Vertebral Formulae and Congenital Vertebral Anomalies in Guinea Pigs: A Retrospective Radiographic Study



Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Radiographic Evaluation
2.2. Data Analysis
3. Results
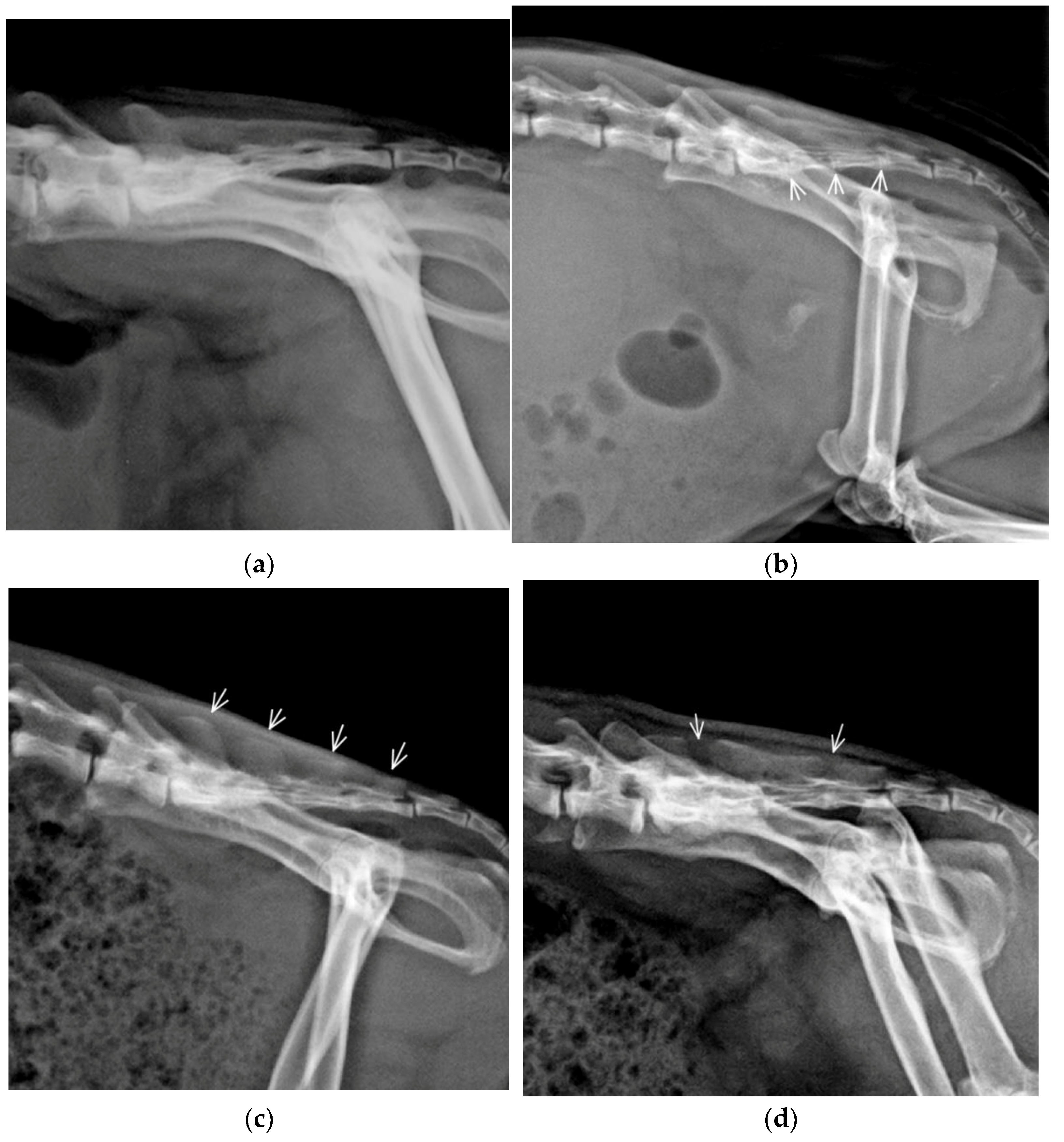
3.1. Vertebral Formulae and Anatomic Variants
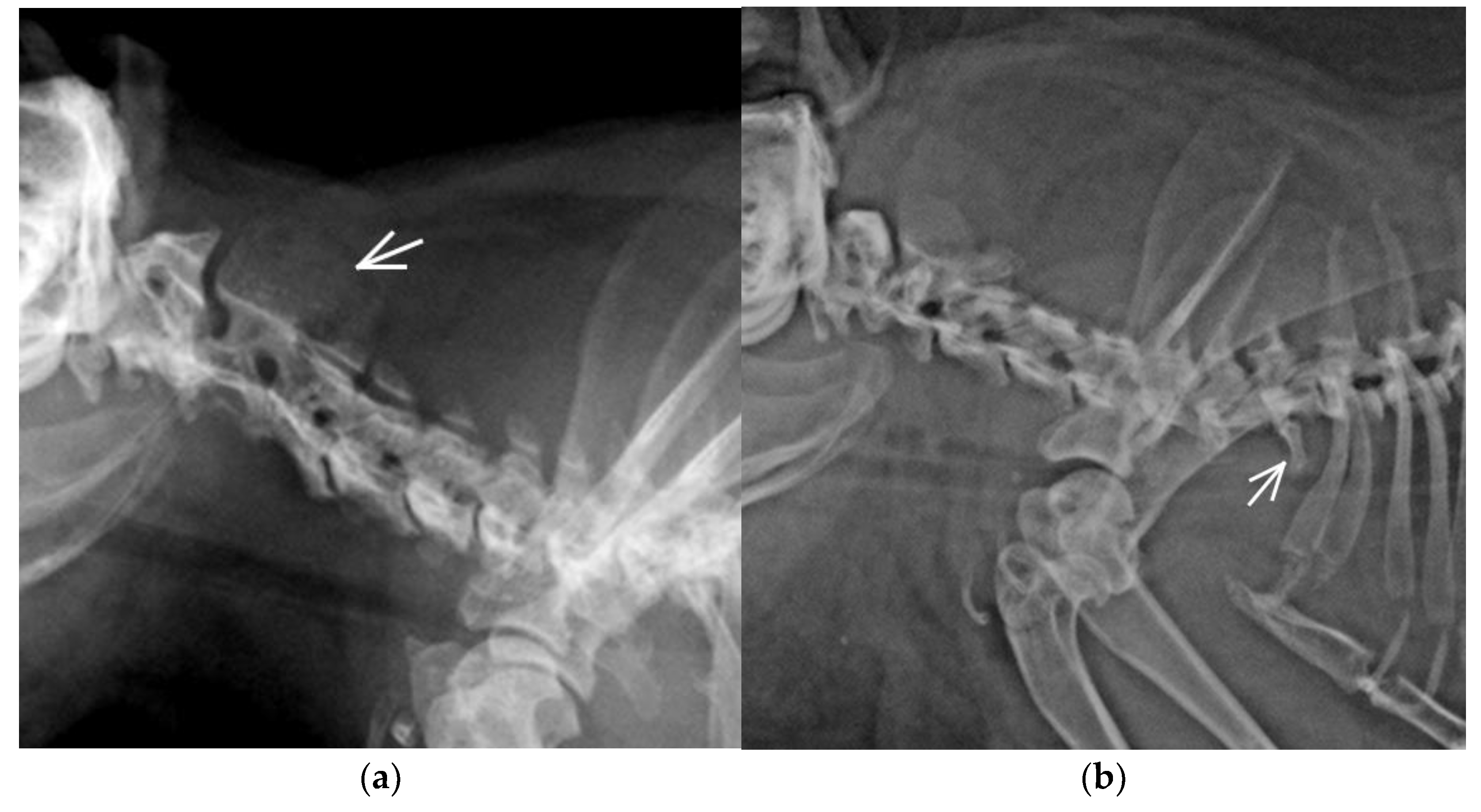
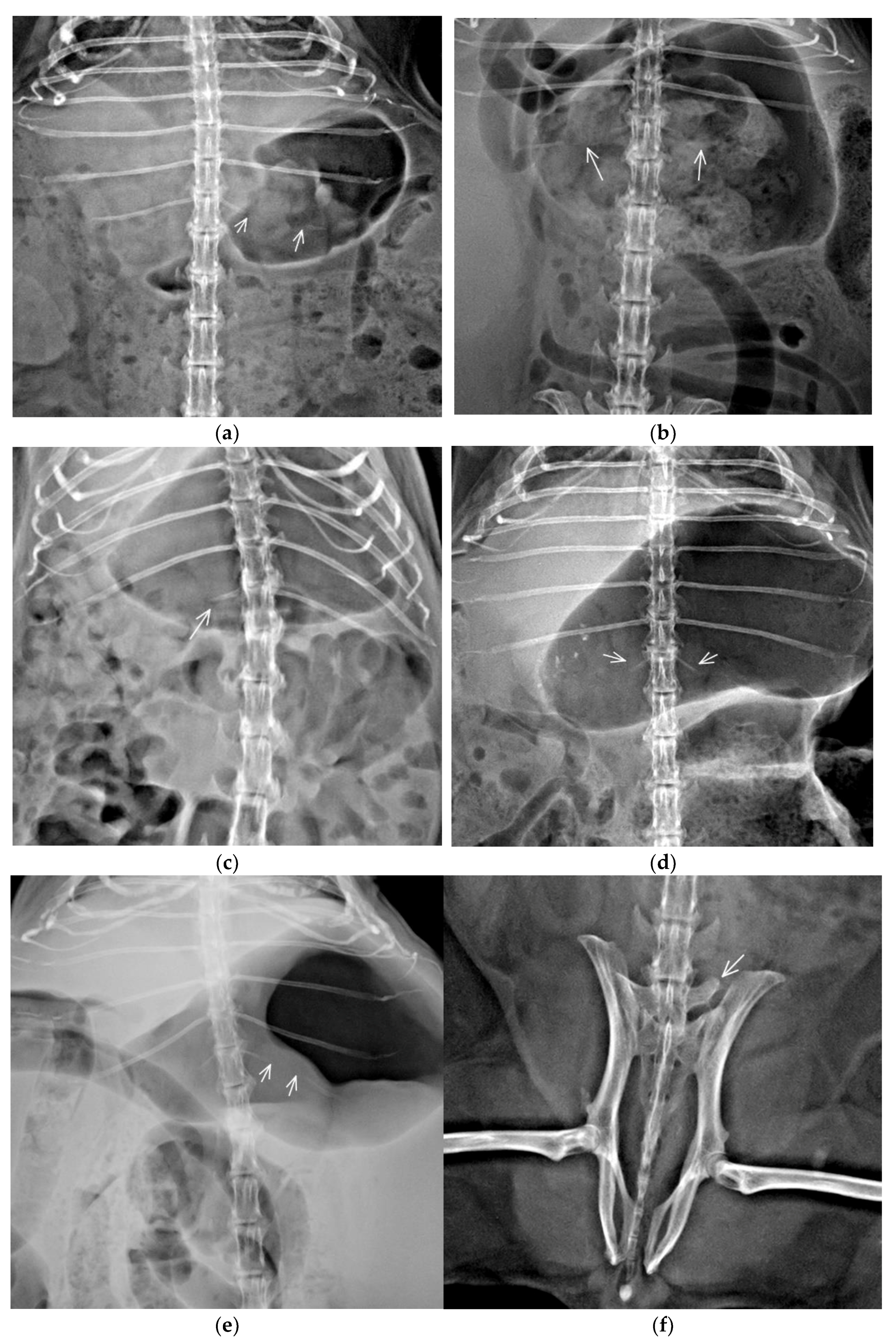
3.2. Congenital Vertebral Anomalies
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sánchez-Macías, D.; Castro, N.; Rivero, M.A.; Argüello, A.; Morales-delaNuez, A. Proposal for standard methods and procedure for guinea pig carcass evaluation, jointing and tissue separation. J. Appl. Anim. Res. 2016, 44, 65–70. [Google Scholar] [CrossRef]
- Jaeger, C.B.; Blight, A.R. Spinal cord compression injury in guinea pigs: Structural changes of endothelium and its perivascular cell associations after blood-brain barrier breakdown and repair. Exp. Neurol. 1997, 144, 381–399. [Google Scholar] [CrossRef]
- Brown, C.J.; Donnelly, T.M. Rodent husbandry and care. Vet. Clin. N. Am. Exot. Anim. Pract. 2004, 7, 201–225. [Google Scholar] [CrossRef]
- Pignon, C.; Mayer, J. 21—Guinea Pigs. In Ferrets, Rabbits, and Rodents, 4th ed.; Quesenberry, K.E., Orcutt, C.J., Mans, C., Carpenter, J.W., Eds.; W.B. Saunders: Philadelphia, PA, USA, 2021; pp. 270–297. [Google Scholar] [CrossRef]
- Nielsen, T.D.; Dean, R.S.; Robinson, N.J.; Massey, A.; Brennan, M.L. Survey of the UK veterinary profession: Common species and conditions nominated by veterinarians in practice. Vet. Rec. 2014, 174, 324. [Google Scholar] [CrossRef]
- Witkowska, A.; Alibhai, A.; Hughes, C.; Price, J.; Klisch, K.; Sturrock, C.J.; Rutland, C.S. Computed tomography analysis of guinea pig bone: Architecture, bone thickness and dimensions throughout development. PeerJ 2014, 2, e615. [Google Scholar] [CrossRef]
- Narita, Y.; Kuratani, S. Evolution of the vertebral formulae in mammals: A perspective on developmental constraints. J. Exp. Zool. B Mol. Dev. Evol. 2005, 304, 91–106. [Google Scholar] [CrossRef] [PubMed]
- Breazile, J.E.; Brown, E.M. Chapter 6—Anatomy. In The Biology of the Guinea Pig; Wagner, J.E., Manning, P.J., Eds.; Academic Press: San Diego, CA, USA, 1976; pp. 53–62. [Google Scholar] [CrossRef]
- Cooper, G.; Schiller, A.L. Anatomy of the Guinea Pig; Harvard University Press: Cambridge, MA, USA, 1975. [Google Scholar]
- Reese, S.; Fehr, M. Small mammals: Radioanatomy. In Diagnostic Imaging of Exotic Pets: Birds, Small Mammals, Reptiles; Krautwald-Junghanns, M.E., Pees, M., Reese, S., Tully, T., Eds.; Schlüttersche: Hannover, Germany, 2011; pp. 158–165. [Google Scholar]
- Lossi, L.; D’Angelo, L.; De Girolamo, P.; Merighi, A. Anatomical features for an adequate choice of experimental animal model in biomedicine: II. Small laboratory rodents, rabbit, and pig. Ann. Anat. 2016, 204, 11–28. [Google Scholar] [CrossRef] [PubMed]
- Clemons, D.J.; Seeman, J.L. The Laboratory Guinea Pig, 2nd ed.; Taylor & Francis: Boca Raton, FL, USA, 2018. [Google Scholar]
- Borgens, R.B.; Shi, R.; Bohnert, D. Behavioral recovery from spinal cord injury following delayed application of polyethylene glycol. J. Exp. Biol. 2002, 205, 1–12. [Google Scholar] [PubMed]
- Meredith, A.; Richardson, J. Neurological diseases of rabbits and rodents. J. Exot. Pet Med. 2015, 24, 21–33. [Google Scholar] [CrossRef]
- Yan, Y.Z.; Li, Q.P.; Wu, C.C.; Pan, X.X.; Shao, Z.X.; Chen, S.Q.; Wang, K.; Chen, X.B.; Wang, X.Y. Rate of presence of 11 thoracic vertebrae and 6 lumbar vertebrae in asymptomatic Chinese adult volunteers. J. Orthop. Surg. Res. 2018, 13, 124. [Google Scholar] [CrossRef]
- Newitt, A.; German, A.J.; Barr, F.J. Congenital abnormalities of the feline vertebral column. Vet. Radiol. Ultrasound 2008, 49, 35–41. [Google Scholar] [CrossRef]
- Morgan, J.P. Congenital Anomalies of the Vertebral Column of the Dog: A Study of the Incidence and Significance Based on a Radiographic and Morphologic Study 1. Vet. Radiol. 1968, 9, 21–29. [Google Scholar] [CrossRef]
- Proks, P.; Stehlik, L.; Paninarova, M.; Irova, K.; Hauptman, K.; Jekl, V. Congenital abnormalities of the vertebral column in ferrets. Vet. Radiol. Ultrasound 2015, 56, 117–123. [Google Scholar] [CrossRef]
- Westworth, D.R.; Sturges, B.K. Congenital spinal malformations in small animals. Vet. Clin. N. Am. Small Anim. Pract. 2010, 40, 951–981. [Google Scholar] [CrossRef]
- Gutierrez-Quintana, R.; Guevar, J.; Stalin, C.; Faller, K.; Yeamans, C.; Penderis, J. A proposed radiographic classification scheme for congenital thoracic vertebral malformations in brachycephalic “screw-tailed” dog breeds. Vet. Radiol. Ultrasound 2014, 55, 585–591. [Google Scholar] [CrossRef]
- Flückiger, M.; Geissbühler, U.; Lang, J. Lumbosacral transitional vertebrae: What is their impact on the health of affected dogs? Schweiz. Arch. Tierheilkd. 2009, 151, 133–135. [Google Scholar] [CrossRef]
- Proks, P.; Stehlik, L.; Nyvltova, I.; Necas, A.; Vignoli, M.; Jekl, V. Vertebral formula and congenital abnormalities of the vertebral column in rabbits. Vet. J. 2018, 236, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.; Halalmeh, D.R.; Sandio, A.; Tubbs, R.S.; Moisi, M.D. Anatomical Variations That Can Lead to Spine Surgery at the Wrong Level: Part I, Cervical Spine. Cureus 2020, 12, e8667. [Google Scholar] [CrossRef]
- Shah, M.; Halalmeh, D.R.; Sandio, A.; Tubbs, R.S.; Moisi, M.D. Anatomical Variations That Can Lead to Spine Surgery at the Wrong Level: Part II Thoracic Spine. Cureus 2020, 12, e8684. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.; Halalmeh, D.R.; Sandio, A.; Tubbs, R.S.; Moisi, M.D. Anatomical Variations That Can Lead to Spine Surgery at the Wrong Level: Part III Lumbosacral Spine. Cureus 2020, 12, e9433. [Google Scholar] [CrossRef]
- Asher, R.J.; Lin, K.H.; Kardjilov, N.; Hautier, L. Variability and constraint in the mammalian vertebral column. J. Evol. Biol. 2011, 24, 1080–1090. [Google Scholar] [CrossRef]
- Brocal, J.; De Decker, S.; José-López, R.; Guevar, J.; Ortega, M.; Parkin, T.; Ter Haar, G.; Gutierrez-Quintana, R. Evaluation of radiography as a screening method for detection and characterisation of congenital vertebral malformations in dogs. Vet. Rec. 2018, 182, 573. [Google Scholar] [CrossRef] [PubMed]
- Du Plessis, A.M.; Greyling, L.M.; Page, B.J. Differentiation and classification of thoracolumbar transitional vertebrae. J. Anat. 2018, 232, 850–856. [Google Scholar] [CrossRef] [PubMed]
- Galis, F.; Carrier, D.R.; van Alphen, J.; van der Mije, S.D.; Van Dooren, T.J.; Metz, J.A.; ten Broek, C.M. Fast running restricts evolutionary change of the vertebral column in mammals. Proc. Natl. Acad. Sci. USA 2014, 111, 11401–11406. [Google Scholar] [CrossRef] [PubMed]
- Williams, S.A.; Spear, J.K.; Petrullo, L.; Goldstein, D.M.; Lee, A.B.; Peterson, A.L.; Miano, D.A.; Kaczmarek, E.B.; Shattuck, M.R. Increased variation in numbers of presacral vertebrae in suspensory mammals. Nat. Ecol. Evol. 2019, 3, 949–956. [Google Scholar] [CrossRef]
- Kumary, S.U.; Moorthy, O.R.; Kannekanti, R.; Ramesh, G. Gross Anatomical Observations on the Sacrum of Guinea Pig (Cavia porcellus). Int. J. Livest. Res. 2020, 2, 67–72. [Google Scholar] [CrossRef]
- Mallo, M. The vertebrate tail: A gene playground for evolution. Cell. Mol. Life Sci. 2020, 77, 1021–1030. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, T.; Thorington, R.W., Jr.; Bohaska, P.W.; Sato, F. Variability and constraint of vertebral formulae and proportions in colugos, tree shrews, and rodents, with special reference to vertebral modification by aerodynamic adaptation. Folia Morphol. 2018, 77, 44–56. [Google Scholar] [CrossRef]
- Inaba, T.; Wakisaka, Y. Congenital malformation of the skeleton in Weiser-Maples guinea pigs. Jikken Dobutsu 1992, 41, 189–201. [Google Scholar] [CrossRef][Green Version]
- Spadliński, Ł.; Cecot, T.; Majos, A.; Stefańczyk, L.; Pietruszewska, W.; Wysiadecki, G.; Topol, M.; Polguj, M. The Epidemiological, Morphological, and Clinical Aspects of the Cervical Ribs in Humans. BioMed Res. Int. 2016, 2016, 8034613. [Google Scholar] [CrossRef]
- van der Geer, A.A.E.; Galis, F. High incidence of cervical ribs indicates vulnerable condition in Late Pleistocene woolly rhinoceroses. PeerJ 2017, 5, e3684. [Google Scholar] [CrossRef]
- Reumer, J.W.; Ten Broek, C.M.; Galis, F. Extraordinary incidence of cervical ribs indicates vulnerable condition in Late Pleistocene mammoths. PeerJ 2014, 2, e318. [Google Scholar] [CrossRef][Green Version]




{kind=link}
{kind=link}
{kind=link}
| Vertebral Column | Total | Female | Male | Sacral Bone Morphology | Number of Caudal Vertebrae | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| F | S | S1 | Cd5 | Cd6 | Cd7 | |||||
| Presacral vertebral column | Sacrocaudal vertebral column | n (%) | n (%) | n (%) | n (%) | n (%) | n (%) | n (%) | n (%) | n (%) |
| C7/Th13/L6 | S4/Cd5-7 | 180 (75) | 112 (46.67) | 68 (28.33) | 102 (42.5) | 48 (20) | 30 (12.5) | 8 (3.33) | 147 (61.25) | 25 (10.42) |
| S3/Cd6-7 | 10 (4.17) | 7 (2.92) | 3 (1.25) | 6 (2.5) | 4 (1.67) | 0 (0) | 0 (0) | 4 (1.67) | 6 (2.5) | |
| S5/Cd5-6 | 4 (1.67) | 1 (0.42) | 3 (1.25) | 1 (0.42) | 0 (0) | 3 (1.25) | 1 (0.42) | 3 (1.25) | 0 (0) | |
| C7/Th13/L7 | S4/Cd6 | 3 (1.25) | 1 (0.42) | 2 (0.83) | 2 (0.83) | 0 (0) | 1 (0.42) | 1 (0.42) | 2 (0.83) | 0 (0) |
| S3/Cd6-7 | 2 (0.83) | 0 (0) | 2 (0.83) | 2 (0.83) | 0 (0) | 0 (0) | 0 (0) | 1 (0.42) | 1 (0.42) | |
| C7/Th13/L5 | S4/Cd6-7 | 6 (2.5) | 4 (1.67) | 2 (0.83) | 3 (1.25) | 3 (1.25) | 0 (0) | 0 (0) | 2 (0.83) | 4 (1.67) |
| S5/Cd7 | 1 (0.42) | 0 (0) | 1 (0.42) | 0 (0) | 0 (0) | 1 (0.42) | 0 (0) | 0 (0) | 1 (0.42) | |
| C7/Th12/L6 | S4/Cd6 | 3 (1.25) | 2 (0.83) | 1 (0.42) | 3 (1.25) | 0 (0) | 0 (0) | 0 (0) | 3 (1.25) | 0 (0) |
| C7/Th12/L7 | S4/Cd5 | 1 (0.42) | 1 (0.42) | 0 (0) | 1 (0.42) | 0 (0) | 0 (0) | 1 (0.42) | 0 (0) | 0 (0) |
| Without congenital abnormalities | 210 (87.5) | 128 (53.33) | 82 (34.17) | 120 (50) | 55 (22.92) | 35 (14.59) | 11 (4.58) | 162 (67.5) | 37 (15.42) | |
| Congenital abnormalities | 30 (12.5) | 20 (8.33) | 10 (4.17) | 17 (7.08) | 8 (3.33) | 5 (2.08) | 4 (1.67) | 17 (7.08) | 9 (3.75) | |
| Total | 240 (100) | 148 (61.66) | 92 (38.34) | 137 (57.08) | 63 (26.25) | 40 (16.67) | 15 (6.25) | 179 (74.58) | 46 (19.17) | |
| Type of Pathology | Morphology | Localization | Vertebral Formula | n | Population (%) | Vertebral Pathology (%) | Gender |
|---|---|---|---|---|---|---|---|
| Single pathology | |||||||
| CTV | Unilateral rudimentary rib | CT junction | C7+CTV/Th12/L6/ S4/Cd6 | 1 | 0.42 | 3.33 | M |
| TTV | Bilateral incomplete ossification of rib | TL junction | C7/Th13+TTV/L6/S3/Cd7 | 1 | 2.5 | 20 | F |
| TL junction | C7/Th12+TTV/L6/S3/Cd7 | 1 | F | ||||
| TL junction | C7/Th12+TTV/L5/S4/Cd6 | 1 | M | ||||
| TL junction | C7/Th12+TTV/L6/S4/Cd6 | 1 | F | ||||
| TL junction | C7/Th12+TTV/L6/S5/Cd6 | 1 | F | ||||
| TL junction | C7/Th12+TTV/L5/S4/Cd7 | 1 | M | ||||
| Unilateral incomplete ossification of rib | TL junction | C7/Th12+TTV/L5/S4/Cd7 | 2 | 3.33 | 26.67 | F | |
| TL junction | C7/Th12+TTV/L6/S4/Cd6 | 1 | M | ||||
| TL junction | C7/Th12+TTV/L6/S4/Cd6 | 1 | F | ||||
| TL junction | C7/Th12+TTV/L5/S4/Cd6 | 1 | F | ||||
| TL junction | C7/Th12+TTV/L6/S4/Cd7 | 1 | F | ||||
| TL junction | C7/Th13+TTV/L5/S4/Cd6 | 1 | F | ||||
| TL junction | C7/Th12+TTV/L5/S4/Cd6 | 1 | M | ||||
| Bilateral rudimentary rib | TL junction | C7/Th12+TTV/L5/S4/Cd7 | 1 | 1.25 | 10 | F | |
| TL junction | C7/Th12+TTV/L5/S4/Cd7 | 1 | M | ||||
| TL junction | C7/Th12+TTV/L6/S5/Cd6 | 1 | F | ||||
| Unilateral rudimentary rib | TL junction | C7/Th12+TTV/L6/S3/Cd7 | 1 | 1.67 | 13.33 | F | |
| TL junction | C7/Th12+TTV/L6/S4/Cd6 | 1 | F | ||||
| TL junction | C7/Th12+TTV/L6/S4/Cd5 | 1 | M | ||||
| TL junction | C7/Th12+TTV/L5/S4/Cd6 | 1 | M | ||||
| Unilateral rudimentary rib and contralateral incomplete ossification of rib | TL junction | C7/Th12+TTV/L6/S4/Cd6 | 1 | 0.42 | 3.33 | F | |
| LTV | Asymmetrical | LS junction | C7/Th13/L5+LTV/S4/Cd6 | 1 | 2.08 | 16.67 | F |
| LS junction | C7/Th13/L5+LTV/S3/Cd6 | 1 | M | ||||
| LS junction | C7/Th13/L5+LTV/S4/Cd5 | 1 | F | ||||
| LS junction | C7/Th13/L5+LTV/S4/Cd6 | 1 | F | ||||
| LS junction | C7/Th13/L6+LTV/S3/Cd5 | 1 | M | ||||
| Multiple pathology | |||||||
| Block vertebra, TTV | Block C2-C3, TTV unilateral rudimentary rib | Cervical, TL junction | C8(blockC2-C3)/Th12+TTV/ L5/S4/Cd6 | 1 | 0.42 | 3.33 | F |
| TTV, LTV | Bilateral rudimentary rib, asymmetrical LTV | TL, LS junction | C7/Th12+TTV/L5+LTV/S4/Cd5 | 1 | 0.42 | 3.33 | F |
| Total | 30 | 12.50 | 100 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Proks, P.; Johansen, T.M.; Nývltová, I.; Komenda, D.; Černochová, H.; Vignoli, M. Vertebral Formulae and Congenital Vertebral Anomalies in Guinea Pigs: A Retrospective Radiographic Study. Animals 2021, 11, 589. https://doi.org/10.3390/ani11030589
Proks P, Johansen TM, Nývltová I, Komenda D, Černochová H, Vignoli M. Vertebral Formulae and Congenital Vertebral Anomalies in Guinea Pigs: A Retrospective Radiographic Study. Animals. 2021; 11(3):589. https://doi.org/10.3390/ani11030589
Chicago/Turabian StyleProks, Pavel, Trude Maria Johansen, Ivana Nývltová, Dominik Komenda, Hana Černochová, and Massimo Vignoli. 2021. "Vertebral Formulae and Congenital Vertebral Anomalies in Guinea Pigs: A Retrospective Radiographic Study" Animals 11, no. 3: 589. https://doi.org/10.3390/ani11030589
APA StyleProks, P., Johansen, T. M., Nývltová, I., Komenda, D., Černochová, H., & Vignoli, M. (2021). Vertebral Formulae and Congenital Vertebral Anomalies in Guinea Pigs: A Retrospective Radiographic Study. Animals, 11(3), 589. https://doi.org/10.3390/ani11030589