Bacteriocins from Lactic Acid Bacteria. A Powerful Alternative as Antimicrobials, Probiotics, and Immunomodulators in Veterinary Medicine

 , and
, and
Simple Summary
Abstract
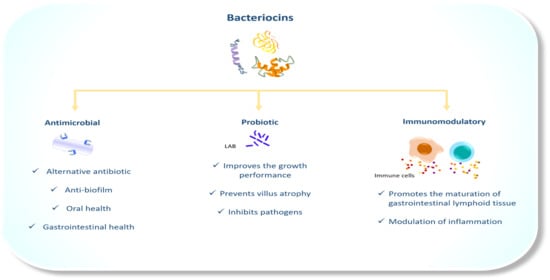
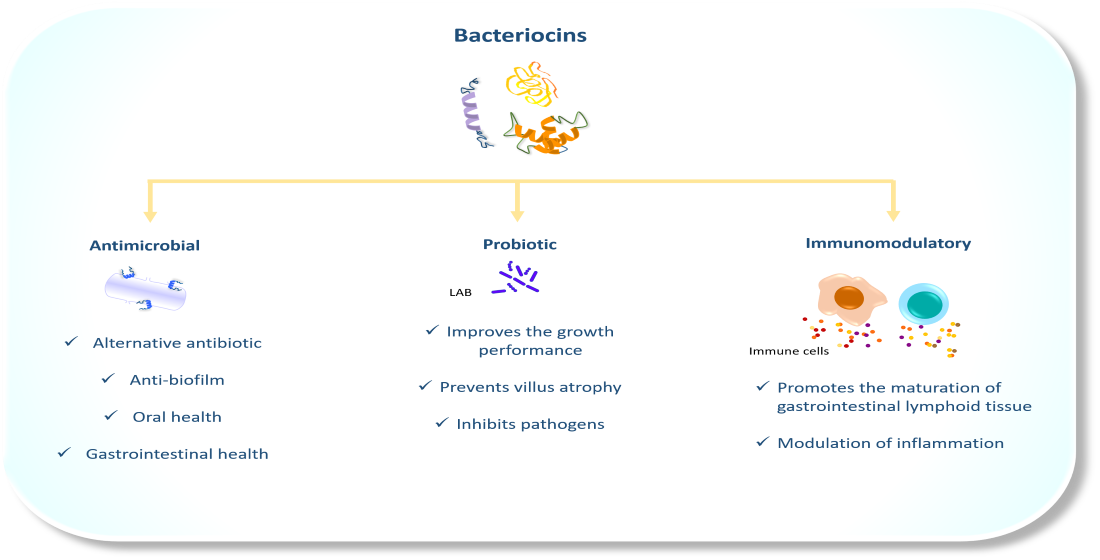
1. Introduction
2. Bacteriocin Overview
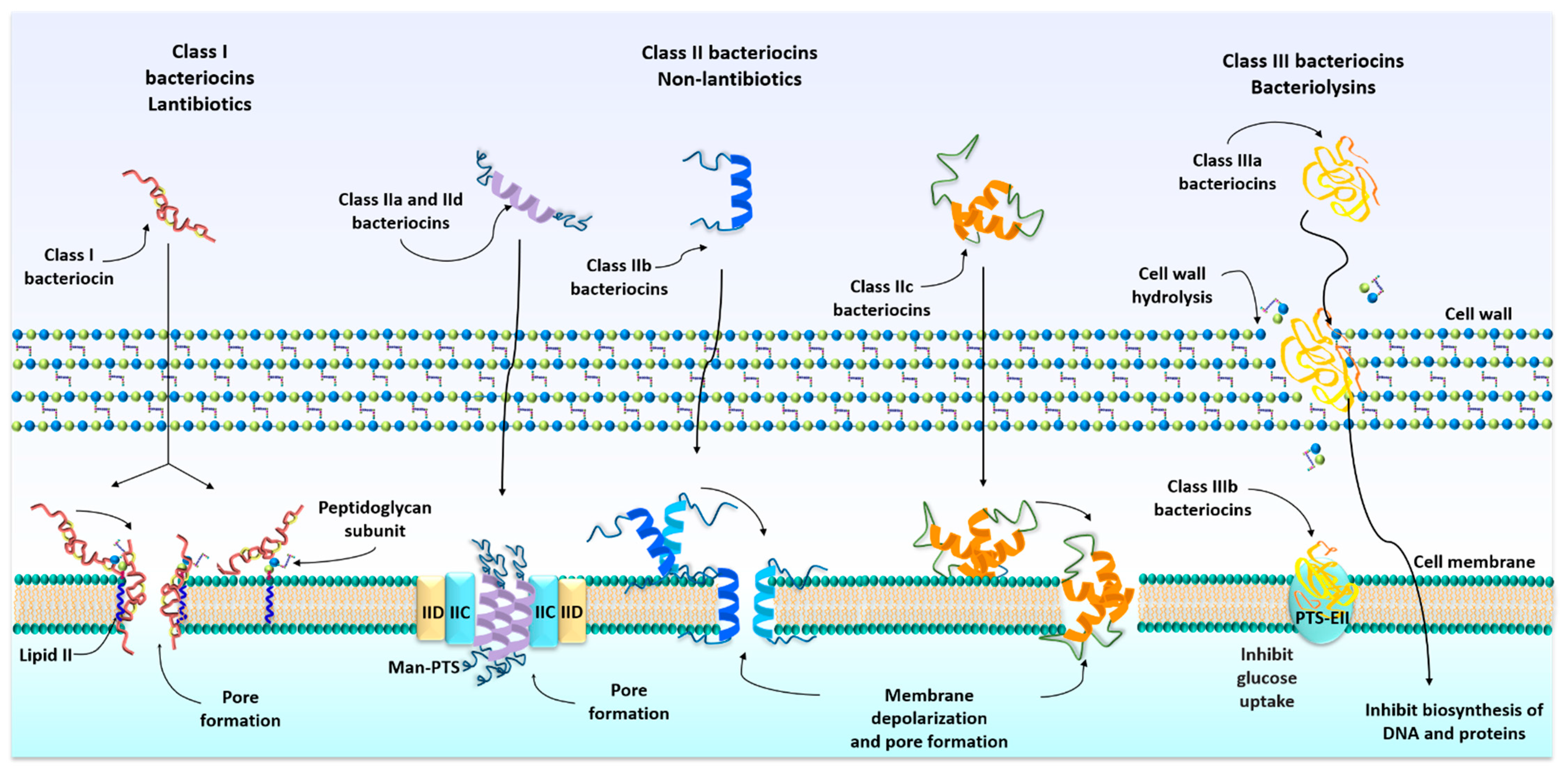
3. Classification of Bacteriocins
4. Mechanism of Action of Bacteriocins
5. The Antimicrobial Properties of Bacteriocins in Veterinary Medicine
6. The Probiotic Activity of LAB Bacteriocins
7. Bacteriocins as Immunomodulators
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zheng, J.; Gänzle, M.G.; Lin, X.B.; Ruan, L.; Sun, M. Diversity and Dynamics of Bacteriocins from Human Microbiome. Environ. Microbiol. 2014, 17, 2133–2143. [Google Scholar] [CrossRef]
- Torres, N.I.; Noll, K.S.; Xu, S.; Li, J.; Huang, Q.; Sinko, P.J.; Wachsman, M.B.; Chikindas, M.L. Safety, Formulation and In Vitro Antiviral Activity of the Antimicrobial Peptide Subtilosin Against Herpes Simplex Virus Type 1. Probiotics Antimicrob. Proteins 2013, 5, 26–35. [Google Scholar] [CrossRef]
- Eyang, S.-C.; Elin, C.-H.; Sung, C.T.; Efang, J.-Y. Antibacterial Activities of Bacteriocins: Application in Foods and Pharmaceuticals. Front. Microbiol. 2014, 5, 241. [Google Scholar] [CrossRef]
- Graham, C.E.; Cruz, M.R.; Garsin, D.A.; Lorenz, M.C. Enterococcus Faecalisbacteriocin EntV Inhibits Hyphal Morphogenesis, Biofilm Formation, and Virulence of Candida Albicans. Proc. Natl. Acad. Sci. USA 2017, 114, 4507–4512. [Google Scholar] [CrossRef] [PubMed]
- Martín-Escolano, R.; Cebrián, R.; Martín-Escolano, J.; Rosales, M.J.; Maqueda, M.; Sánchez-Moreno, M.; Marín, C. Insights into Chagas Treatment Based on the Potential of Bacteriocin AS-48. Int. J. Parasitol. Drugs Drug Resist. 2019, 10, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Mokoena, M.P. Lactic Acid Bacteria and Their Bacteriocins: Classification, Biosynthesis and Applications against Uropathogens: A Mini-Review. Molecules 2017, 22, 1255. [Google Scholar] [CrossRef] [PubMed]
- Lahtinen, S.; Ouwehand, A.C.; Salminen, S.; Wright, A. Von Lactic Acid Bacteria: Microbiological and Functional Aspects, 4th ed.; CRC Press: Boca Raton, FL, USA, 2011; ISBN 978-1-4398-3678-1. [Google Scholar]
- Agriopoulou, S.; Stamatelopoulou, E.; Sachadyn-Król, M.; Varzakas, T. Lactic Acid Bacteria as Antibacterial Agents to Extend the Shelf Life of Fresh and Minimally Processed Fruits and Vegetables: Quality and Safety Aspects. Microorganisms 2020, 8, 952. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Pang, H.; Zhang, H.; Cai, Y. Biodiversity of Lactic Acid Bacteria. In Lactic Acid Bacteria; J.B. Metzler, Ed.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 103–203. [Google Scholar]
- Kheadr, E.; Zihler, A.; Dabour, N.; Lacroix, C.; Le Blay, G.; Fliss, I. Study of the Physicochemical and Biological Stability of Pediocin PA-1 in the Upper Gastrointestinal Tract Conditions Using a Dynamicin Vitromodel. J. Appl. Microbiol. 2010, 109, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.C.G.; Silva, S.P.M.; Ribeiro, S.C. Application of Bacteriocins and Protective Cultures in Dairy Food Preservation. Front. Microbiol. 2018, 9, 594. [Google Scholar] [CrossRef]
- Maurício, E.; Rosado, C.; Duarte, M.P.; Verissimo, J.; Bom, S.; Vasconcelos, L. Efficiency of Nisin as Preservative in Cosmetics and Topical Products. Cosmetics 2017, 4, 41. [Google Scholar] [CrossRef]
- Bédard, F.; Biron, E. Recent Progress in the Chemical Synthesis of Class II and S-Glycosylated Bacteriocins. Front. Microbiol. 2018, 9, 1048. [Google Scholar] [CrossRef] [PubMed]
- Dowarah, R.; Verma, A.; Agarwal, N. The Use of Lactobacillus as an Alternative of Antibiotic Growth Promoters in Pigs: A Review. Anim. Nutr. 2017, 3, 1–6. [Google Scholar] [CrossRef]
- Vieco-Saiz, N.; Belguesmia, Y.; Raspoet, R.; Auclair, E.; Gancel, F.; Kempf, I.; Drider, D. Benefits and Inputs from Lactic Acid Bacteria and Their Bacteriocins as Alternatives to Antibiotic Growth Promoters During Food-Animal Production. Front. Microbiol. 2019, 10, 57. [Google Scholar] [CrossRef] [PubMed]
- Riley, M.A.; Wertz, J.E. Bacteriocins: Evolution, Ecology, and Application. Annu. Rev. Microbiol. 2002, 56, 117–137. [Google Scholar] [CrossRef] [PubMed]
- Ben Lagha, A.; Haas, B.; Gottschalk, M.; Grenier, D. Antimicrobial Potential of Bacteriocins in Poultry and Swine Production. Veter. Res. 2017, 48, 1–12. [Google Scholar] [CrossRef]
- Noda, M.; Miyauchi, R.; Danshiitsoodol, N.; Matoba, Y.; Kumagai, T.; Sugiyama, M. Expression of Genes Involved in Bacteriocin Production and Self-Resistance inLactobacillus brevis174A Is Mediated by Two Regulatory Proteins. Appl. Environ. Microbiol. 2018, 84, e02707-17. [Google Scholar] [CrossRef] [PubMed]
- Zimina, M.; Babich, O.; Prosekov, A.; Sukhikh, S.; Ivanova, S.; Shevchenko, M.; Noskova, S. Overview of Global Trends in Classification, Methods of Preparation and Application of Bacteriocins. Antibiotics 2020, 9, 553. [Google Scholar] [CrossRef] [PubMed]
- Kumariya, R.; Garsa, A.K.; Rajput, Y.; Sood, S.; Akhtar, N.; Patel, S. Bacteriocins: Classification, Synthesis, Mechanism of Action and Resistance Development in Food Spoilage Causing Bacteria. Microb. Pathog. 2019, 128, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Bédard, F.; Hammami, R.; Zirah, S.; Rebuffat, S.; Fliss, I.; Biron, E. Synthesis, Antimicrobial Activity and Conformational Analysis of the Class IIa Bacteriocin Pediocin PA-1 and Analogs Thereof. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Schofs, L.; Sparo, M.D.; Bruni, S.F.S. Gram-Positive Bacteriocins: Usage as Antimicrobial Agents in Veterinary Medicine. Veter. Res. Commun. 2020, 44, 1–12. [Google Scholar] [CrossRef]
- Meade, E.; Slattery, M.A.; Garvey, M. Bacteriocins, Potent Antimicrobial Peptides and the Fight against Multi Drug Resistant Species: Resistance Is Futile? Antibiotics 2020, 9, 32. [Google Scholar] [CrossRef] [PubMed]
- Siegers, K.; Entian, K.D. Genes Involved in Immunity to the Lantibiotic Nisin Produced by Lactococcus Lactis 6F3. Appl. Environ. Microbiol. 1995, 61, 1082–1089. [Google Scholar] [CrossRef] [PubMed]
- Alkhatib, Z.; Abts, A.; Mavaro, A.; Schmitt, L.; Smits, S.H. Lantibiotics: How Do Producers Become Self-Protected? J. Biotechnol. 2012, 159, 145–154. [Google Scholar] [CrossRef]
- Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocins—A Viable Alternative to Antibiotics? Nat. Rev. Genet. 2013, 11, 95–105. [Google Scholar] [CrossRef]
- Cavera, V.L.; Arthur, T.D.; Kashtanov, D.; Chikindas, M.L. Bacteriocins and Their Position in the Next Wave of Conventional Antibiotics. Int. J. Antimicrob. Agents 2015, 46, 494–501. [Google Scholar] [CrossRef]
- Bacteriocins as a New Generation of Antimicrobials: Toxicity Aspects and Regulations|FEMS Microbiology Reviews, Oxford Academic. Available online: https://academic.oup.com/femsre/article/45/1/fuaa039/5900525 (accessed on 18 March 2021).
- Sakayori, Y.; Muramatsu, M.; Hanada, S.; Kamagata, Y.; Kawamoto, S.; Shima, J. Characterization of Enterococcus Faecium Mutants Resistant to Mundticin KS, a Class IIa Bacteriocin. Microbiology 2003, 149, 2901–2908. [Google Scholar] [CrossRef]
- McBride, S.M.; Sonenshein, A.L. The dlt Operon Confers Resistance to Cationic Antimicrobial Peptides in Clostridium Difficile. Microbiology 2011, 157, 1457–1465. [Google Scholar] [CrossRef]
- Hammami, R.; Zouhir, A.; Ben Hamida, J.; Fliss, I. Bactibase: A New Web-Accessible Database for Bacteriocin Characterization. BMC Microbiol. 2007, 7, 89. [Google Scholar] [CrossRef]
- Al Kassaa, I.; Rafei, R.; Moukhtar, M.; Zaylaa, M.; Gharsallaoui, A.; Asehraou, A.; El Omari, K.; Shahin, A.; Hamze, M.; Chihib, N.-E. LA Biocin Database: A New Database Designed Specifically for Lactic Acid Bacteria Bacteriocins. Int. J. Antimicrob. Agents 2019, 54, 771–779. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, V.; Khan, M.S.; Jamal, Q.M.S.; Alzohairy, M.A.; Al Karaawi, M.A.; Siddiqui, M.U. Antimicrobial Potential of Bacteriocins: In Therapy, Agriculture and Food Preservation. Int. J. Antimicrob. Agents 2017, 49, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Klaenhammer, T.R. Genetics of Bacteriocins Produced by Lactic Acid Bacteria. FEMS Microbiol. Rev. 1993, 12, 39–85. [Google Scholar] [CrossRef]
- Güllüce, M.; Karadayı, M.; Bariş, Ö. Bacteriocins: Promising Natural Antimicrobials. In Science, Technology and Education; Mendes-Vilas, A., Ed.; Formatex: Madrid, Spain, 2013; pp. 1016–1027. [Google Scholar]
- Guder, A.; Wiedemann, I.; Sahl, H.G. Posttranslationally Modified Bacteriocins—The Lantibiotics. Biopolymers 2000, 55, 62–73. [Google Scholar] [CrossRef]
- Bierbaum, G.; Götz, F.; Peschel, A.; Kupke, T.; Van De Kamp, M.; Sahl, H.-G. The Biosynthesis of the Lantibiotics Epidermin, Gallidermin, Pep5 and Epilancin K7. Antonie Leeuwenhoek 1996, 69, 119–127. [Google Scholar] [CrossRef]
- Alvarez-Sieiro, P.; Montalbán-López, M.; Mu, D.; Kuipers, O.P. Bacteriocins of Lactic Acid Bacteria: Extending the Family. Appl. Microbiol. Biotechnol. 2016, 100, 2939–2951. [Google Scholar] [CrossRef] [PubMed]
- Deegan, L.H.; Cotter, P.D.; Hill, C.; Ross, P. Bacteriocins: Biological Tools for Bio-Preservation and Shelf-Life Extension. Int. Dairy, J. 2006, 16, 1058–1071. [Google Scholar] [CrossRef]
- Fimland, G.; Johnsen, L.; Dalhus, B.; Nissen-Meyer, J. Pediocin-like Antimicrobial Peptides (Class IIa Bacteriocins) and Their Immunity Proteins: Biosynthesis, Structure, and Mode of Action. J. Pept. Sci. 2005, 11, 688–696. [Google Scholar] [CrossRef] [PubMed]
- Nissen-Meyer, J.; Oppegård, C.; Rogne, P.; Haugen, H.S.; Kristiansen, P.E. Structure and Mode-of-Action of the Two-Peptide (Class-IIb) Bacteriocins. Probiotics Antimicrob. Proteins 2009, 2, 52–60. [Google Scholar] [CrossRef]
- Oppegård, C.; Rogne, P.; Emanuelsen, L.; Kristiansen, P.E.; Fimland, G.; Nissen-Meyer, J. The Two-Peptide Class II Bacteriocins: Structure, Production, and Mode of Action. J. Mol. Microbiol. Biotechnol. 2007, 13, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Sawa, N.; Zendo, T.; Kiyofuji, J.; Fujita, K.; Himeno, K.; Nakayama, J.; Sonomoto, K. Identification and Characterization of Lactocyclicin Q, a Novel Cyclic Bacteriocin Produced by Lactococcus sp. Strain QU 12. Appl. Environ. Microbiol. 2009, 75, 1552–1558. [Google Scholar] [CrossRef]
- Borrero, J.; Brede, D.A.; Skaugen, M.; Diep, D.B.; Herranz, C.; Nes, I.F.; Cintas, L.M.; Hernández, P.E. Characterization of Garvicin ML, a Novel Circular Bacteriocin Produced by Lactococcus garvieaeDCC43, Isolated from Mallard Ducks (Anas platyrhynchos). Appl. Environ. Microbiol. 2010, 77, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Gabrielsen, C.; Brede, D.A.; Nes, I.F.; Diep, D.B. Circular Bacteriocins: Biosynthesis and Mode of Action. Appl. Environ. Microbiol. 2014, 80, 6854–6862. [Google Scholar] [CrossRef] [PubMed]
- Iwatani, S.; Zendo, T.; Sonomoto, K. Class IId or Linear and Non-Pediocin-Like Bacteriocins. Prokaryotic Antimicrob. Peptides 2011, 237–252. [Google Scholar] [CrossRef]
- Nilsen, T.; Nes, I.F.; Holo, H. Enterolysin A, a Cell Wall-Degrading Bacteriocin from Enterococcus faecalis LMG 2333. Appl. Environ. Microbiol. 2003, 69, 2975–2984. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Wang, X.; Zhang, X.; Wu, H.; Zou, Y.; Li, P.; Sun, C.; Xu, W.; Liu, F.; Wang, D. Class III bacteriocin Helveticin-M Causes Sublethal Damage on Target Cells through Impairment of Cell Wall and Membrane. J. Ind. Microbiol. Biotechnol. 2018, 45, 213–227. [Google Scholar] [CrossRef]
- Bauer, R.; Dicks, L. Mode of Action of Lipid II-Targeting Lantibiotics. Int. J. Food Microbiol. 2005, 101, 201–216. [Google Scholar] [CrossRef]
- Cotter, P.D.; Hill, C.; Ross, R.P. Bacteriocins: Developing Innate Immunity for Food. Nat. Rev. Genet. 2005, 3, 777–788. [Google Scholar] [CrossRef]
- Paiva, A.D.; Breukink, E.; Mantovani, H.C. Role of Lipid II and Membrane Thickness in the Mechanism of Action of the Lantibiotic Bovicin HC5. Antimicrob. Agents Chemother. 2011, 55, 5284–5293. [Google Scholar] [CrossRef] [PubMed]
- Diep, D.B.; Skaugen, M.; Salehian, Z.; Holo, H.; Nes, I.F. Common Mechanisms of Target Cell Recognition and Immunity for Class II Bacteriocins. Proc. Natl. Acad. Sci. USA 2007, 104, 2384–2389. [Google Scholar] [CrossRef]
- Nes, I.F.; Brede, D.A.; Diep, D.B. Chapter 16—Class II Non-Lantibiotic Bacteriocins. In Handbook of Biologically Active Peptides, 2nd ed.; Kastin, A.J., Ed.; Academic Press: Boston, MA, USA, 2013; pp. 85–92. ISBN 978-0-12-385095-9. [Google Scholar]
- Moll, G.; Hildeng-Hauge, H.; Nissen-Meyer, J.; Nes, I.F.; Konings, W.N.; Driessen, A.J.M. Mechanistic Properties of the Two-Component Bacteriocin lactococcin. G. J. Bacteriol. 1998, 180, 96–99. [Google Scholar] [CrossRef] [PubMed]
- Van Belkum, M.J.; Martin-Visscher, L.A.; Vederas, J.C. Structure and Genetics of Circular Bacteriocins. Trends Microbiol. 2011, 19, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Perez, R.H.; Zendo, T.; Sonomoto, K. Circular and Leaderless Bacteriocins: Biosynthesis, Mode of Action, Applications, and Prospects. Front. Microbiol. 2018, 9, 2085. [Google Scholar] [CrossRef]
- Simmonds, R.S.; Pearson, L.; Kennedy, R.C.; Tagg, J.R. Mode of Action of a Lysostaphin-like Bacteriolytic Agent Produced by Streptococcus zooepidemicus 4881. Appl. Environ. Microbiol. 1996, 62, 4536–4541. [Google Scholar] [CrossRef] [PubMed]
- Müller, E.; Radler, F. Caseicin, a Bacteriocin from Lactobacillus casei. Folia Microbiol. 1993, 38, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Swe, P.M.; Cook, G.M.; Tagg, J.R.; Jack, R.W. Mode of Action of Dysgalacticin: A Large Heat-Labile Bacteriocin. J. Antimicrob. Chemother. 2009, 63, 679–686. [Google Scholar] [CrossRef] [PubMed]
- Preciado, G.M.; Michel, M.M.; Villarreal-Morales, S.L.; Flores-Gallegos, A.C.; Aguirre-Joya, J.; Morlett-Chávez, J.; Aguilar, C.N.; Rodríguez-Herrera, R. Chapter 16—Bacteriocins and Its Use for Multidrug-Resistant Bacteria Control. In Antibiotic Resistance; Kon, K., Rai, M., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 329–349. ISBN 978-0-12-803642-6. [Google Scholar]
- De Arauz, L.J.; Jozala, A.F.; Mazzola, P.G.; Penna, T.C.V. Nisin Biotechnological Production and Application: A Review. Trends Food Sci. Technol. 2009, 20, 146–154. [Google Scholar] [CrossRef]
- Howell, T.H.; Fiorellini, J.P.; Blackburn, P.; Projan, S.J.; De La Harpe, J.; Williams, R.C. The Effect of a Mouthrinse Based on Nisin, a Bacteriocin, on Developing Plaque and Gingivitis in Beagle Dogs. J. Clin. Periodontol. 1993, 20, 335–339. [Google Scholar] [CrossRef]
- Cunha, E.; Trovão, T.; Pinheiro, A.; Nunes, T.; Santos, R.; Da Silva, J.M.; Braz, B.S.; Tavares, L.; Veiga, A.S.; Oliveira, M. Potential of Two Delivery Systems for Nisin Topical Application to Dental Plaque Biofilms in Dogs. BMC Vet. Res. 2018, 14, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Harvey, C.E. Periodontal Disease in Dogs. Vet. Clin. N. Am. Small Anim. Pract. 1998, 28, 1111–1128. [Google Scholar] [CrossRef]
- Oliveira, M.; Tavares, M.; Gomes, D.; Touret, T.; Braz, B.S.; Tavares, L.; Semedo-Lemsaddek, T. Virulence Traits and Antibiotic Resistance among Enterococci Isolated from Dogs with Periodontal Disease. Comp. Immunol. Microbiol. Infect. Dis. 2016, 46, 27–31. [Google Scholar] [CrossRef]
- Cunha, E.; Rebelo, S.; Carneiro, C.; Tavares, L.; Carreira, L.M.; Oliveira, M. A Polymicrobial Biofilm Model for Testing the Antimicrobial Potential of a Nisin-Biogel for Canine Periodontal Disease Control. BMC Vet. Res. 2020, 16, 1–10. [Google Scholar] [CrossRef]
- Iseppi, R.; Messi, P.; Anacarso, I.; Bondi, M.; Sabia, C.; Condò, C.; de Niederhausern, S. Antimicrobial Resistance and Virulence Traits in Enterococcus Strains Isolated from Dogs and Cats. New Microbiol. 2015, 38, 369–378. [Google Scholar] [PubMed]
- Bertelloni, F.; Salvadori, C.; Lotti, G.; Cerri, D.; Ebani, V.V. Antimicrobial Resistance in Enterococcus Strains Isolated from Healthy Domestic Dogs. Acta Microbiol. Immunol. Hung. 2016, 64, 301–312. [Google Scholar] [CrossRef]
- Zoonoses. Available online: https://www.who.int/news-room/fact-sheets/detail/zoonoses (accessed on 16 February 2021).
- Cunha, E.; Freitas, F.B.; Braz, B.S.; Da Silva, J.M.; Tavares, L.; Veiga, A.S.; Oliveira, M. Polyphasic Validation of a Nisin-Biogel to Control Canine Periodontal Disease. Antibiotics 2020, 9, 180. [Google Scholar] [CrossRef] [PubMed]
- Hamp, S.-E.; Lindhe, J.; Löe, H. Long Term Effect of Chlorhexidine on Developing Gingivitis in the Beagle Dog. J. Periodontal Res. 1973, 8, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Charbonneau, D.; Snider, A. Reduced Chlorhexidine Tooth Stain Coverage by Sequential Administration of Monoperoxyphthalic Acid in the Beagle Dog. J. Dent. Res. 1997, 76, 1596–1601. [Google Scholar] [CrossRef]
- Damborg, P.; Top, J.; Hendrickx, A.P.A.; Dawson, S.; Willems, R.J.L.; Guardabassi, L. Dogs Are a Reservoir of Ampicillin-Resistant Enterococcus faecium Lineages Associated with Human Infections. Appl. Environ. Microbiol. 2009, 75, 2360–2365. [Google Scholar] [CrossRef] [PubMed]
- Furlaneto-Maia, L.; Ramalho, R.; Rocha, K.R.; Furlaneto, M.C. Antimicrobial Activity of Enterocins against Listeria SP. and Other Food Spoilage Bacteria. Biotechnol. Lett. 2020, 42, 797–806. [Google Scholar] [CrossRef]
- Kubašová, I.; Diep, D.B.; Ovchinnikov, K.V.; Lauková, A.; Strompfová, V. Bacteriocin Production and Distribution of Bacteriocin-Encoding Genes in Enterococci from Dogs. Int. J. Antimicrob. Agents 2020, 55, 105859. [Google Scholar] [CrossRef]
- Taylor, J.; Hirsh, A.; Mattick, A. The Treatment of Bovine Streptococcal and Staphylococcal Mastitis with Nisin. Vet. Res. 1949, 61, 197–198. [Google Scholar]
- Broadbent, J.; Chou, Y.; Gillies, K.; Kondo, J. Nisin Inhibits Several Gram-Positive, Mastitis-Causing Pathogens. J. Dairy Sci. 1989, 72, 3342–3345. [Google Scholar] [CrossRef]
- Cao, L.; Wu, J.; Xie, F.; Hu, S.; Mo, Y. Efficacy of Nisin in Treatment of Clinical Mastitis in Lactating Dairy Cows. J. Dairy Sci. 2007, 90, 3980–3985. [Google Scholar] [CrossRef]
- Wu, J.; Hu, S.; Cao, L. Therapeutic Effect of Nisin Z on Subclinical Mastitis in Lactating Cows. Antimicrob. Agents Chemother. 2007, 51, 3131–3135. [Google Scholar] [CrossRef]
- Feitosa, E.; Bonassi, N.M.; Loh, W. Vesicle−Micelle Transition in Mixtures of Dioctadecyldimethylammonium Chloride and Bromide with Nonionic and Zwitterionic Surfactants. Langmuir 2006, 22, 4512–4517. [Google Scholar] [CrossRef] [PubMed]
- Castelani, L.; Arcaro, J.; Braga, J.; Bosso, A.; Moura, Q.; Esposito, F.; Sauter, I.; Cortez, M.; Lincopan, N. Short Communication: Activity of Nisin, Lipid Bilayer Fragments and Cationic Nisin-Lipid Nanoparticles against Multi-Drug-Resistant Staphylococcus spp. Isolated from Bovine Mastitis. J. Dairy Sci. 2019, 102, 678–683. [Google Scholar] [CrossRef]
- Xue, T.; Chen, X.; Shang, F. Short Communication: Effects of Lactose and Milk on the Expression of Biofilm-Associated Genes in Staphylococcus Aureus Strains Isolated from a Dairy Cow with Mastitis. J. Dairy Sci. 2014, 97, 6129–6134. [Google Scholar] [CrossRef] [PubMed]
- Ceotto-Vigoder, H.; Marques, S.L.S.; Santos, I.N.S.; Alves, M.D.B.; Barrias, E.S.; Potter, A.; Alviano, D.S.; Bastos, M.C.F. Nisin and Lysostaphin Activity against Preformed Biofilm of Staphylococcus aureus Involved in Bovine Mastitis. J. Appl. Microbiol. 2016, 121, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Andre, C.; Pimentel-Filho, N.D.J.; Costa, P.M.D.A.; Vanetti, M.C.D. Changes in the Composition and Architecture of Staphylococcal Biofilm by Nisin. Braz. J. Microbiol. 2019, 50, 1083–1090. [Google Scholar] [CrossRef]
- LeBel, G.; Piché, F.; Frenette, M.; Gottschalk, M.; Grenier, D. Antimicrobial Activity of Nisin against the Swine Pathogen Streptococcus suis and Its Synergistic Interaction with Antibiotics. Positron 2013, 50, 19–23. [Google Scholar] [CrossRef]
- Zhu, H.; Han, L.; Ni, Y.; Yu, Z.; Wang, D.; Zhou, J.; Li, B.; Zhang, W.; He, K. In vitro and In vivo Antibacterial Effects of Nisin Against Streptococcus suis. Probiotics Antimicrob. Proteins 2021, 1–13. [Google Scholar] [CrossRef]
- Campion, A.; Casey, P.G.; Field, D.; Cotter, P.D.; Hill, C.; Ross, R.P. In vivo activity of Nisin A and Nisin V against Listeria monocytogenes in mice. BMC Microbiol. 2013, 13, 23. [Google Scholar] [CrossRef]
- Lebel, G.; Vaillancourt, K.; Frenette, M.; Gottschalk, M.; Grenier, D. Suicin 90-1330 from a Nonvirulent Strain of Streptococcus suis: A Nisin-Related Lantibiotic Active on Gram-Positive Swine Pathogens. Appl. Environ. Microbiol. 2014, 80, 5484–5492. [Google Scholar] [CrossRef] [PubMed]
- Vaillancourt, K.; Lebel, G.; Frenette, M.; Fittipaldi, N.; Gottschalk, M.; Grenier, D. Purification and Characterization of Suicin 65, a Novel Class I Type B Lantibiotic Produced by Streptococcus suis. PLoS ONE 2015, 10, e0145854. [Google Scholar] [CrossRef]
- Vaillancourt, K.; Lebel, G.; Frenette, M.; Gottschalk, M.; Grenier, D. Suicin 3908, a New Lantibiotic Produced by a Strain of Streptococcus suis Serotype 2 Isolated from a Healthy Carrier Pig. PLoS ONE 2015, 10, e0117245. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Gao, Y.; Teng, K.; Zhang, J.; Sun, S.; Zhong, J. Restoration of Bioactive Lantibiotic Suicin from a Remnantlan Locus of Pathogenic Streptococcus suis Serotype 2. Appl. Environ. Microbiol. 2013, 80, 1062–1071. [Google Scholar] [CrossRef] [PubMed]
- Bin, P.; Tang, Z.; Liu, S.; Chen, S.; Xia, Y.; Liu, J.; Wu, H.; Zhu, G. Intestinal Microbiota Mediates Enterotoxigenic Escherichia Coli-Induced Diarrhea in Piglets. BMC Vet. Res. 2018, 14, 1–13. [Google Scholar] [CrossRef]
- Liu, Y.-Y.; Wang, Y.; Walsh, T.R.; Yi, L.-X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of PlasMid-mediated Colistin Resistance Mechanism MCR-1 in Animals and Human Beings in China: A Microbiological and Molecular Biological Study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Aguirre, L.; Vidal, A.; Seminati, C.; Tello, M.; Redondo, N.; Darwich, L.; Martín, M. Antimicrobial Resistance Profile and Prevalence of Extended-Spectrum Beta-Lactamases (ESBL), AmpC Beta-Lactamases and Colistin Resistance (MCR) Genes in Escherichia Coli from Swine Between 1999 and 2018. Porc. Health Manag. 2020, 6, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Al Atya, A.K.; Abriouel, H.; Kempf, I.; Jouy, E.; Auclair, E.; Vachée, A.; Drider, D. Effects of Colistin and Bacteriocins Combinations on the In Vitro Growth of Escherichia coli Strains from Swine Origin. Probiotics Antimicrob. Proteins 2016, 8, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Lauková, A.; Styková, E.; Kubašová, I.; Gancarčíková, S.; Plachá, I.; Mudroňová, D.; Kandričáková, A.; Miltko, R.; Belzecki, G.; Valocký, I.; et al. Enterocin M and its Beneficial Effects in Horses—A Pilot Experiment. Probiotics Antimicrob. Proteins 2018, 10, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Lauková, A.; Chrastinová, Ľ.; Plachá, I.; Kandričáková, A.; Szabóová, R.; Strompfová, V.; Chrenková, M.; Čobanová, K.; Žitňan, R. Beneficial Effect of Lantibiotic Nisin in Rabbit Husbandry. Probiotics Antimicrob. Proteins 2014, 6, 41–46. [Google Scholar] [CrossRef]
- Simonová, M.P.; Chrastinová, Ľ.; Chrenková, M.; Formelová, Z.; Lauková, A. Lantibiotic Nisin Applied in Broiler Rabbits and Its Effect on the Growth Performance and Carcass Quality. Probiotics Antimicrob. Proteins 2019, 11, 1414–1417. [Google Scholar] [CrossRef] [PubMed]
- Araújo, C.; Muñoz-Atienza, E.; Pérez-Sánchez, T.; Poeta, P.; Igrejas, G.; Hernández, P.E.; Herranz, C.; Ruiz-Zarzuela, I.; Cintas, L.M.; Nisin, Z. Production by Lactococcus lactis subsp. Cremoris WA2-67 of Aquatic Origin as a Defense Mechanism to Protect Rainbow Trout (Oncorhynchus mykiss, Walbaum) Against Lactococcus garvieae. Mar. Biotechnol. 2015, 17, 820–830. [Google Scholar] [CrossRef] [PubMed]
- Baños, A.; Ariza, J.J.; Nuñez, C.; Gil-Martínez, L.; García-López, J.D.; Martínez-Bueno, M.; Valdivia, E. Effects of Enterococcus Faecalis UGRA10 and the Enterocin AS-48 against the Fish Pathogen Lactococcus garvieae. Studies In Vitro and In Vivo. Food Microbiol. 2019, 77, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Sha, Y.; Wang, B.; Liu, M.; Jiang, K.; Wang, L. Interaction between Lactobacillus Pentosus HC-2 and Vibrio Parahaemolyticus E1 in Litopenaeus Vannamei in Vivo and in Vitro. Aquaculture 2016, 465, 117–123. [Google Scholar] [CrossRef]
- Lüders, T.; Birkemo, G.A.; Fimland, G.; Nissen-Meyer, J.; Nes, I.F. Strong Synergy between a Eukaryotic Antimicrobial Peptide and Bacteriocins from Lactic Acid Bacteria. Appl. Environ. Microbiol. 2003, 69, 1797–1799. [Google Scholar] [CrossRef] [PubMed]
- Van Der Linden, D.S.; Short, D.; Dittmann, A.; Yu, P.-L. Synergistic Effects of Ovine-Derived Cathelicidins and Other Antimicrobials against Escherichia coli O157:H7 and Staphylococcus aureus 1056 MRSA. Biotechnol. Lett. 2009, 31, 1265–1267. [Google Scholar] [CrossRef] [PubMed]
- Galvin, M.; Hill, C.; Ross, R.P. Lacticin 3147 Displays Activity in Buffer against Gram-Positive Bacterial Pathogens Which Appear Insensitive in Standard Plate Assays. Lett. Appl. Microbiol. 1999, 28, 355–358. [Google Scholar] [CrossRef] [PubMed]
- McAuliffe, O.; Hill, C.; Ross, R.P. Inhibition of Listeria monocytogenesin Cottage Cheese Manufactured with a Lacticin 3147-Producing Starter Culture. J. Appl. Microbiol. 1999, 86, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Rea, M.C.; Clayton, E.; O’Connor, P.M.; Shanahan, F.; Kiely, B.; Ross, R.P.; Hill, C. Antimicrobial Activity of Lacticin 3147 against Clinical Clostridium Difficile Strains. J. Med Microbiol. 2007, 56, 940–946. [Google Scholar] [CrossRef]
- Piper, C.; Draper, L.A.; Cotter, P.D.; Ross, R.P.; Hill, C. A Comparison of the Activities of Lacticin 3147 and Nisin against Drug-Resistant Staphylococcus aureus and Enterococcus Species. J. Antimicrob. Chemother. 2009, 64, 546–551. [Google Scholar] [CrossRef]
- Carroll, J.; Draper, L.A.; O’Connor, P.M.; Coffey, A.; Hill, C.; Ross, R.P.; Cotter, P.D.; O’Mahony, J. Comparison of the Activities of the Lantibiotics Nisin and Lacticin 3147 against Clinically Significant Mycobacteria. Int. J. Antimicrob. Agents 2010, 36, 132–136. [Google Scholar] [CrossRef]
- Kitching, M.; Mathur, H.; Flynn, J.; Byrne, N.; Dillon, P.; Sayers, R.; Rea, M.C.; Hill, C.; Ross, R.P. A Live Bio-Therapeutic for Mastitis, Containing Lactococcus lactis DPC3147 with Comparable Efficacy to Antibiotic Treatment. Front. Microbiol. 2019, 10, 2220. [Google Scholar] [CrossRef] [PubMed]
- Ryan, M.; Flynn, J.; Hill, C.; Ross, R.; Meaney, W. The Natural Food Grade Inhibitor, Lacticin 3147, Reduced the Incidence of Mastitis After Experimental Challenge with Streptococcus dysgalactiae in Nonlactating Dairy Cows. J. Dairy Sci. 1999, 82, 2625–2631. [Google Scholar] [CrossRef]
- Twomey, D.; Wheelock, A.; Flynn, J.; Meaney, W.; Hill, C.; Ross, R. Protection against Staphylococcus aureus Mastitis in Dairy Cows Using a Bismuth-Based Teat Seal Containing the Bacteriocin, Lacticin 3147. J. Dairy Sci. 2000, 83, 1981–1988. [Google Scholar] [CrossRef]
- Kim, S.; Shin, S.; Koo, H.; Youn, J.-H.; Paik, H.-D.; Park, Y. In Vitro Antimicrobial Effect and in Vivo Preventive and Therapeutic Effects of Partially Purified Lantibiotic Lacticin NK34 against Infection by Staphylococcus Species Isolated from Bovine Mastitis. J. Dairy Sci. 2010, 93, 3610–3615. [Google Scholar] [CrossRef] [PubMed]
- Ogunbanwo, S.; Sanni, A.; Onilude, A. Influence of Bacteriocin in the Control of Escherichia coli Infection of Broiler Chickens in Nigeria. World J. Microbiol. Biotechnol. 2004, 20, 51–56. [Google Scholar] [CrossRef]
- Wang, Q.; Cui, Y.; Wang, W.; Xu, J.; Xu, L. Production of Two Bacteriocins in Various Growth Conditions Produced by Gram-Positive Bacteria Isolated from Chicken Cecum. Can. J. Microbiol. 2012, 58, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Stern, N.J.; Svetoch, E.A.; Eruslanov, B.V.; Kovalev, Y.N.; Volodina, L.I.; Perelygin, V.V.; Mitsevich, E.V.; Mitsevich, I.P.; Levchuk, V.P. Paenibacillus polymyxa Purified Bacteriocin To Control Campylobacter jejuni in Chickens. J. Food Prot. 2005, 68, 1450–1453. [Google Scholar] [CrossRef] [PubMed]
- Daeschel, M.A.; Klaenhammer, T.R. Association of a 13.6-Megadalton Plasmid in Pediococcus pentosaceus with Bacteriocin Activity. Appl. Environ. Microbiol. 1985, 50, 1538–1541. [Google Scholar] [CrossRef] [PubMed]
- Piva, A.; Headon, D.R. Pediocin A, a Bacteriocin Produced by Pediococcus pentosaceus FBB61. Microbiology 1994, 140, 697–702. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Keyburn, A.L.; Boyce, J.D.; Vaz, P.; Bannam, T.L.; Ford, M.E.; Parker, D.; Di Rubbo, A.; Rood, J.I.; Moore, R.J. NetB, a New Toxin That Is Associated with Avian Necrotic Enteritis Caused by Clostridium perfringens. PLoS Pathog. 2008, 4, e26. [Google Scholar] [CrossRef]
- Grilli, E.; Messina, M.R.; Catelli, E.; Morlacchini, M.; Piva, A. Pediocin A Improves Growth Performance of Broilers Challenged with Clostridium perfringens. Poult. Sci. 2009, 88, 2152–2158. [Google Scholar] [CrossRef]
- Józefiak, D.; Sip, A.; Rawski, M.; Rutkowski, A.; Kaczmarek, S.; Hojberg, O.; Jensen, B.; Engberg, R. Dietary Divercin Modifies Gastrointestinal Microbiota and Improves Growth Performance in Broiler Chickens. Br. Poult. Sci. 2011, 52, 492–499. [Google Scholar] [CrossRef] [PubMed]
- Józefiak, D.; Kierończyk, B.; Juśkiewicz, J.; Zduńczyk, Z.; Rawski, M.; Długosz, J.; Sip, A.; Højberg, O. Dietary Nisin Modulates the Gastrointestinal Microbial Ecology and Enhances Growth Performance of the Broiler Chickens. PLoS ONE 2013, 8, e85347. [Google Scholar] [CrossRef] [PubMed]
- Kierończyk, B.; Rawski, M.; Mikołajczak, Z.; Świątkiewicz, S.; Józefiak, D. Nisin as a Novel Feed Additive: The Effects on Gut Microbial Modulation and Activity, Histological Parameters, and Growth Performance of Broiler Chickens. Animals 2020, 10, 101. [Google Scholar] [CrossRef] [PubMed]
- Carrasco, J.M.D.; Casanova, N.A.; Miyakawa, M.E.F. Microbiota, Gut Health and Chicken Productivity: What Is the Connection? Microorganisms 2019, 7, 374. [Google Scholar] [CrossRef] [PubMed]
- Line, J.E.; Svetoch, E.A.; Eruslanov, B.V.; Perelygin, V.V.; Mitsevich, E.V.; Mitsevich, I.P.; Levchuk, V.P.; Svetoch, O.E.; Seal, B.S.; Siragusa, G.R.; et al. Isolation and Purification of Enterocin E-760 with Broad Antimicrobial Activity against Gram-Positive and Gram-Negative Bacteria. Antimicrob. Agents Chemother. 2007, 52, 1094–1100. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, R.; Todorov, S.D.; Leon, M.T.D. Mode of action and In Vitro Susceptibility of Mastitis Pathogens to Macedocin ST91KM and preparation of a teat seal containing the bacteriocin. Braz. J. Microbiol. 2010, 41, 133–145. [Google Scholar] [CrossRef]
- Dicks, L.M.T.; Dreyer, L.; Smith, C.; Van Staden, A.D. A Review: The Fate of Bacteriocins in the Human Gastro-Intestinal Tract: Do They Cross the Gut–Blood Barrier? Front. Microbiol. 2018, 9, 2297. [Google Scholar] [CrossRef] [PubMed]
- Stern, N.J.; Svetoch, E.A.; Eruslanov, B.V.; Perelygin, V.V.; Mitsevich, E.V.; Mitsevich, I.P.; Pokhilenko, V.D.; Levchuk, V.P.; Svetoch, O.E.; Seal, B.S. Isolation of a Lactobacillus salivarius Strain and Purification of Its Bacteriocin, Which Is Inhibitory to Campylobacter jejuni in the Chicken Gastrointestinal System. Antimicrob. Agents Chemother. 2006, 50, 3111–3116. [Google Scholar] [CrossRef]
- Umu, Ö.C.; Bäuerl, C.; Oostindjer, M.; Pope, P.B.; Hernández, P.E.; Pérez-Martínez, G.; Diep, D.B. The Potential of Class II Bacteriocins to Modify Gut Microbiota to Improve Host Health. PLoS ONE 2016, 11, e0164036. [Google Scholar] [CrossRef] [PubMed]
- Chaucheyras-Durand, F.; Durand, H. Probiotics in Animal Nutrition and Health. Benef. Microbes 2010, 1, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Sun, K.; Wu, Y.; Yang, Y.; Tso, P.; Wu, Z. Interactions between Intestinal Microbiota and Host Immune Response in Inflammatory Bowel Disease. Front. Immunol. 2017, 8, 942. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Gilbert, J. Microbial Exposure and Human Health. Curr. Opin. Microbiol. 2018, 44, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Kubašová, I.; Strompfová, V.; Lauková, A. Safety Assessment of Commensal Enterococci from Dogs. Folia Microbiol. 2017, 62, 491–498. [Google Scholar] [CrossRef]
- Strompfová, V.; Kubašová, I.; Ščerbová, J.; Maďari, A.; Gancarčíková, S.; Mudroňová, D.; Miltko, R.; Belzecki, G.; Lauková, A. Oral Administration of Bacteriocin-Producing and Non-producing Strains of Enterococcus faecium in Dogs. Appl. Microbiol. Biotechnol. 2019, 103, 4953–4965. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Deng, J.; Li, Y.; Yang, Q. The effect of Lactobacillus on the Expression of Porcine β-Defensin-2 in the Digestive Tract of Piglets. Livest. Sci. 2011, 138, 259–265. [Google Scholar] [CrossRef]
- Messaoudi, S.; Manai, M.; Kergourlay, G.; Prévost, H.; Connil, N.; Chobert, J.-M.; Dousset, X. Lactobacillus salivarius: Bacteriocin and Probiotic Activity. Food Microbiol. 2013, 36, 296–304. [Google Scholar] [CrossRef]
- Corr, S.C.; Li, Y.; Riedel, C.U.; O’Toole, P.W.; Hill, C.; Gahan, C.G.M. Bacteriocin Production as a Mechanism for the Antiinfective Activity of Lactobacillus salivarius UCC118. Proc. Natl. Acad. Sci. USA 2007, 104, 7617–7621. [Google Scholar] [CrossRef] [PubMed]
- Claesson, M.J.; Li, Y.; Leahy, S.; Canchaya, C.; Van Pijkeren, J.P.; Cerdeño-Tárraga, A.M.; Parkhill, J.; Flynn, S.; O’Sullivan, G.C.; Collins, J.K.; et al. Multireplicon Genome Architecture of Lactobacillus salivarius. Proc. Natl. Acad. Sci. USA 2006, 103, 6718–6723. [Google Scholar] [CrossRef]
- Riboulet-Bisson, E.; Sturme, M.H.J.; Jeffery, I.B.; O’Donnell, M.M.; Neville, B.A.; Forde, B.M.; Claesson, M.J.; Harris, H.; Gardiner, G.E.; Casey, P.G.; et al. Effect of Lactobacillus salivarius Bacteriocin Abp118 on the Mouse and Pig Intestinal Microbiota. PLoS ONE 2012, 7, e31113. [Google Scholar] [CrossRef]
- Hu, J.; Ma, L.; Nie, Y.; Chen, J.; Zheng, W.; Wang, X.; Xie, C.; Zheng, Z.; Wang, Z.; Yang, T.; et al. A Microbiota-Derived Bacteriocin Targets the Host to Confer Diarrhea Resistance in Early-Weaned Piglets. Cell Host Microbe 2018, 24, 817–832.e8. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, A.B.; Back, M.; Daguer, H.; Palmeira, M.; Ploêncio, L.A.D.S.; Molognoni, L.; Peripolli, V.; Bianchi, I. Carry-over and Contamination of Veterinary Drugs in Feed Production Lines for Poultry and Pigs. Food Addit. Contam. Part A 2019, 36, 740–751. [Google Scholar] [CrossRef] [PubMed]
- Kizerwetter-Swida, M.; Binek, M. Selection of Potentially Probiotic Lactobacillus Strains towards Their Inhibitory Activity against Poultry Enteropathogenic bacteria. Pol. J. Microbiol. 2005, 54, 287–294. [Google Scholar]
- Timmerman, H.M.; Veldman, A.; Elsen, E.V.D.; Rombouts, F.M.; Beynen, A.C. Mortality and Growth Performance of Broilers Given Drinking Water Supplemented with Chicken-Specific Probiotics. Poult. Sci. 2006, 85, 1383–1388. [Google Scholar] [CrossRef] [PubMed]
- Giannenas, I.; Tsalie, E.; Triantafillou, E.; Hessenberger, S.; Teichmann, K.; Mohnl, M.; Tontis, D. Assessment of Probiotics Supplementation via Feed or Water on the Growth Performance, Intestinal Morphology and Microflora of Chickens after Experimental Infection Witheimeria acervulina, Eimeria maxima andeimeria tenella. Avian Pathol. 2014, 43, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Ritzi, M.M.; Abdelrahman, W.; Van-Heerden, K.; Mohnl, M.; Barrett, N.W.; Dalloul, R.A. Combination of Probiotics and Coccidiosis Vaccine Enhances Protection against an Eimeria Challenge. Vet. Res. 2016, 47, 1–8. [Google Scholar] [CrossRef]
- Pender, C.; Kim, S.; Potter, T.; Ritzi, M.; Young, M.; Dalloul, R. Effects of in Ovo Supplementation of Probiotics on Performance and Immunocompetence of Broiler Chicks to an Eimeria Challenge. Benef. Microbes 2016, 7, 699–705. [Google Scholar] [CrossRef] [PubMed]
- Shin, M.; Han, S.; Ji, A.; Kim, K.; Lee, W. Isolation and Characterization of Bacteriocin-Producing Bacteria from the Gastrointestinal Tract of Broiler Chickens for Probiotic Use. J. Appl. Microbiol. 2008, 105, 2203–2212. [Google Scholar] [CrossRef]
- Sabo, S.D.S.; Mendes, M.A.; Araújo, E.D.S.; Muradian, L.B.D.A.; Makiyama, E.N.; Leblanc, J.G.; Borelli, P.; Fock, R.A.; Knöbl, T.; Oliveira, R.P.D.S. Bioprospecting of Probiotics with Antimicrobial Activities against Salmonella Heidelberg and That Produce B-Complex Vitamins as Potential Supplements in Poultry Nutrition. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef]
- Hansen, J.N.; Sandine, W.E. Nisin as a Model Food Preservative. Crit. Rev. Food Sci. Nutr. 1994, 34, 69–93. [Google Scholar] [CrossRef] [PubMed]
- Pablo, M.A.; Gaforio, J.J.; Gallego, A.M.; Ortega, E.; Gã¡lvez, A.M.; Lã³Pez, G.A.D.C. Evaluation of Immunomodulatory Effects of Nisin-Containing Diets on Mice. FEMS Immunol. Med Microbiol. 1999, 24, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Małaczewska, J.; Kaczorek-Łukowska, E.; Wójcik, R.; Rękawek, W.; Siwicki, A.K. In Vitro Immunomodulatory Effect of Nisin on Porcine Leucocytes. J. Anim. Physiol. Anim. Nutr. 2019, 103, 882–893. [Google Scholar] [CrossRef] [PubMed]
- Aranha, C.; Gupta, S.; Reddy, K. Assessment of Cervicovaginal Cytokine Levels Following Exposure to Microbicide Nisin Gel in Rabbits. Cytokine 2008, 43, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.M.; Aranha, C.C.; Mohanty, M.C.; Reddy, K.V.R. Toll-Like Receptors and Cytokines as Surrogate Biomarkers for Evaluating Vaginal Immune Response following Microbicide Administration. Mediat. Inflamm. 2008, 2008, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Begde, D.; Bundale, S.; Mashitha, P.; Rudra, J.; Nashikkar, N.; Upadhyay, A. Immunomodulatory Efficacy of Nisin-A Bacterial Lantibiotic Peptide. J. Pept. Sci. 2011, 17, 438–444. [Google Scholar] [CrossRef]
- Kindrachuk, J.; Jenssen, H.; Elliott, M.; Nijnik, A.; Magrangeas-Janot, L.; Pasupuleti, M.; Thorson, L.; Ma, S.; Easton, D.M.; Bains, M.; et al. Manipulation of Innate Immunity by a Bacterial Secreted Peptide: Lantibiotic Nisin Z Is Selectively Immunomodulatory. Innate Immun. 2012, 19, 315–327. [Google Scholar] [CrossRef]
- Villamil, L.; Figueras, A.; Novoa, B. Immunomodulatory Effects of Nisin in Turbot (Scophthalmus maximus L.). Fish Shellfish. Immunol. 2003, 14, 157–169. [Google Scholar] [CrossRef] [PubMed]
- Malvisi, M.; Stuknytė, M.; Magro, G.; Minozzi, G.; Giardini, A.; De Noni, I.; Piccinini, R. Antibacterial Activity and Immunomodulatory Effects on a Bovine Mammary Epithelial Cell Line Exerted by Nisin A-Producing Lactococcus Lactis Strains. J. Dairy Sci. 2016, 99, 2288–2296. [Google Scholar] [CrossRef]
- Jia, Z.; He, M.; Wang, C.; Chen, A.; Zhang, X.; Xu, J.; Fu, H.; Liu, B. Nisin Reduces Uterine Inflammation in Rats by Modulating Concentrations of Pro- and Anti-Inflammatory Cytokines. Am. J. Reprod. Immunol. 2019, 81, e13096. [Google Scholar] [CrossRef]
- Fu, T.; Yu, M.; Yan, Q.; Liu, Y.-M. Bacteriocin Isolated from Lactobacillus Rhamnosus L34 Has Antibacterial Effects in a Rabbit Model of Infection After Mandible Fracture Fixation. Med Sci. Monit. 2018, 24, 8009–8014. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Zhang, D. Antibacterial Effects of Bacteriocins Isolated from Lactobacillus rhamnosus (ATCC 53103) In a Rabbit Model of Knee Implant Infection. Exp. Ther. Med. 2018, 15, 2985–2989. [Google Scholar] [CrossRef] [PubMed]
- Gong, L.; He, H.; Li, D.; Cao, L.; Khan, T.A.; Li, Y.; Pan, L.; Yan, L.; Ding, X.; Sun, Y.; et al. A New Isolate of Pediococcus pentosaceus (SL001) with Antibacterial Activity against Fish Pathogens and Potency in Facilitating the Immunity and Growth Performance of Grass Carps. Front. Microbiol. 2019, 10, 1384. [Google Scholar] [CrossRef] [PubMed]
- Li, M.-F.; Zhang, B.-C.; Li, J.; Sun, L. Sil: A Streptococcus iniae Bacteriocin with Dual Role as an Antimicrobial and an Immunomodulator That Inhibits Innate Immune Response and Promotes, S. iniae Infection. PLoS ONE 2014, 9, e96222. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Class of Bacteriocin | Subclasses | Molecular Properties | Reference |
|---|---|---|---|
| Class I Lantibiotic | Ia Lanthipeptides Ib Globular and inflexible bacteriocins Ic Sactipeptides | Small, heat-stable bacteriocins (<5 kDa), have a post-translational modification, resulting in the formation of atypical amino acids lanthionine and methyllanthionine. | [33,36,37,38,39] |
| Class II Non-lantibiotic | IIa Pediocin-like IIb Two peptides IIc Leader less IId Non pediocin-like Single-peptide | Small and flexible bacteriocins (<10 kDa), with an amphiphilic helical structure. These peptides do not contain modified amino acid residues and are pH and heat-resistant. | [13,40,41,42,43,44,45,46] |
| Class III | IIIa Bacteriolysins IIIb Nonlytic | High molecular weight bacteriocins (>30 kDa), thermolabile and unmodified peptides. | [47,48] |
| Bacteriocin | LAB Producer of Bacteriocin | Susceptible Bacteria | Reference |
|---|---|---|---|
| Enterocin AS-48 | Enterococcus faecalis UGRA10 | Lactococcus garvieae | [100] |
| Enterocin M | Enterococcus faecium AL41 | Campylobacter spp. Clostridium spp. | [96] |
| Enterocin CLE34 | Enterococcus faecium CLE34 | Salmonella pullorum | [114] |
| Enterocin E-760 | Enterococcus durans Enterococcus faecium Enterococcus hirae | Salmonella enterica serovar Enteritidis S. enterica serovar Choleraesuis S. enterica serovar Typhimurium S. enterica serovar Gallinarum Escherichia coli O157:H7 Yersinia enterocolitica Staphylococcus aureus Campylobacter jejuni | [124] |
| Lacticin 3147 | Lactococcus lactis DPC3147. | Streptococcus dysgalactiae, Streptococcus agalactiae Staphylococcus aureus Streptococcus uberis | [110] |
| Mycobacterium avium subsp. paratuberculosis | [108] | ||
| Macedocin ST91KM | Streptococcus gallolyticus subsp. macedonicus ST91KM | Streptococcus agalactiae Streptococcus dysgalactiae Streptococcus uberis Staphylococcus aureus | [125] |
| Nisin A | Lactococcus lactic subsp. lactis | Enterococcus faecalis ssp. liquefaciens Streptococcus equinus Staphylococcus epidermidis Staphylococcus aureus Streptococcus uberis Streptococcus dysgalactiae Streptococcus agalactiae | [77] |
| Streptococcus suis Mycobacterium avium subsp. paratuberculosis | [85] [108] | ||
| Nisin A Nisin V | L. lactis NZ9700 L. lactis NZ9800nisA:M21V | Listeria monocytogenes | [87] |
| Nisin Z | L. lactis NIZO22186 | Staphylococcus aureus Streptococcus agalactiae | [78] |
| Pediocin A | Pediococcus pentosaceus FBB61 | Clostridium perfringens | [119] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernández-González, J.C.; Martínez-Tapia, A.; Lazcano-Hernández, G.; García-Pérez, B.E.; Castrejón-Jiménez, N.S. Bacteriocins from Lactic Acid Bacteria. A Powerful Alternative as Antimicrobials, Probiotics, and Immunomodulators in Veterinary Medicine. Animals 2021, 11, 979. https://doi.org/10.3390/ani11040979
Hernández-González JC, Martínez-Tapia A, Lazcano-Hernández G, García-Pérez BE, Castrejón-Jiménez NS. Bacteriocins from Lactic Acid Bacteria. A Powerful Alternative as Antimicrobials, Probiotics, and Immunomodulators in Veterinary Medicine. Animals. 2021; 11(4):979. https://doi.org/10.3390/ani11040979
Chicago/Turabian StyleHernández-González, Juan Carlos, Abigail Martínez-Tapia, Gebim Lazcano-Hernández, Blanca Estela García-Pérez, and Nayeli Shantal Castrejón-Jiménez. 2021. "Bacteriocins from Lactic Acid Bacteria. A Powerful Alternative as Antimicrobials, Probiotics, and Immunomodulators in Veterinary Medicine" Animals 11, no. 4: 979. https://doi.org/10.3390/ani11040979
APA StyleHernández-González, J. C., Martínez-Tapia, A., Lazcano-Hernández, G., García-Pérez, B. E., & Castrejón-Jiménez, N. S. (2021). Bacteriocins from Lactic Acid Bacteria. A Powerful Alternative as Antimicrobials, Probiotics, and Immunomodulators in Veterinary Medicine. Animals, 11(4), 979. https://doi.org/10.3390/ani11040979