
| Female | Male | ||||||
|---|---|---|---|---|---|---|---|
| Analyte 1 | Saline 2 | Fasting | Poly(I:C) | Saline | Fasting | Poly(I:C) | SEM |
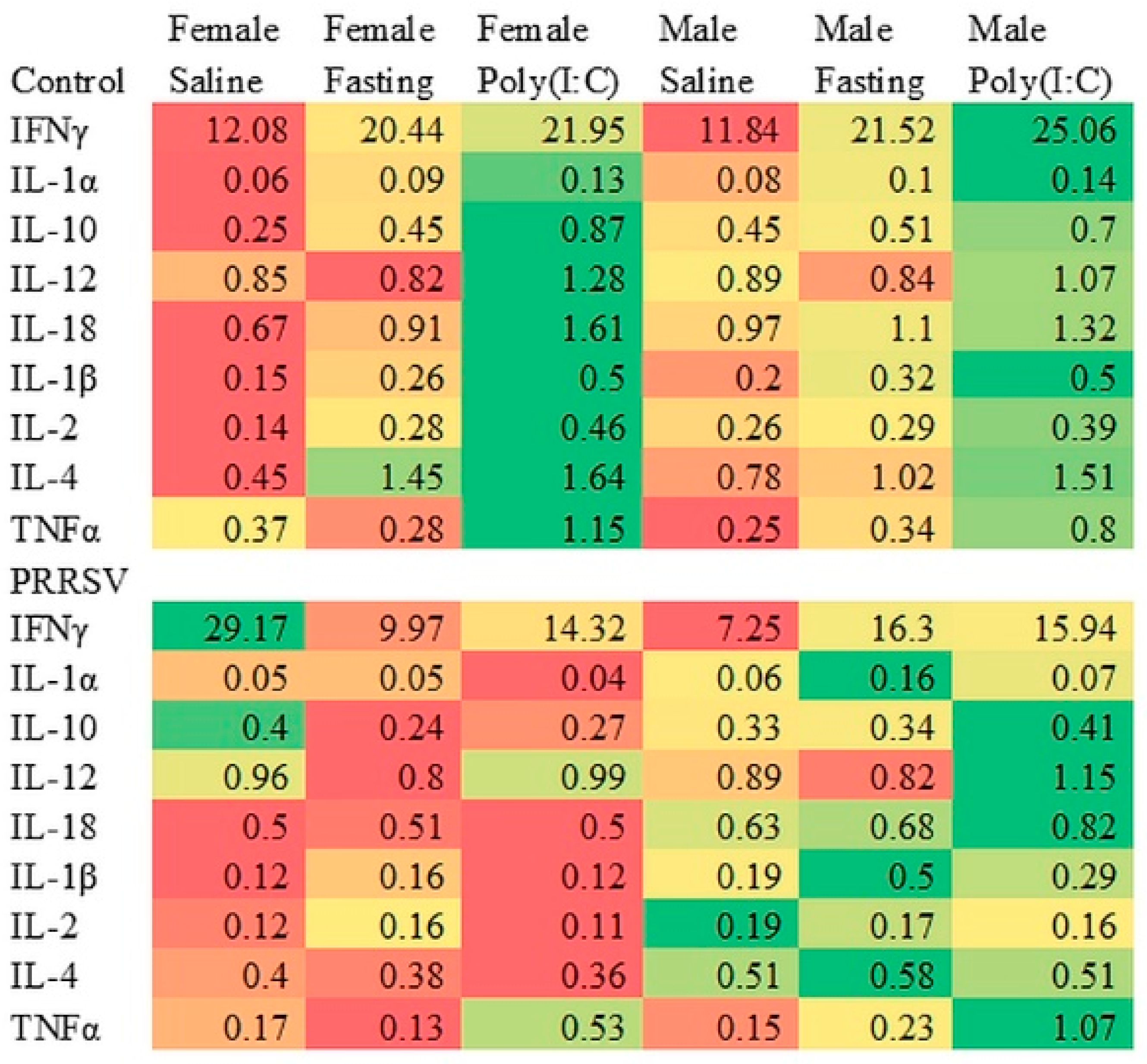
| IFN-γ | 12.08 | 20.44 | 21.95 | 11.84 | 21.52 | 25.06 | 3.61 |
| IL-1α | 0.06 a | 0.09 a,b | 0.13 a,b | 0.08 a,b | 0.1 a,b | 0.14 b | 0.02 |
| IL-10 | 0.25 | 0.45 | 0.87 | 0.45 | 0.51 | 0.7 | 0.20 |
| IL-12 | 0.85 | 0.82 | 1.28 | 0.89 | 0.84 | 1.07 | 0.17 |
| IL-18 | 0.67 | 0.91 | 1.61 | 0.97 | 1.10 | 1.32 | 0.31 |
| IL-1β | 0.15 a | 0.26 a,b | 0.50 b | 0.20 a,b | 0.32 a,b | 0.50 b | 0.10 |
| IL-2 | 0.14 | 0.28 | 0.46 | 0.26 | 0.29 | 0.39 | 0.12 |
| IL-4 | 0.45 | 1.45 | 1.64 | 0.78 | 1.02 | 1.51 | 0.44 |
| TNF-α | 0.37 a | 0.28 a | 1.15 b | 0.25 a | 0.34 a | 0.8 a,b | 0.14 |
| Female | Male | ||||||
|---|---|---|---|---|---|---|---|
| Analyte 1 | Saline 2 | Fasting | Poly(I:C) | Saline | Fasting | Poly(I:C) | SEM |
| IFN-γ | 29.17 b | 9.97 a | 14.32 a,b | 7.25 a | 16.3 a,b | 15.94 a,b | 5.06 |
| IL-1α | 0.05 | 0.05 | 0.04 | 0.06 | 0.16 | 0.07 | 0.04 |
| IL-10 | 0.4 | 0.24 | 0.27 | 0.33 | 0.34 | 0.41 | 0.15 |
| IL-12 | 0.96 | 0.8 | 0.99 | 0.89 | 0.82 | 1.15 | 0.17 |
| IL-18 | 0.5 | 0.51 | 0.5 | 0.63 | 0.68 | 0.82 | 0.26 |
| IL-1β | 0.12 | 0.16 | 0.12 | 0.19 | 0.50 | 0.29 | 0.10 |
| IL-2 | 0.12 | 0.16 | 0.11 | 0.19 | 0.17 | 0.16 | 0.08 |
| IL-4 | 0.4 | 0.38 | 0.36 | 0.51 | 0.58 | 0.51 | 0.27 |
| TNF-α | 0.17 a | 0.13 a | 0.53 a,b | 0.15 a | 0.23 a | 1.07 b | 0.15 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rymut, H.E.; Rund, L.A.; Bolt, C.R.; Villamil, M.B.; Bender, D.E.; Southey, B.R.; Johnson, R.W.; Rodriguez-Zas, S.L. Biochemistry and Immune Biomarkers Indicate Interacting Effects of Pre- and Postnatal Stressors in Pigs across Sexes. Animals 2021, 11, 987. https://doi.org/10.3390/ani11040987
Rymut HE, Rund LA, Bolt CR, Villamil MB, Bender DE, Southey BR, Johnson RW, Rodriguez-Zas SL. Biochemistry and Immune Biomarkers Indicate Interacting Effects of Pre- and Postnatal Stressors in Pigs across Sexes. Animals. 2021; 11(4):987. https://doi.org/10.3390/ani11040987
Chicago/Turabian StyleRymut, Haley E., Laurie A. Rund, Courtni R. Bolt, María B. Villamil, Diane E. Bender, Bruce R. Southey, Rodney W. Johnson, and Sandra L. Rodriguez-Zas. 2021. "Biochemistry and Immune Biomarkers Indicate Interacting Effects of Pre- and Postnatal Stressors in Pigs across Sexes" Animals 11, no. 4: 987. https://doi.org/10.3390/ani11040987
APA StyleRymut, H. E., Rund, L. A., Bolt, C. R., Villamil, M. B., Bender, D. E., Southey, B. R., Johnson, R. W., & Rodriguez-Zas, S. L. (2021). Biochemistry and Immune Biomarkers Indicate Interacting Effects of Pre- and Postnatal Stressors in Pigs across Sexes. Animals, 11(4), 987. https://doi.org/10.3390/ani11040987