
Towards a Sustainable Reproduction Management of Dairy Sheep: Glycerol-Based Formulations as Alternative to eCG in Milked Ewes Mated at the End of Anoestrus Period

 , ,
, ,  and
and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
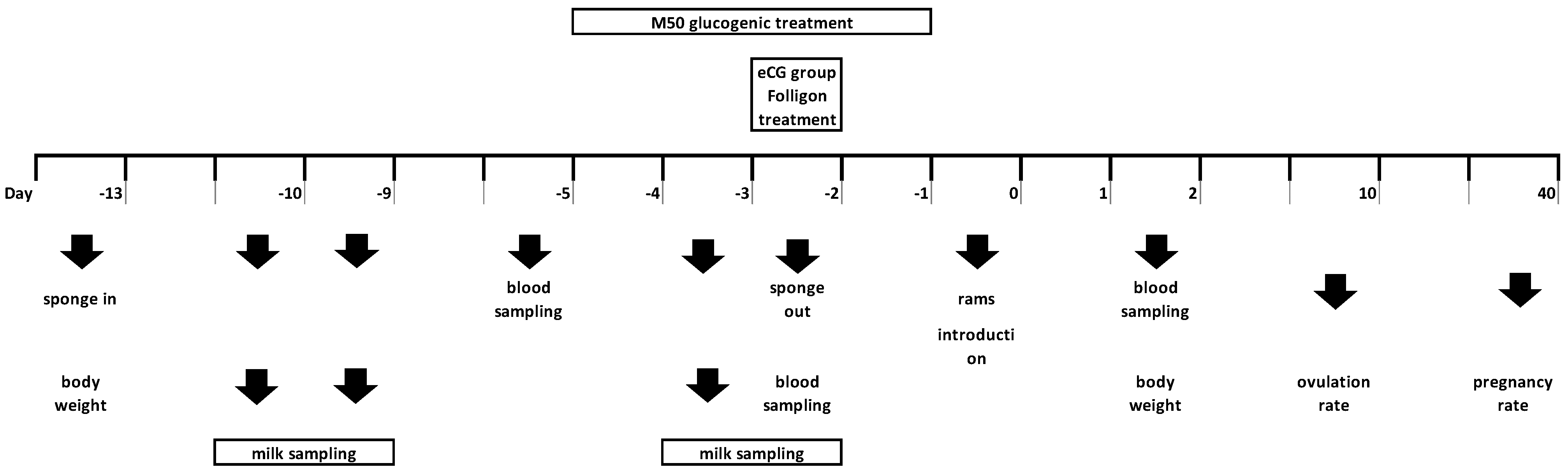
2.1. Animals and Treatments
2.2. Feedstuff Composition
2.3. Blood Samplings
2.4. Plasma Osmolality Determination
2.5. Metabolites
2.6. Insulin
2.7. Intake, Body Weight, Body Condition Score, Milk Yield and Milk Composition
2.8. Ultrasound Scanning
2.9. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scaramuzzi, R.J.; Campbell, B.K.; Downing, J.A.; Kendall, N.R.; Khalid, M.; Munoz-Gutierrez, M.; Somchit, A.; Muñoz-Gutiérrez, M.; Somchit, A. A review of the effects of supplementary nutrition in the ewe on the concentrations of reproductive and metabolic hormones and the mechanisms that regulate folliculogenesis and ovulation rate. Reprod. Nutr. Dev. 2006, 46, 339–354. [Google Scholar] [CrossRef]
- Bodin, L.; Drion, P.V.; Remy, B.; Brice, G.; Cognié, Y.; Beckers, J.F. Anti-PMSG antibody levels in sheep subjected annually to oestrus synchronisation. Reprod. Nutr. Dev. 1997, 37, 651–660. [Google Scholar] [CrossRef]
- Porcu, C.; Manca, C.; Cabiddu, A.; Dattena, M.; Gallus, M.; Pasciu, V.; Succu, S.; Naitana, S.; Berlinguer, F.; Molle, G. Effects of short-term administration of a glucogenic mixture at mating on feed intake, metabolism, milk yield and reproductive performance of lactating dairy ewes. Anim. Feed Sci. Technol. 2018, 243, 10–21. [Google Scholar] [CrossRef]
- Porcu, C.; Sotgiu, F.D.; Pasciu, V.; Cappai, M.G.; Barbero-Fernández, A.; Gonzalez-Bulnes, A.; Dattena, M.; Gallus, M.; Molle, G.; Berlinguer, F. Administration of glycerol-based formulations in sheep results in similar ovulation rate to eCG but red blood cell indices may be affected. BMC Vet. Res. 2020, 16, 207. [Google Scholar] [CrossRef] [PubMed]
- Berlinguer, F.; Gonzalez-Bulnes, A.; Contreras-Solis, I.; Spezzigu, A.; Torres-Rovira, L.; Succu, S.; Naitana, S.; Leoni, G.G. Glucogenic supply increases oocyte developmental competence in sheep. Reprod. Fertil. Dev. 2012, 24, 1055–1062. [Google Scholar] [CrossRef] [PubMed]
- Sutton-McDowall, M.L.; Gilchrist, R.B.; Thompson, J.G. The pivotal role of glucose metabolism in determining oocyte developmental competence. Reproduction 2010, 139, 685. [Google Scholar] [CrossRef]
- Gutierrez, C.G.; Ferraro, S.; Martinez, V.; Saharrea, A.; Cortez, C.; Lassala, A.; Basurto, H.; Hernandez, J. Increasing ovulation quota: More than a matter of energy. Acta Sci. Vet. 2011, 39, 305–316. [Google Scholar]
- Iglesias, R.M.R.; Ciccioli, N.H.; Irazoqui, H.; Giglioli, C. Ovulation rate in ewes after single oral glucogenic dosage during a ram-induced follicular phase. Anim. Reprod. Sci. 1996, 44, 211–221. [Google Scholar] [CrossRef]
- Letelier, C.; Mallo, F.; Encinas, T.; Ros, J.M.; Gonzalez-Bulnes, A. Glucogenic supply increases ovulation rate by modifying follicle recruitment and subsequent development of preovulatory follicles without effects on ghrelin secretion. Reproduction 2008, 136, 65–72. [Google Scholar] [CrossRef]
- Porcu, C.; Pasciu, V.; Succu, S.; Baralla, E.; Manca, M.E.E.; Serra, E.; Leoni, G.G.G.; Dattena, M.; Bomboi, G.C.C.; Molle, G.; et al. Glucogenic treatment creates an optimal metabolic milieu for the conception period in ewes. Domest. Anim. Endocrinol. 2017, 59, A7–A12. [Google Scholar] [CrossRef]
- Diskin, M.G.; Mackey, D.R.; Roche, J.F.; Sreenan, J.M. Effects of nutrition and metabolic status on circulating hormones and ovarian follicle development in cattle. Anim. Reprod. Sci. 2003, 78, 345–370. [Google Scholar] [CrossRef]
- Gunn, P.J.; Neary, M.K.; Lemenager, R.P.; Lake, S.L. Effects of crude glycerin on performance and carcass characteristics of finishing wether lambs. J. Anim. Sci. 2010. [Google Scholar] [CrossRef] [PubMed]
- Novais-Eiras, D.; de Carvalho, G.G.P.; Leite, L.C.; Eiras, C.E.; de Freitas, J.E.; Pina, D.D.S.; Ferreira, F.G.; dos Santos, G.T.; Grande, P.A. Crude glycerin in the feed supplementation of lactating goats on pasture. Small Rumin. Res. 2018, 168, 39–46. [Google Scholar] [CrossRef]
- Dupont, J.; Scaramuzzi, R.J.; Reverchon, M. The effect of nutrition and metabolic status on the development of follicles, oocytes and embryos in ruminants. Animal 2014, 8, 1031–1044. [Google Scholar] [CrossRef]
- Elrod, C.C.; Butler, W.R. Reduction of fertility and alteration of uterine pH in heifers fed excess ruminally degradable protein. J. Anim. Sci. 1993, 71, 694–701. [Google Scholar] [CrossRef]
- Dupont, J.; Reverchon, M.; Bertoldo, M.J.; Froment, P. Nutritional signals and reproduction. Mol. Cell. Endocrinol. 2014, 382, 527–537. [Google Scholar] [CrossRef]
- McEvoy, T.G.; Robinson, J.J.; Aitken, R.P.; Findlay, P.A.; Robertson, I.S. Dietary excesses of urea influence the viability and metabolism of preimplantation sheep embryos and may affect fetal growth among survivors. Anim. Reprod. Sci. 1997, 47, 71–90. [Google Scholar] [CrossRef]
- Leroy, J.L.M.R.; Vanholder, T.; Mateusen, B.; Christophe, A.; Opsomer, G.; de Kruif, A.; Genicot, G.; Van Soom, A. Non-esterified fatty acids in follicular fluid of dairy cows and their effect on developmental capacity of bovine oocytes in vitro. Reproduction 2005, 130, 485–495. [Google Scholar] [CrossRef]
- Leroy, J.L.M.R.; Vanholder, T.; Delanghe, J.R.; Opsomer, G.; Van Soom, A.; Bols, P.E.J.; De Kruif, A. Metabolite and ionic composition of follicular fluid from different-sized follicles and their relationship to serum concentrations in dairy cows. Anim. Reprod. Sci. 2004, 80, 201–211. [Google Scholar] [CrossRef]
- Aardema, H.; Vos, P.L.A.M.; Lolicato, F.; Roelen, B.A.J.; Knijn, H.M.; Vaandrager, A.B.; Helms, B.J.; Gadella, B.M. Oleic acid prevents detrimental effects of saturated fatty acids on bovine oocyte developmental competence. Biol. Reprod. 2011, 85, 62–69. [Google Scholar] [CrossRef]
- Ying, S.J.; Xiao, S.H.; Wang, C.L.; Zhong, B.S.; Zhang, G.M.; Wang, Z.Y.; He, D.Y.; Ding, X.L.; Xing, H.J.; Wang, F. Effect of nutrition on plasma lipid profile and mRNA levels of ovarian genes involved in steroid hormone synthesis in Hu sheep during luteal phase. J. Anim. Sci. 2013, 91, 5229–5239. [Google Scholar] [CrossRef]
- The association of official analytical chemists. Official methods of analysis. Anal. Chim. Acta 1990, 231, 1–347. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Aufrère, J.; Demarquilly, C. Predicting organic matter digestibility of forage by two pepsin-cellulase methods. In Proceedings of the 16th International Grassland Congress, Nice, France, 4–11 October 1989; pp. 877–878. [Google Scholar]
- Cannas, A. Feeding of lactating ewe. In Dairy Sheep Nutrition; Pulina, G., Ed.; CAB International: Wallingford, UK, 2004; pp. 31–49. [Google Scholar]
- Russel, A.J.F.; Doney, J.M.; Gunn, R.G. Subjective assessment of body fat in live sheep. J. Agric. Sci. 1969, 72, 451–454. [Google Scholar] [CrossRef]
- Santos-jimenez, Z.; Guillen-gargallo, S.; Encinas, T.; Berlinguer, F.; Veliz-deras, F.G.; Martinez-ros, P.; Gonzalez-bulnes, A. Use of propylene-glycol as a cosolvent for GnRH in synchronization of estrus and ovulation in sheep. Animals 2020, 10, 897. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Ros, P.; Gonzalez-Bulnes, A. Efficiency of cidr-based protocols including gnrh instead of ecg for estrus synchronization in sheep. Animals 2019, 9, 146. [Google Scholar] [CrossRef]
- Manteca Vilanova, X.; De Briyne, N.; Beaver, B.; Turner, P. V Horse Welfare During Equine Chorionic Gonadotropin (eCG) Production. Animals 2019, 9, 1053. [Google Scholar] [CrossRef]
- Martin, G.B.; Kadokawa, H. “Clean, green and ethical” animal production. Case study: Reproductive efficiency in small ruminants. J. Reprod. Dev. 2006, 52, 145–152. [Google Scholar] [CrossRef]
- Quintero-Elisea, J.A.; Macías-Cruz, U.; Álvarez-Valenzuela, F.D.; Correa-Calderón, A.; González-Reyna, A.; Lucero-Magaña, F.A.; Soto-Navarro, S.A.; Avendaño-Reyes, L. The effects of time and dose of pregnant mare serum gonadotropin (PMSG) on reproductive efficiency in hair sheep ewes. Trop. Anim. Health Prod. 2011, 43, 1567–1573. [Google Scholar] [CrossRef]
- Marai, I.F.M.; Habeeb, A.A.M.; Gad, A.E. Reproductive traits of female rabbits as affected by heat stress and lighting regime under subtropical conditions of Egypt. Anim. Sci. 2004, 78, 119–127. [Google Scholar] [CrossRef]
- Manca, C.; Porcu, C.; Giovanetti, V. O161 Effect of a flushing treatment based on a glucogenic mixture on sheep thermal imaging of body temperature and fertility O162 Ovine foot-rot: A welfare and economic concern. Ital. J. Anim. Sci. 2017, 16, 2017. [Google Scholar]
- Bizjak, D.A.; Jungen, P.; Bloch, W.; Grau, M. Cryopreservation of red blood cells: Effect on rheologic properties and associated metabolic and nitric oxide related parameters. Cryobiology 2018, 84, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, E.K.; Lambert, I.H.; Pedersen, S.F. Physiology of cell volume regulation in vertebrates. Physiol. Rev. 2009, 89, 193–277. [Google Scholar] [CrossRef]
- Kholif, A.E. Glycerol use in dairy diets: A systemic review. Anim. Nutr. 2019, 5, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Drackley, J.K.; Cardoso, F.C. Prepartum and postpartum nutritional management to optimize fertility in high-yielding dairy cows in confined TMR systems. Animal 2014, 8, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Campbell, P.J.; Hill, J.; Peter, J.; Carlson, M.G.; Hill, J. Regulation of free fatty acid metabolism by insulin in humans: Role of lipolysis and reesterification. Am. J. Physiol. Endocrinol. Metab. 1992, 263, E1063–E1069. [Google Scholar] [CrossRef]
- GOLAY, A.; CHEN, Y.-D.I.; REAVEN, G.M. Effect of Differences in Glucose Tolerance on Insulin’s Ability to Regulate Carbohydrate and Free Fatty Acid Metabolism in Obese Individuals*. J. Clin. Endocrinol. Metab. 1986, 62, 1081–1088. [Google Scholar] [CrossRef]
- Bortz, W.M.; Paul, P.; Haff, A.C.; Holmes, W.L. Glycerol Turnover and Oxidation in Man. J. Clin. Investig. 1972, 51, 1537–1546. [Google Scholar] [CrossRef]
- Robergs, R.A.; Griffin, S.E. Glycerol. Biochemistry, Pharmacokinetics and Clinical and Practical Applications. Sports Med. 1998, 26, 145–167. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, I. Effects of dietary glycerol on the serum glyceride level of men and women. Br. J. Nutr. 1970, 24, 537–543. [Google Scholar] [CrossRef]
- Naismith, D.J.; Karikari, B.P. The relevance of glycerol metabolism to the hyperlipidaemic property of sucrose. Proc. Nutr. Soc. 1973, 32, 96A–97A. [Google Scholar] [PubMed]
- de Andrade, G.P.; de Carvalho, F.F.R.; Batista, Â.M.V.; Pessoa, R.A.S.; da Costa, C.A.; Cardoso, D.B.; Maciel, M.d.V. Evaluation of crude glycerin as a partial substitute of corn grain in growing diets for lambs. Small Rumin. Res. 2018, 165, 41–47. [Google Scholar] [CrossRef]
- Ribeiro, R.D.X.; Carvalho, G.G.P.; Silva, T.M.; Costa, J.B.; Bezerra, L.R.; Cambuí, G.B.; Barbosa, A.M.; Oliveira, R.L. Effects of crude glycerin from biodiesel on the diets of lambs: Intake, digestibility, performance, feeding behavior, and serum metabolites. J. Anim. Sci. 2018, 96, 1952–1961. [Google Scholar] [CrossRef]
- Mazur, A.; Ozgo, M.; Rayssiguier, Y. Altered plasma triglyceride-rich lipoproteins and triglyceride secretion in feed-restricted pregnant ewes. Vet. Med. (Praha) 2009, 54, 412–418. [Google Scholar] [CrossRef]
- Grummer, R.R.; Carroll, D.J. A Review of Lipoprotein Cholesterol Metabolism: Importance to Ovarian Function. J. Anim. Sci. 1988, 66, 3160–3173. [Google Scholar] [CrossRef] [PubMed]
- Larson, S.F.; Butler, W.R.; Currie, W.B. Reduced Fertility Associated with Low Progesterone Postbreeding and Increased Milk Urea Nitrogen in Lactating Cows. J. Dairy Sci. 1997, 80, 1288–1295. [Google Scholar] [CrossRef]
- Branca, A.; Molle, G.; Sitzia, M.; Decandia, M.; Landau, S. Short-term dietary effects on reproductive wastage after induced ovulation and artificial insemination in primiparous lactating Sarda ewes. Anim. Reprod. Sci. 2000, 58, 59–71. [Google Scholar] [CrossRef]
- Thoh, D.; Pakdeechanuan, P.; Chanjula, P. Effect of supplementary glycerin on milk composition and heat stability in dairy goats. Asian Australas. J. Anim. Sci. 2017, 30, 1711–1717. [Google Scholar] [CrossRef]
- De Lima, A.M.; Rodrigo, G.; Germano, R.; Ribeiro, N.L.; Mesquita, E.; Filho, B.; De Sousa, S.; Justino, S.; Gonçalves, D. Physical-chemical and microbiological quality of milk and cheese of goats fed with bidestilated glycerin. 2020, 2061. ahead of print. [Google Scholar]


{kind=link}
| Concentrate | Grass Hay | Dehydrated Lucerne Hay | ||||
|---|---|---|---|---|---|---|
| Mean | s.d. | Mean | s.d. | Mean | s.d. | |
| DM | 87.94 | 0.65 | 87.54 | 0.23 | 89.95 | 0.49 |
| Ash | 8.66 | 1.16 | 13.50 | 7.43 | 10.28 | 0.30 |
| EE | 3.36 | 0.12 | 1.42 | 0.25 | 1.85 | 0.01 |
| CP | 15.43 | 1.29 | 7.49 | 1.65 | 15.04 | 0.03 |
| NDF | 23.68 | 3.39 | 61.95 | 12.26 | 49.53 | 0.30 |
| ADF | 11.41 | 1.99 | 39.81 | 9.53 | 36.47 | 0.22 |
| ADL | 2.25 | 0.41 | 6.45 | 0.35 | 8.11 | 0.21 |
| Starch | 36.65 | 6.33 | nd | nd | ||
| IVDMD | 85.10 | 2.36 | 45.37 | 9.84 | 59.68 | 1.98 |
| Treatment | Ovulation Rate | Pregnancy Rate | ||||
|---|---|---|---|---|---|---|
| Mean | SE | Mean | SE | |||
| GLU | 1.52 | ± | 0.15 | 0.52 | ± | 0.11 |
| GON | 1.33 | ± | 0.12 | 0.57 | ± | 0.11 |
| Treatment | p Values | |||||
|---|---|---|---|---|---|---|
| GLU | GON | Groups | Time | Groups*Time | ||
| Osmolality (Osm/kg) | D−2 T0 h | 0.301 ± 0.002 a | 0.307 ± 0.002 | 0.221 | 0 | 0.001 |
| D−2 T4 h | 0.324 ± 0.003 b* | 0.312 ± 0.002 * | ||||
| Glycerol (mg/mL) | D−2 T0 h | 0.578 ± 0.009 a | 0.511 ± 0.018 | 0 | 0 | 0 |
| D−2 T4 h | 23.520 ± 5.120 b* | 0.573 ± 0.009 * | ||||
| Glucose (mg/dL) | D−2 T0 h | 50.504 ± 0.692 a | 50.550 ± 1.050 | 0 | 0 | 0 |
| D−2 T4 h | 84.810 ± 4.170 b* | 51.900 ± 1.370 * | ||||
| Insulin (µg/L) | D−2 T0 h | 0.529 ± 0.126 a* | 0.128 ± 0.023 * | 0.048 | 0.004 | 0 |
| D−2 T4 h | 1.001 ± 0.121 b* | 0.219 ± 0.021 * | ||||
| NEFA (mmol/L) | D−2 T0 h | 0.112 ± 0.005 * | 0.147 ± 0.009 A* | 0.03 | 0.003 | 0.014 |
| D−2 T4 h | 0.106 ± 0.007 | 0.109 ± 0.008 B | ||||
| UREA (mg/dL) | D−2 T0 h | 41.650 ± 2.010 | 52.470 ± 2.440 | 0.319 | 0.032 | 0 |
| D−2 T4 h | 44.400 ± 2.300 | 59.880 ± 2.540 | ||||
| Triglycerides (mg/dL) | D−2 T0 h | 8.149 ± 0.379 a | 16.323 ± 0.831 | 0 | 0 | 0 |
| D−2 T4 h | 234.500 ± 37.300 b* | 15.103 ± 0.920 * | ||||
| Cholesterol (mg/dL) | D−2 T0 h | 44.880 ± 1.410 | 49.060 ± 1.030 | 0.919 | 0.022 | 0.001 |
| D−2 T4 h | 42.180 ± 1.380 | 46.110 ± 0.947 | ||||
| ALT (U/L) | D−2 T0 h | 25.510 ± 1.200 | 26.320 ± 0.992 | 0.85 | 0.674 | 0.57 |
| D−2 T4 h | 26.160 ± 1.150 | 26.566 ± 0.891 | ||||
| AST (U/L) | D−2 T0 h | 95.220 ± 2.460 | 102.250 ± 2.260 | 0.515 | 0.308 | 0.024 |
| D−2 T4 h | 99.220 ± 2.480 | 103.140 ± 2.330 | ||||
| Albumin (g/dL) | D−2 T0 h | 3.013 ± 0.031 | 3.110 ± 0.037 | 0.6 | 0.166 | 0.001 |
| D−2 T4 h | 2.946 ± 0.038 | 3.080 ± 0.033 | ||||
| Total protein (g/dL) | D−2 T0 h | 7.272 ± 0.093 | 7.315 ± 0.093 | 0.856 | 0.067 | 0.525 |
| D−2 T4 h | 7.079 ± 0.103 | 7.157 ± 0.091 | ||||
| Creatinine (mg/dL) | D−2 T0 h | 0.846 ± 0.016 | 0.855 ± 0.019 | 0.726 | 0.209 | 0.834 |
| D−2 T4 h | 0.831 ± 0.015 | 0.829 ± 0.016 | ||||
| Treatments | p Values | |||||
|---|---|---|---|---|---|---|
| GLU | GON | Groups | Days | Groups*Days | ||
| Milk yield (L) | D−10/D−9 | 0.753 ± 0.054 | 0.791 ± 0.033 | 0.76 | 0.224 | 0.603 |
| D−3/D−2 | 0.720 ± 0.054 | 0.710 ± 0.041 | ||||
| Milk fat (g/100 g milk) | D−10/D−9 | 7.176 ± 0.198 | 7.023 ± 0.275 | 0.668 | 0.002 | 0.731 |
| D−3/D−2 | 6.485 ± 0.167 | 6.468 ± 0.118 | ||||
| Protein (g/100 g milk) | D−10/D−9 | 5.883 ± 0.139 | 5.661 ± 0.210 | 0.2 | 0.987 | 0.895 |
| D−3/D−2 | 5.865 ± 0.160 | 5.684 ± 0.089 | ||||
| Lactose (g/100 g milk) | D−10/D−9 | 4.210 ± 0.102 | 4.257 ± 0.144 | 0.494 | 0.127 | 0.769 |
| D−3/D−2 | 3.988 ± 0.123 | 4.106 ± 0.111 | ||||
| Somatic cells (log 10 SCC/mL) | D−10/D−9 | 2.582 ± 0.104 | 2.543 ± 0.087 | 0.31 | 0.414 | 0.539 |
| D−3/D−2 | 2.722 ± 0.106 | 2.563 ± 0.091 | ||||
| Casein (g/100 g milk) | D−10/D−9 | 4.432 ± 0.121 | 4.308 ± 0.165 | 0.327 | 0.806 | 0.983 |
| D−3/D−2 | 4.403 ± 0.135 | 4.273 ± 0.082 | ||||
| Urea (mg/dL) | D−10/D−9 | 44.300 ± 1.670 a | 41.310 ± 2.020 | 0.085 | 0.002 | 0.001 |
| D−3/D−2 | 33.000 ± 1.440 b* | 41.810 ± 1.490 * | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sotgiu, F.D.; Porcu, C.; Pasciu, V.; Dattena, M.; Gallus, M.; Argiolas, G.; Berlinguer, F.; Molle, G. Towards a Sustainable Reproduction Management of Dairy Sheep: Glycerol-Based Formulations as Alternative to eCG in Milked Ewes Mated at the End of Anoestrus Period. Animals 2021, 11, 922. https://doi.org/10.3390/ani11040922
Sotgiu FD, Porcu C, Pasciu V, Dattena M, Gallus M, Argiolas G, Berlinguer F, Molle G. Towards a Sustainable Reproduction Management of Dairy Sheep: Glycerol-Based Formulations as Alternative to eCG in Milked Ewes Mated at the End of Anoestrus Period. Animals. 2021; 11(4):922. https://doi.org/10.3390/ani11040922
Chicago/Turabian StyleSotgiu, Francesca D., Cristian Porcu, Valeria Pasciu, Maria Dattena, Marilia Gallus, Giuseppe Argiolas, Fiammetta Berlinguer, and Giovanni Molle. 2021. "Towards a Sustainable Reproduction Management of Dairy Sheep: Glycerol-Based Formulations as Alternative to eCG in Milked Ewes Mated at the End of Anoestrus Period" Animals 11, no. 4: 922. https://doi.org/10.3390/ani11040922
APA StyleSotgiu, F. D., Porcu, C., Pasciu, V., Dattena, M., Gallus, M., Argiolas, G., Berlinguer, F., & Molle, G. (2021). Towards a Sustainable Reproduction Management of Dairy Sheep: Glycerol-Based Formulations as Alternative to eCG in Milked Ewes Mated at the End of Anoestrus Period. Animals, 11(4), 922. https://doi.org/10.3390/ani11040922