
Short-Term Variations of C18:1 Trans Fatty Acids in Plasma Lipoproteins and Ruminal Fermentation Parameters of Non-Lactating Cows Subjected to Ruminal Pulses of Oils




Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Treatments
2.2. Ruminal Fluid Samples
2.3. Plasma Samples
2.4. Fatty Acid Analysis
2.5. Statistical Analysis
3. Results and Discussion
3.1. Fatty Acid Profile of Oils and Emulsions
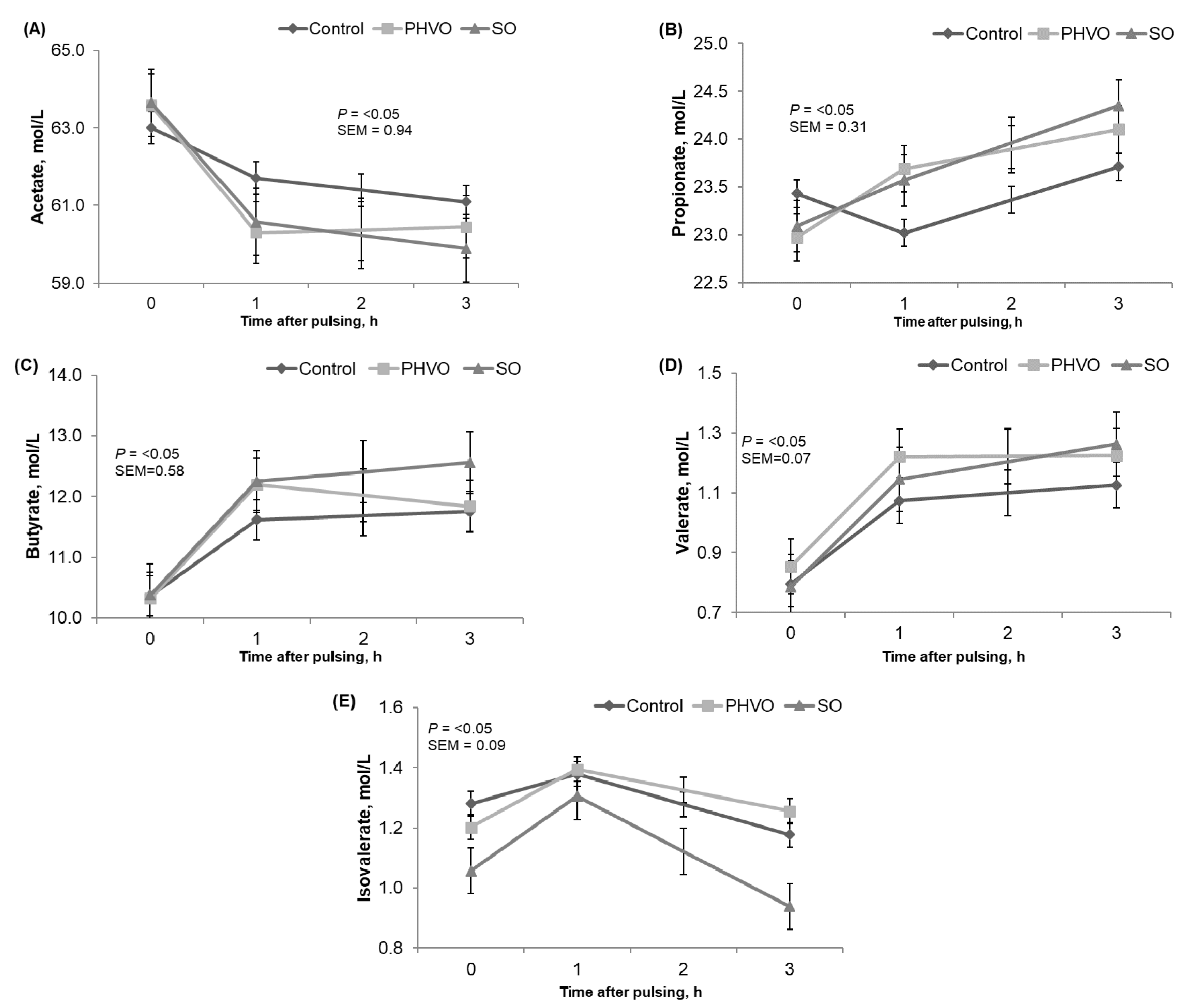
3.2. Ruminal Fermentation Parameters
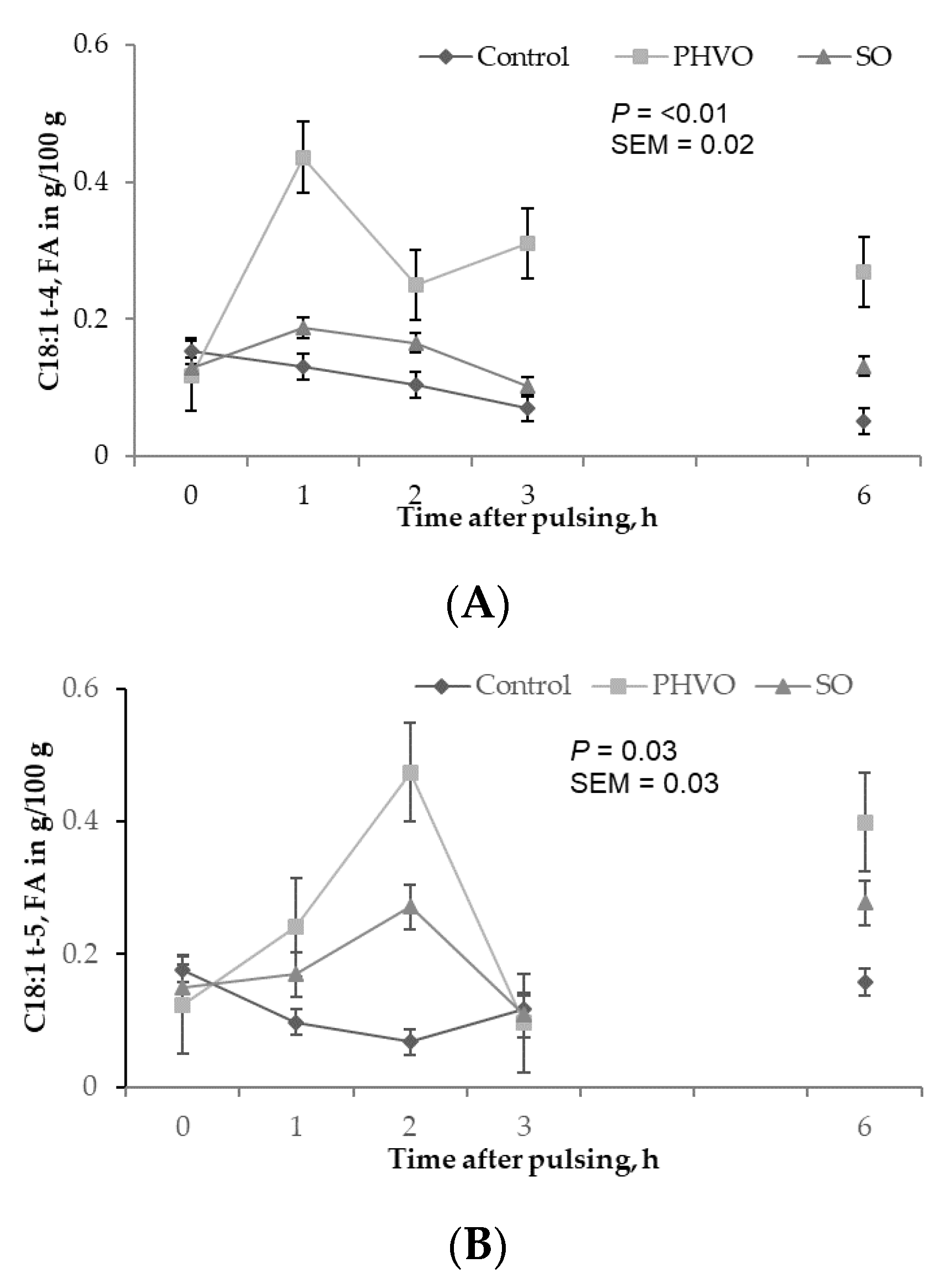
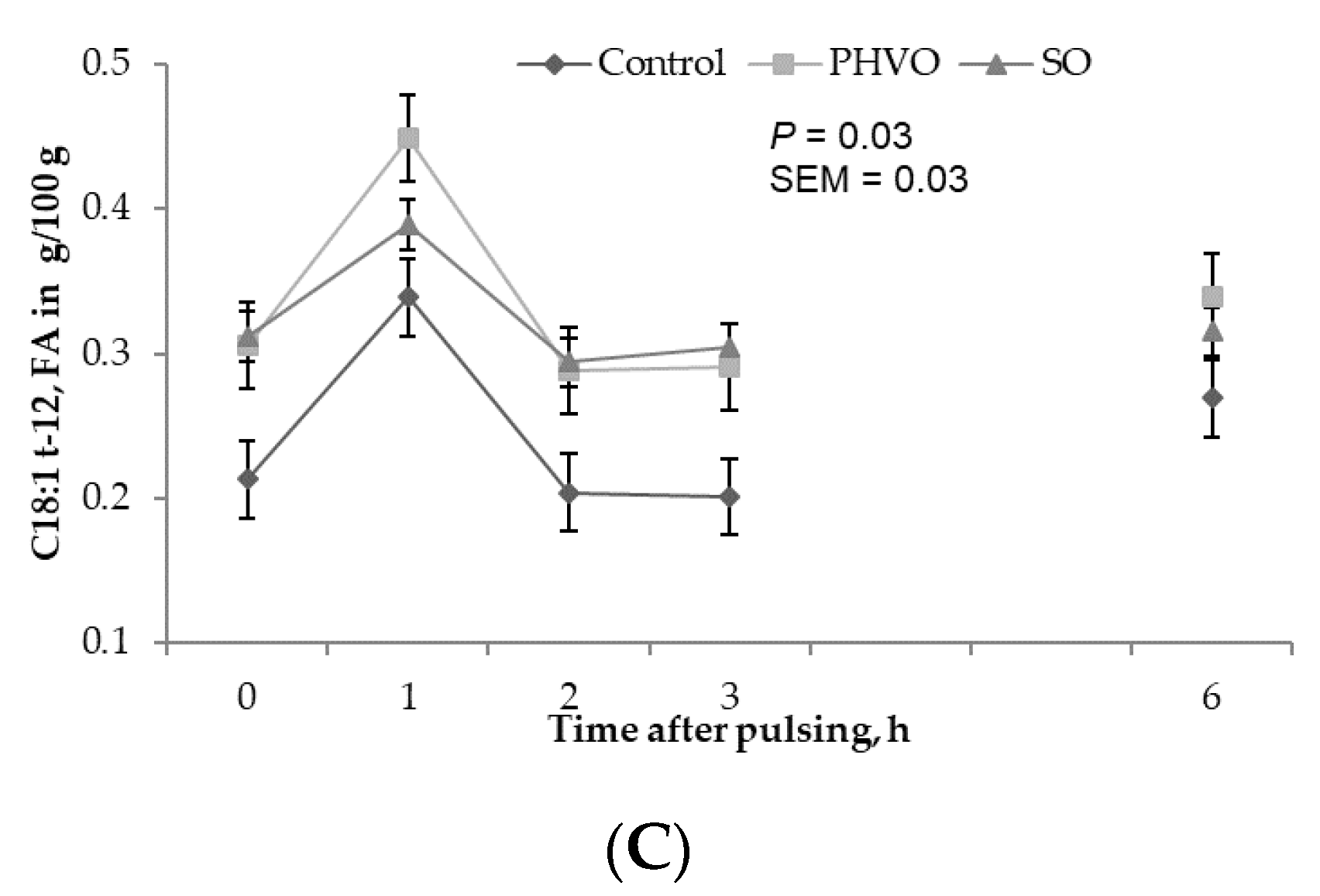
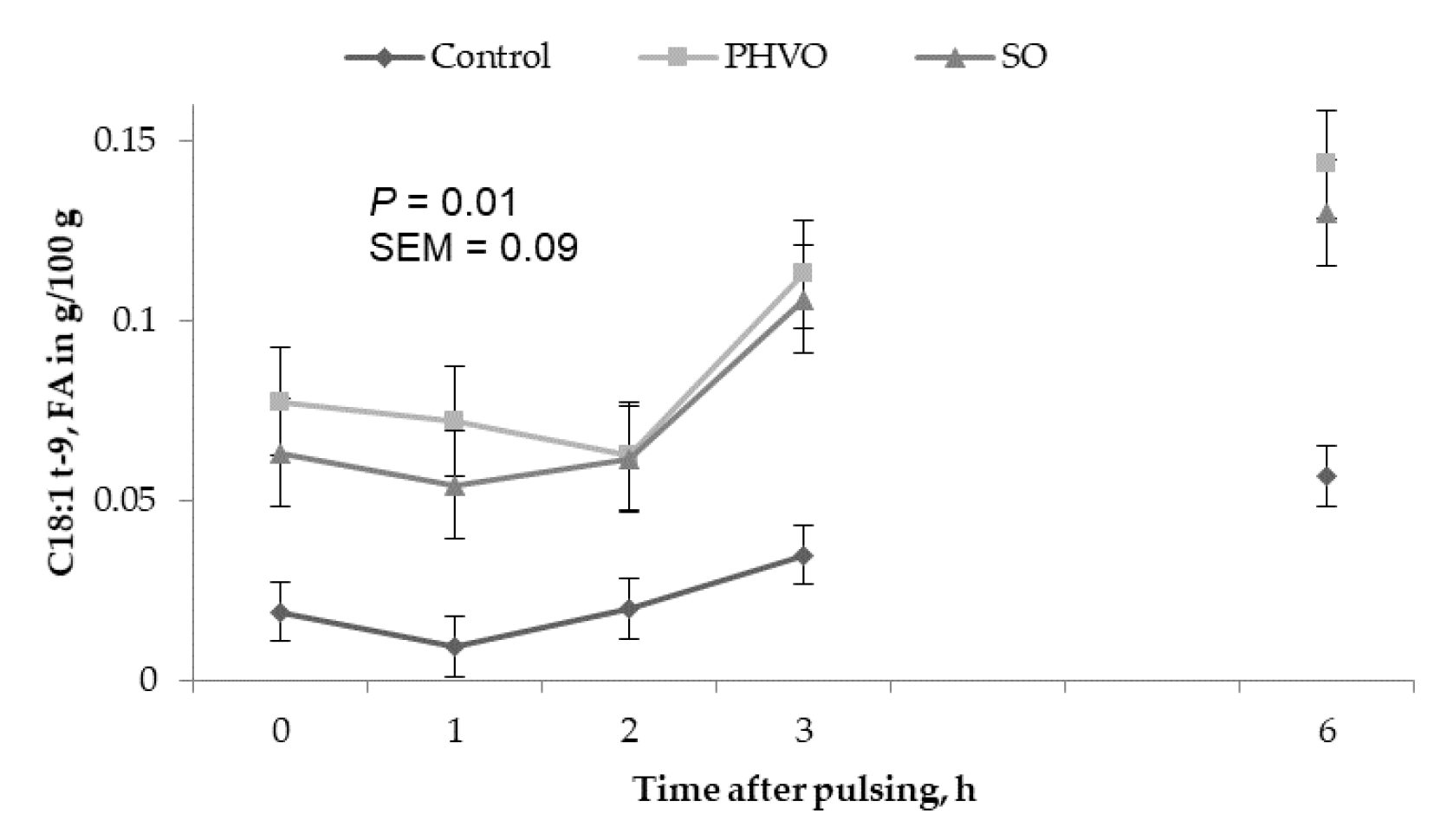
3.3. Treatment Effects on Plasma and Lipoprotein Fractions
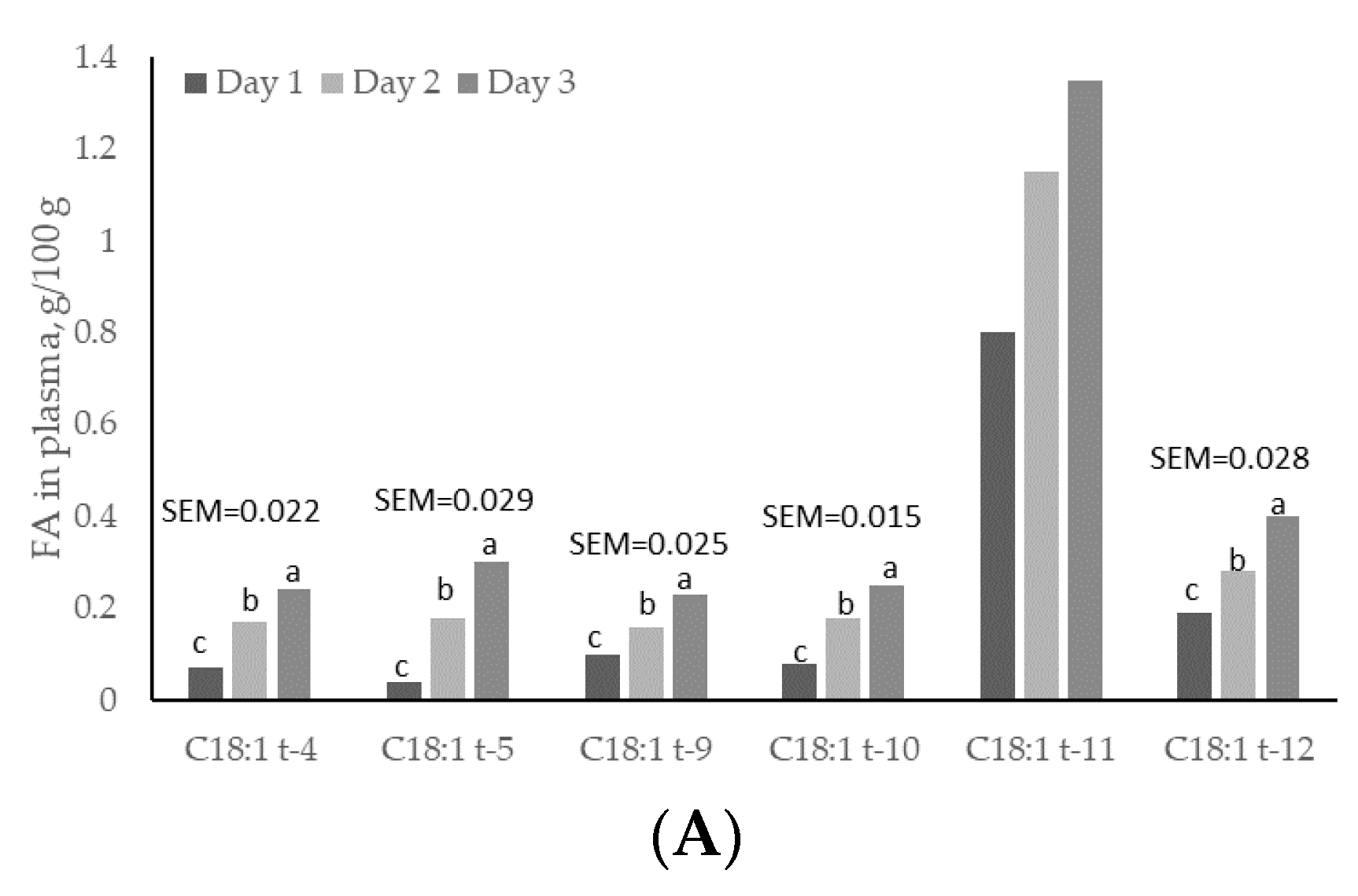
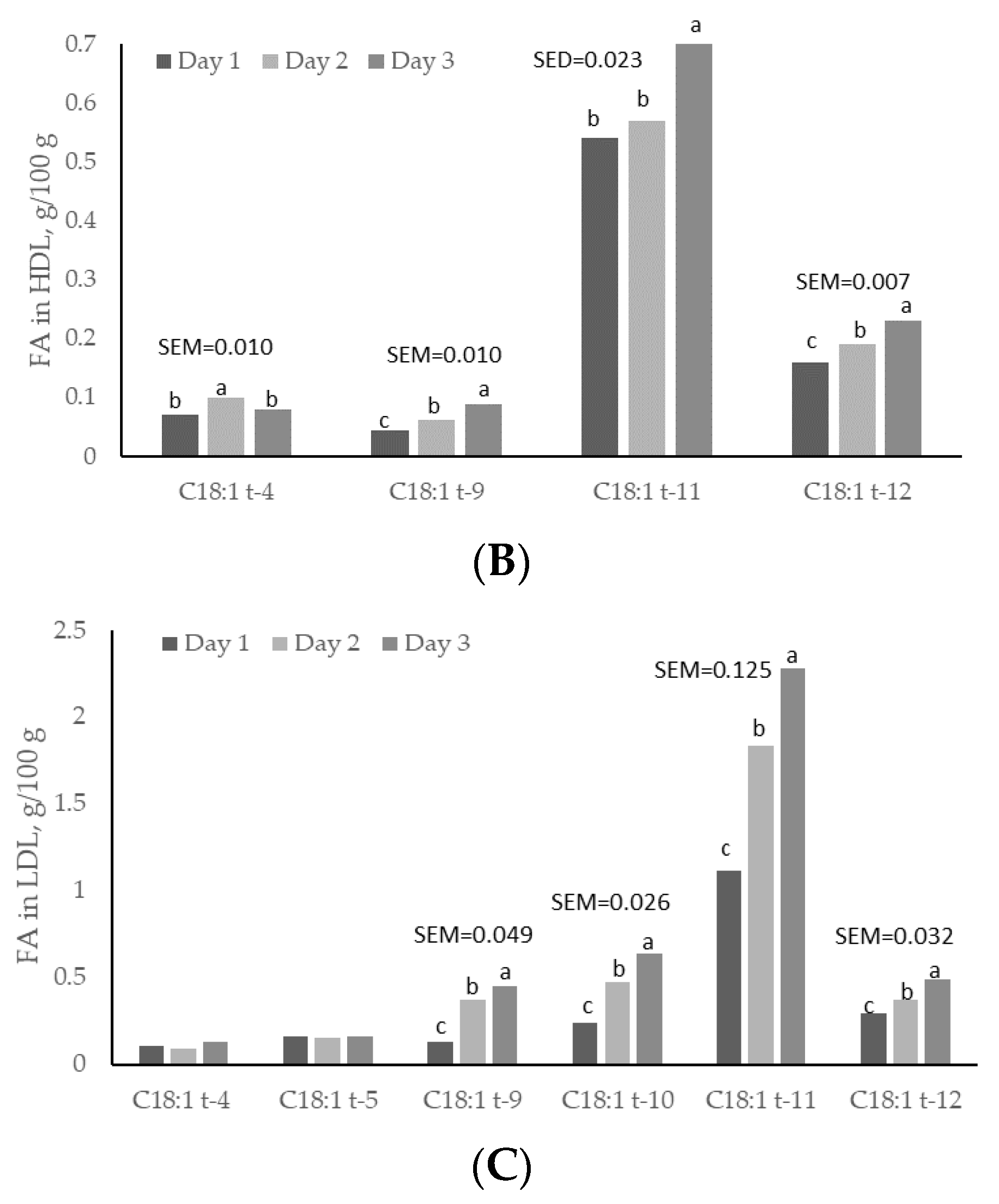
3.4. Plasma and Lipoproteins: Day and Time Changes
3.5. Further Considerations
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Piperova, L.S.; Moallem, U.; Teter, B.B.; Sampugna, J.; Yurawecz, M.P.; Morehouse, K.M.; Luchini, D.; Erdman, R.A. Changes in milk fat in response to dietary supplementation with calcium salts of trans-18:1 or conjugated linoleic fatty acids in lactating dairy cows. J. Dairy Sci. 2004, 87, 3836–3844. [Google Scholar] [CrossRef]
- Meijer, G.W.; van Tol, A.; van Berkel, T.J.C.; Weststrate, J.A. Effect of dietary elaidic versus vaccenic acid on blood and liver lipids in the hamster. Atherosclerosis 2001, 157, 31–40. [Google Scholar] [CrossRef]
- Kuhnt, K.; Baehr, M.; Rohrer, C.; Jahreis, G. Trans fatty acid isomers and the trans-9/trans-11 index in fat containing foods. Eur. J. Lipid. Sci. Technol. 2011, 113, 1281–1292. [Google Scholar] [CrossRef]
- Oteng, A.-B.; Kersten, S. Mechanisms of Action of trans Fatty Acids. Adv. Nutr. 2020, 11, 697–708. [Google Scholar] [CrossRef] [PubMed]
- Bernard, L.; Bonnet, M.; Delavaud, C.; Delosière, M.; Ferlay, A.; Fougère, H.; Graulet, B. Milk Fat Globule in Ruminant: Major and Minor Compounds, Nutritional Regulation and Differences Among Species. Eur. J. Lipid Sci. Technol. 2018, 120, 1700039. [Google Scholar] [CrossRef]
- Vargas-Bello-Pérez, E.; Fehrmann-Cartes, K.; Íñiguez-González, G.; Toro-Mujica, P.; Garnsworthy, P.C. Short communication: Chemical composition, fatty acid composition, and sensory characteristics of Chanco cheese from dairy cows supplemented with soybean and hydrogenated vegetable oils. J. Dairy Sci. 2015, 98, 111–117. [Google Scholar] [CrossRef]
- Lock, A.L.; Garnsworthy, P.C. Independent effects of dietary linoleic and linolenic fatty acids on the conjugated linoleic acid content of cows’ milk. Anim. Sci. 2002, 74, 163–176. [Google Scholar] [CrossRef]
- Ahmad, F.; Sun, Q.; Patel, D.; Stommel, J.M. Cholesterol metabolism: A potential therapeutic target in glioblastoma. Cancers 2019, 11, 146. [Google Scholar] [CrossRef] [PubMed]
- Offer, N.W.; Speake, B.K.; Dixon, J.; Marsden, M. Effect of fish-oil supplementation on levels of (n-3) poly-unsaturated fatty acids in the lipoprotein fractions of bovine plasma. Anim. Sci. 2001, 73, 523–531. [Google Scholar] [CrossRef]
- Sun, X.Q.; Gibbs, S.J. Diurnal variation in fatty acid profiles in rumen digesta from dairy cows grazing high-quality pasture. Anim. Feed Sci. Technol. 2012, 177, 152–160. [Google Scholar] [CrossRef]
- Loor, J.J.; Ueda, K.; Ferlay, A.; Chilliard, Y.; Doreau, M. Short Communication: Diurnal Profiles of Conjugated Linoleic Acids and Trans Fatty Acids in Ruminal Fluid from Cows Fed a High Concentrate Diet Supplemented with Fish Oil, Linseed Oil, or Sunflower Oil. J. Dairy Sci. 2004, 87, 2468–2471. [Google Scholar] [CrossRef]
- Colman, E.; Tas, B.M.; Waegeman, W.; De Baets, B.; Fievez, V. The logistic curve as a tool to describe the daily ruminal pH pattern and its link with milk fatty acids. J. Dairy Sci. 2012, 95, 5845–5865. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Bello-Pérez, E.; Íñiguez-González, G.; Cancino-Padilla, N.; Loor, J.J.; Garnsworthy, P.C. Effect of dietary vegetable oils on the fatty acid profile of plasma lipoproteins in dairy cows. Arch. Anim. Nutr. 2016, 70, 322–332. [Google Scholar] [CrossRef]
- Vargas-Bello-Pérez, E.; Íñiguez-González, G.; Garnsworthy, P.C.; Loor, J.J. Transport of fatty acids within plasma lipoproteins in lactating and non-lactating cows fed on fish oil and hydrogenated palm oil. J. Anim. Physiol. Anim. Nutr. 2017, 101, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Boudon, A.; Faverdin, P.; Delagarde, R.; Lamberton, P.; Peyraud, J.L. Effects of Rumen or Duodenal Glucose Infusions on Intake in Dairy Cows Fed Fresh Perennial Ryegrass Indoors. J. Dairy Sci. 2007, 90, 4397–4410. [Google Scholar] [CrossRef] [PubMed]
- Doreau, M.; Batisse, V.; Bauchart, D. Apprrciation de l’hydrogrnation des acides gras alimentaires darts le rumen de la vache: 6tude mrthodologique preliminaire. Ann. Zootech. 1989, 38, 139–144. [Google Scholar] [CrossRef]
- Vargas-Bello-Pérez, E.; Geldsetzer-Mendoza, C.; Morales, M.S.; Toro-Mujica, P.; Fellenberg, M.A.; Ibáñez, R.A.; Gómez-Cortés, P.; Garnsworthy, P.C. Effect of olive oil in dairy cow diets on the fatty acid profile and sensory characteristics of cheese. Int. Dairy J. 2018, 85, 8–15. [Google Scholar] [CrossRef]
- Vargas-Bello-Pérez, E.; Cancino-Padilla, N.; Geldsetzer-Mendoza, C.; Vyhmeister, S.; Morales, M.S.; Leskinen, H.; Romero, J.; Garnsworthy, P.C.; Ibáñez, R.A. Effect of Feeding Cows with Unsaturated Fatty Acid Sources on Milk Production, Milk Composition, Milk Fatty Acid Profile, and Physicochemical and Sensory Characteristics of Ice Cream. Animals 2019, 9, 568. [Google Scholar] [CrossRef]
- Contreras, G.A.; O’Boyle, N.J.; Herdt, T.H.; Sordillo, L.M. Lipomobilization in periparturient dairy cows influences the composition of plasma nonesterified fatty acids and leukocyte phospholipid fatty acids. J. Dairy Sci. 2010, 93, 2508–2516. [Google Scholar] [CrossRef]
- Watts, J.S.; Rezamand, P.; Sevier, D.L.; Price, W.; McGuire, M.A. Short-term effects of dietary trans fatty acids compared with saturated fatty acids on selected measures of inflammation, fatty acid profiles, and production in early lactating Holstein dairy cows. J. Dairy Sci. 2013, 96, 6932–6943. [Google Scholar] [CrossRef]
- Andersen, J.B.; Ridder, C.; Larsen, T. Priming the Cow for Mobilization in the Periparturient Period: Effects of Supplementing the Dry Cow with Saturated Fat or Linseed. J. Dairy Sci. 2008, 91, 1029–1043. [Google Scholar] [CrossRef]
- Santschi, D.E.; Wettstein, H.-R.; Leiber, F.; Witschi, A.-K.M.; Kreuzer, M. Colostrum and milk fatty acids of dairy cows as influenced by extruded linseed supplementation during the transition period. Can. J. Anim. Sci. 2009, 89, 383–392. [Google Scholar] [CrossRef]
- Leiber, F.; Hochstrasser, R.; Wettstein, H.R.; Kreuzer, M. Feeding transition cows with oilseeds: Effects on fatty acid composition of adipose tissue, colostrum and milk. Livest. Sci. 2011, 138, 1–12. [Google Scholar] [CrossRef]
- Petit, H.V.; Benchaar, C. Milk production, milk composition, blood composition, and conception rate of transition dairy cows fed different profiles of fatty acids. Can. J. Anim. Sci. 2007, 87, 591–600. [Google Scholar] [CrossRef]
- Badiei, A.; Aliverdilou, A.; Amanlou, H.; Beheshti, M.; Dirandeh, E.; Masoumi, R.; Moosakhani, F.; Petit, H.V. Postpartum responses of dairy cows supplemented with n-3 fatty acids for different durations during the peripartal period. J. Dairy Sci. 2014, 97, 6391–6399. [Google Scholar] [CrossRef] [PubMed]
- Agriculture and Food Research Council. Energy and Protein Requirements of Ruminants. An Advisory Manual Prepared by the AFRC Technical Committee on Responses to Nutrients; CAB International: Wallingford, UK, 1993. [Google Scholar]
- Ferlay, A.; Doreau, M. Influence of Method of Administration of Rapeseed Oil in Dairy Cows. 1. Digestion of Nonlipid Components. J. Dairy Sci. 1992, 75, 3020–3027. [Google Scholar] [CrossRef]
- Radding, C.M.; Steinberg, D. Studies on the synthesis and secretion of serum lipoproteins by rat liver slices. J. Clin. Investig. 1960, 39, 1560–1569. [Google Scholar] [CrossRef]
- Hara, A.; Radin, N.S. Lipid extraction of tissues with a low-toxicity solvent. Anal. Biochem. 1978, 90, 420–426. [Google Scholar] [CrossRef]
- Chichlowski, M.W.; Schroeder, J.W.; Park, C.S.; Keller, W.L.; Schimek, D.E. Altering the fatty acids in milk fat by including canola seed in dairy cattle diets. J. Dairy Sci. 2005, 88, 3084–3094. [Google Scholar] [CrossRef]
- Harvatine, K.J.; Allen, M.S. Effects of Fatty Acid Supplements on Ruminal and Total Tract Nutrient Digestion in Lactating Dairy Cows. J. Dairy Sci. 2006, 89, 1092–1103. [Google Scholar] [CrossRef]
- Hristov, A.N.; Ropp, J.K. Effect of Dietary Carbohydrate Composition and Availability on Utilization of Ruminal Ammonia Nitrogen for Milk Protein Synthesis in Dairy Cows. J. Dairy Sci. 2003, 86, 2416–2427. [Google Scholar] [CrossRef]
- Knothe, G.; Dunn, R.O. A Comprehensive Evaluation of the Melting Points of Fatty Acids and Esters Determined by Differential Scanning Calorimetry. J. Am. Oil Chem. Soc. 2009, 86, 843–856. [Google Scholar] [CrossRef]
- Vargas-Bello-Pérez, E.; Cancino-Padilla, N.; Romero, J.; Garnsworthy, P.C. Quantitative analysis of ruminal bacterial populations involved in lipid metabolism in dairy cows fed different vegetable oils. Animal 2016, 10, 1821–1828. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.L.; Bu, D.P.; Wang, J.Q.; Hu, Z.Y.; Li, D.; Wei, H.Y.; Zhou, L.Y.; Loor, J.J. Soybean oil and linseed oil supplementation affect profiles of ruminal microorganisms in dairy cows. Animal 2009, 3, 1562–1569. [Google Scholar] [CrossRef]
- Guyader, J.; Eugène, M.; Nozière, P.; Morgavi, D.P.; Doreau, M.; Martin, C. Influence of rumen protozoa on methane emission in ruminants: A meta-analysis approach. Animal 2014, 8, 1816–1825. [Google Scholar] [CrossRef] [PubMed]
- Jordan, E.; Lovett, D.K.; Monahan, F.J.; Callan, J.; Flynn, B.; O’Mara, F.P. Effect of refined coconut oil or copra meal on methane output and on intake and performance of beef heifers. J. Anim. Sci. 2006, 84, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Hackmann, T.J.; Firkins, J.L. Maximizing efficiency of rumen microbial protein production. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef]
- Gustafsson, A.H.; Palmquist, D.L. Diurnal variation of rumen ammonia, serum urea, and milk urea in dairy cows at high and low yields. J. Dairy Sci. 1993, 76, 475–484. [Google Scholar] [CrossRef]
- Roy, A.; Mandal, G.P.; Patra, A.K. Effects of different vegetable oils on rumen fermentation and conjugated linoleic acid concentration in vitro. Vet. World 2017, 10, 11–16. [Google Scholar] [CrossRef][Green Version]
- Bauman, D.E.; Mather, I.H.; Wall, R.J.; Lock, A.L. Major advances associated with the biosynthesis of milk. J. Dairy Sci. 2006, 89, 1235–1243. [Google Scholar] [CrossRef]
- Bauman, D.E.; Harvatine, K.J.; Lock, A.L. Nutrigenomics, Rumen-Derived Bioactive Fatty Acids, and the Regulation of Milk Fat Synthesis. Annu. Rev. Nutr. 2011, 31, 299–319. [Google Scholar] [CrossRef]
- Grażyna, C.; Hanna, C.; Adam, A.; Magdalena, B.M. Natural antioxidants in milk and dairy products. Int. J. Dairy Technol. 2017, 70, 165–178. [Google Scholar] [CrossRef]
- Young, J.F.; Therkildsen, M.; Ekstrand, B.; Che, B.N.; Larsen, M.K.; Oksbjerg, N.; Stagsted, J. Novel aspects of health promoting compounds in meat. Meat Sci. 2013, 95, 904–911. [Google Scholar] [CrossRef] [PubMed]
- Doreau, M.; Ferlay, A. Digestion and utilisation of fatty acids by ruminants. Anim. Feed Sci. Technol. 1994, 45, 379–396. [Google Scholar] [CrossRef]
- Glasser, F.; Ferlay, A.; Doreau, M.; Schmidely, P.; Sauvant, D.; Chilliard, Y. Long-Chain Fatty Acid Metabolism in Dairy Cows: A Meta-Analysis of Milk Fatty Acid Yield in Relation to Duodenal Flows and De Novo Synthesis. J. Dairy Sci. 2008, 91, 2771–2785. [Google Scholar] [CrossRef]
- Arrigoni, M.D.B.; Martins, C.L.; Factori, M.A. Lipid metabolism in the rumen. In Rumenology; Springer: Berlin/Heidelberg, Germany, 2016; pp. 103–126. [Google Scholar]
- Antongiovanni, M.; Buccioni, A.; Petacchi, F.; Secchiari, P.; Mele, M.; Serra, A. Upgrading the lipid fraction of foods of animal origin by dietary means: Rumen activity and presence of trans fatty acids and CLA in milk and meat. Ital. J. Anim. Sci. 2003, 2, 3–28. [Google Scholar] [CrossRef]
- Jenkins, T.C. Lipid Metabolism in the Rumen. J. Dairy Sci. 1993, 76, 3851–3863. [Google Scholar] [CrossRef]
- Christie, W.W. The composition, structure and function of lipids in the tissues of ruminant animals. In Lipid Metabolism in Ruminant Animals; Christie, Ed.; Pergamon Press: New York, NY, USA, 1981; p. 95. [Google Scholar]
- Bitman, J.; Wood, D.L.; Lefcourt, A.M. Rhythms in Cholesterol, Cholesteryl Esters, Free Fatty Acids, and Triglycerides in Blood of Lactating Dairy Cows. J. Dairy Sci. 1990, 73, 948–955. [Google Scholar] [CrossRef]
- Palmquist, D.L.; Jenkins, T.C. A 100-Year Review: Fat feeding of dairy cows. J. Dairy Sci. 2017, 100, 10061–10077. [Google Scholar] [CrossRef]
- Kim, E.J.; Huws, S.A.; Lee, M.R.F.; Scollan, N.D. Dietary Transformation of Lipid in the Rumen Microbial Ecosystem. Asian-Australas. J. Anim. Sci. 2009, 22, 1341–1350. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentrate 1 | |
|---|---|
| Ingredient composition (% of dry matter) | |
| Rolled barley | 50 |
| Sugar beet pulp | 15 |
| Rolled wheat | 15 |
| Cane molasses | 5 |
| Limestone flour | 5 |
| Vitamins | 5 |
| Minerals | 5 |
| Chemical composition (% of dry matter) | |
| Neutral detergent fiber | 8 |
| Crude protein | 14 |
| Ether extract | 3 |
| Ash | 7 |
| Oils | Treatment Emulsions 4 | ||||
|---|---|---|---|---|---|
| Fatty Acid, g/100 g | SO 1 | PHVO 2 | Control 3 | SO + SM | PHVO + SM |
| C10:0 | - | - | 2.51 | - | - |
| C12:0 | - | 0.23 | 2.14 | - | 0.24 |
| C14:0 | 0.07 | 0.18 | 7.89 | 0.08 | 0.30 |
| C14:1 | - | - | 0.50 | - | - |
| C15:0 | - | - | 0.98 | - | - |
| C16:0 | 10.74 | 7.82 | 27.02 | 10.84 | 7.80 |
| C16:1 | - | - | 1.54 | - | 0.13 |
| C17:0 | - | - | 0.63 | 0.08 | 0.07 |
| C18:0 | 4.28 | 8.35 | 11.69 | 4.30 | 8.15 |
| C18:1 trans-4 | - | 0.59 | - | - | 0.48 |
| C18:1 trans-5 | - | 1.48 | - | - | 1.38 |
| C18:1 trans-6–8 | - | 8.39 | - | - | 8.33 |
| C18:1 trans-9 | - | 17.60 | - | - | 16.67 |
| C18:1 trans-10 | - | 9.28 | - | - | 8.96 |
| C18:1 trans-11 | - | 7.67 | 1.70 | - | 7.54 |
| C18:1 trans-12 | - | 7.35 | - | - | 6.97 |
| C18:1 cis-9 | 22.42 | 6.06 | 21.08 | 22.33 | 6.27 |
| C18:2 trans-9, cis-12 | - | 1.13 | - | - | 0.84 |
| C18:2 cis-9, cis-12 | 53.03 | 0.42 | 3.39 | 52.90 | 0.46 |
| C20:0 | 0.34 | 0.76 | - | 0.34 | 0.73 |
| C18:3 cis-6, 9, 12 | - | 0.42 | - | - | 0.58 |
| C20:1 | 0.20 | - | - | 0.18 | 0.15 |
| C18:3 cis-9, 12, 15 | 6.78 | 0.06 | 0.61 | 6.75 | 0.14 |
| C18:2 cis-9, trans-11 | - | - | 1.03 | - | - |
| C20:2 | 0.05 | 0.05 | - | - | - |
| C22:0 | 0.38 | 0.44 | - | 0.37 | 0.42 |
| C24:0 | 0.15 | 0.15 | - | 0.14 | 0.18 |
| Other 5 | 1.56 | 21.57 | 17.29 | 1.69 | 23.21 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vargas-Bello-Pérez, E.; Loor, J.J.; Garnsworthy, P.C. Short-Term Variations of C18:1 Trans Fatty Acids in Plasma Lipoproteins and Ruminal Fermentation Parameters of Non-Lactating Cows Subjected to Ruminal Pulses of Oils. Animals 2021, 11, 788. https://doi.org/10.3390/ani11030788
Vargas-Bello-Pérez E, Loor JJ, Garnsworthy PC. Short-Term Variations of C18:1 Trans Fatty Acids in Plasma Lipoproteins and Ruminal Fermentation Parameters of Non-Lactating Cows Subjected to Ruminal Pulses of Oils. Animals. 2021; 11(3):788. https://doi.org/10.3390/ani11030788
Chicago/Turabian StyleVargas-Bello-Pérez, Einar, Juan J. Loor, and Philip C. Garnsworthy. 2021. "Short-Term Variations of C18:1 Trans Fatty Acids in Plasma Lipoproteins and Ruminal Fermentation Parameters of Non-Lactating Cows Subjected to Ruminal Pulses of Oils" Animals 11, no. 3: 788. https://doi.org/10.3390/ani11030788
APA StyleVargas-Bello-Pérez, E., Loor, J. J., & Garnsworthy, P. C. (2021). Short-Term Variations of C18:1 Trans Fatty Acids in Plasma Lipoproteins and Ruminal Fermentation Parameters of Non-Lactating Cows Subjected to Ruminal Pulses of Oils. Animals, 11(3), 788. https://doi.org/10.3390/ani11030788