
Inclusion of Oat and Yeast Culture in Sow Gestational and Lactational Diets Alters Immune and Antimicrobial Associated Proteins in Milk

 ,
,  , and
, and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Housing and Care
2.2. Feed Administration
2.3. Piglet Feed Intake and Weight Measurements
2.4. Fecal Scoring
2.5. Milk Collection
2.6. Milk Immunoglobulin and Proteome Analysis
2.7. Statistical Analysis
3. Results
3.1. Piglet Weights
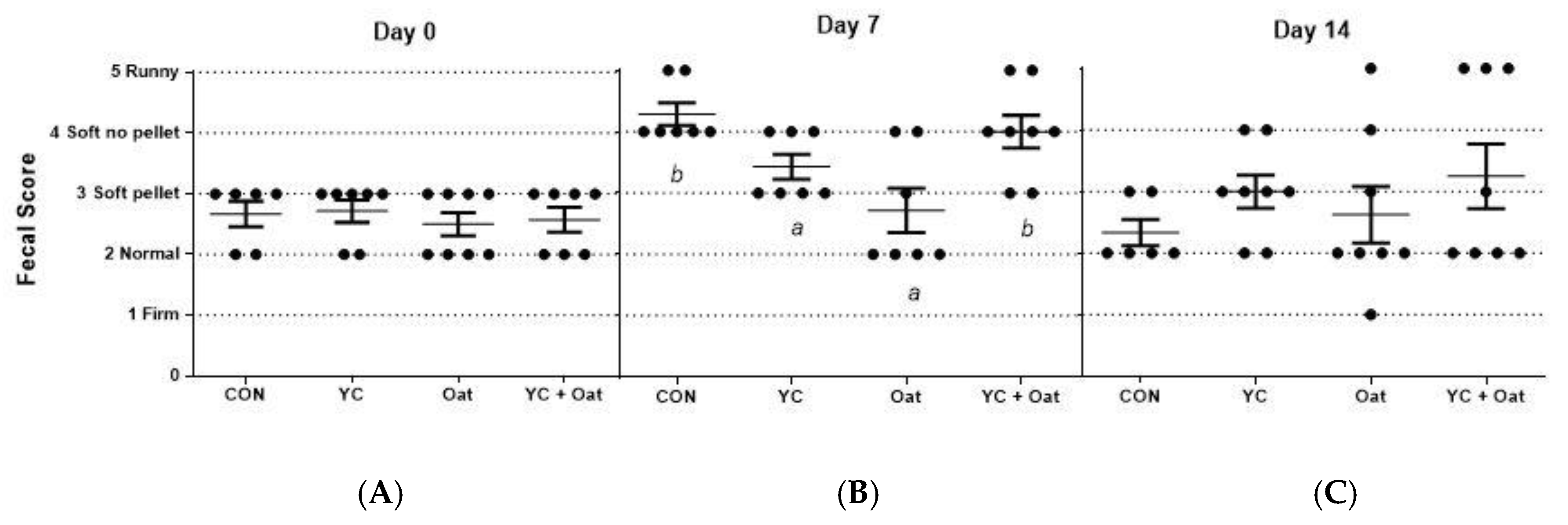
3.2. Post-Weaning Diarrhea Scores
3.3. Milk Immunoglobulins
3.4. Immunoglobulin Reactivity
3.5. Milk Proteome Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- National Pork Board. World per Capita Pork Consumption. Available online: https://www.pork.org/facts/stats/u-s-pork-exports/world-per-capita-pork-consumption/ (accessed on 2 December 2020).
- US Department of Agriculture Foreign Agricultural Service. Livestock and Poultry: World Markets and Trade. Available online: https://apps.fas.usda.gov/psdonline/circulars/livestock_poultry.pdf (accessed on 9 October 2020).
- Koketsu, Y.; Tani, S.; Iida, R. Factors for improving reproductive performance of sows and herd productivity in commercial breeding herds. Porc. Health Manag. 2017, 3, 1–10. [Google Scholar] [CrossRef]
- Fairbrother, J.M.; Éric, N.; Gyles, C.L. Escherichia coli in postweaning diarrhea in pigs: An update on bacterial types, pathogenesis, and prevention strategies. Anim. Health Res. Rev. 2005, 6, 17–39. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, B.; Nyachoti, C.M. Husbandry practices and gut health outcomes in weaned piglets: A review. Anim. Nutr. 2017, 3, 205–211. [Google Scholar] [CrossRef]
- Rhouma, M.; Fairbrother, J.M.; Beaudry, F.; Letellier, A. Post weaning diarrhea in pigs: Risk factors and non-colistin-based control strategies. Acta Vet. Scand. 2017, 59, 1–19. [Google Scholar] [CrossRef]
- Šinkora, M.; Butler, J.E. The ontogeny of the porcine immune system. Dev. Comp. Immunol. 2009, 33, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Pluske, J.R.; Hampson, D.J.; Williams, I.H. Factors influencing the structure and function of the small intestine in the weaned pig: A review. Livest. Prod. Sci. 1997, 51, 215–236. [Google Scholar] [CrossRef]
- Campbell, J.M.; Crenshaw, J.D.; Polo, J. The biological stress of early weaned piglets. J. Anim. Sci. Biotechnol. 2013, 4, 19. [Google Scholar] [CrossRef]
- McLamb, B.L.; Gibson, A.J.; Overman, E.L.; Stahl, C.; Moeser, A.J. Early Weaning Stress in Pigs Impairs Innate Mucosal Immune Responses to Enterotoxigenic E. coli Challenge and Exacerbates Intestinal Injury and Clinical Disease. PLoS ONE 2013, 8, e59838. [Google Scholar] [CrossRef] [PubMed]
- Gimsa, U.; Tuchscherer, M.; Kanitz, E. Psychosocial Stress and Immunity—What Can We Learn From Pig Studies? Front. Behav. Neurosci. 2018, 12, 64. [Google Scholar] [CrossRef] [PubMed]
- Dhabhar, F.S.; McEwen, B.S. Acute Stress Enhances while Chronic Stress Suppresses Cell-Mediated Immunity in Vivo: A Potential Role for Leukocyte Trafficking. Brain Behav. Immun. 1997, 11, 286–306. [Google Scholar] [CrossRef]
- Segerstrom, S.C.; Miller, G.E. Psychological Stress and the Human Immune System: A Meta-Analytic Study of 30 Years of Inquiry. Psychol. Bull. 2004, 130, 601–630. [Google Scholar] [CrossRef] [PubMed]
- Morey, J.N.; A Boggero, I.; Scott, A.B.; Segerstrom, S.C. Current directions in stress and human immune function. Curr. Opin. Psychol. 2015, 5, 13–17. [Google Scholar] [CrossRef]
- Virdi, V.; Coddens, A.; De Buck, S.; Millet, S.; Goddeeris, B.M.; Cox, E.; De Greve, H.; Depicker, A. Orally fed seeds producing designer IgAs protect weaned piglets against enterotoxigenic Escherichia coli infection. Proc. Natl. Acad. Sci. USA 2013, 110, 11809–11814. [Google Scholar] [CrossRef] [PubMed]
- Gresse, R.; Chaucheyras-Durand, F.; Fleury, M.A.; Van de Wiele, T.; Forano, E.; Blanquet-Diot, S. Gut Microbiota Dysbiosis in Postweaning Piglets: Understanding the Keys to Health. Trends Microbiol. 2017, 25, 851–873. [Google Scholar] [CrossRef]
- Guevarra, R.B.; Lee, J.H.; Lee, S.H.; Seok, M.-J.; Kim, D.W.; Na Kang, B.; Johnson, T.J.; Isaacson, R.E.; Kim, H.B. Piglet gut microbial shifts early in life: Causes and effects. J. Anim. Sci. Biotechnol. 2019, 10, 1–10. [Google Scholar] [CrossRef]
- Theil, P.; Hurley, W. The Protein Component of Sow Colostrum and Milk. In Milk Proteins—From Structure to Biological Properties and Health Aspects; Intech Open: London, UK, 2016; p. 10577262841. [Google Scholar]
- Ballard, O.; Morrow, A.L. Human Milk Composition. Pediatr. Clin. North Am. 2013, 60, 49–74. [Google Scholar] [CrossRef] [PubMed]
- Hurley, W.; Farmer, C. Composition of sow colostrum and milk. In The Gestating and Lactating Sow; Wageningen Academic Publishers: Wageningen, The Netherlands, 2015; pp. 193–230. [Google Scholar]
- Salmon, H.; Berri, M.; Gerdts, V.; Meurens, F. Humoral and cellular factors of maternal immunity in swine. Dev. Comp. Immunol. 2009, 33, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Bandrick, M.; Ariza-Nieto, C.; Baidoo, S.K.; Molitor, T.W. Colostral antibody-mediated and cell-mediated immunity contributes to innate and antigen-specific immunity in piglets. Dev. Comp. Immunol. 2014, 43, 114–120. [Google Scholar] [CrossRef]
- Nechvatalova, K.; Kudlackova, H.; Leva, L.; Babickova, K.; Faldyna, M. Transfer of humoral and cell-mediated immunity via colostrum in pigs. Vet. Immunol. Immunopathol. 2011, 142, 95–100. [Google Scholar] [CrossRef]
- Nguyen, T.V.; Yuan, L.; Azevedo, M.S.; Jeong, K.-I.; Gonzalez, A.-M.; Saif, L.J. Transfer of maternal cytokines to suckling piglets: In vivo and in vitro models with implications for immunomodulation of neonatal immunity. Vet. Immunol. Immunopathol. 2007, 117, 236–248. [Google Scholar] [CrossRef]
- Oddy, W.H.; McMahon, R.J. Milk-derived or recombinant transforming growth factor-beta has effects on immunological outcomes: A review of evidence from animal experimental studies. Clin. Exp. Allergy 2011, 41, 783–793. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Chen, F.; Zhang, Y.; Lv, Y.; Heng, J.; Min, T.; Li, L.; Guan, W. Recent progress of porcine milk components and mammary gland function. J. Anim. Sci. Biotechnol. 2018, 9, 1–13. [Google Scholar] [CrossRef]
- Neville, M.C.; Anderson, S.M.; McManaman, J.L.; Badger, T.M.; Bunik, M.; Contractor, N.; Crume, T.; Dabelea, D.; Donovan, S.M.; Forman, N.; et al. Lactation and Neonatal Nutrition: Defining and Refining the Critical Questions. J. Mammary Gland. Biol. Neoplasia 2012, 17, 167–188. [Google Scholar] [CrossRef] [PubMed]
- Berding, K.; Wang, M.; Monaco, M.H.; Alexander, L.S.; Mudd, A.T.; Chichlowski, M.; Waworuntu, R.V.; Berg, B.M.; Miller, M.J.; Dilger, R.N.; et al. Prebiotics and Bioactive Milk Fractions Affect Gut Development, Microbiota, and Neurotransmitter Expression in Piglets. J. Pediatr. Gastroenterol. Nutr. 2016, 63, 688–697. [Google Scholar] [CrossRef] [PubMed]
- Gregory, K.E.; Samuel, B.S.; Houghteling, P.; Shan, G.; Ausubel, F.M.; Sadreyev, R.I.; Walker, W.A. Influence of maternal breast milk ingestion on acquisition of the intestinal microbiome in preterm infants. Microbiome 2016, 4, 1–15. [Google Scholar] [CrossRef]
- Salcedo, J.; Frese, S.A.; Mills, D.A.; Barile, D. Characterization of porcine milk oligosaccharides during early lactation and their relation to the fecal microbiome. J. Dairy Sci. 2016, 99, 7733–7743. [Google Scholar] [CrossRef]
- Le Doare, K.; Holder, B.; Bassett, A.; Pannaraj, P.S. Mother’s Milk: A Purposeful Contribution to the Development of the Infant Microbiota and Immunity. Front. Immunol. 2018, 9, 361. [Google Scholar] [CrossRef] [PubMed]
- Bravi, F.; Wiens, F.; DeCarli, A.; Pont, A.D.; Agostoni, C.; Ferraroni, M. Impact of maternal nutrition on breast-milk composition: A systematic review. Am. J. Clin. Nutr. 2016, 104, 646–662. [Google Scholar] [CrossRef]
- Padilha, M.; Danneskiold-Samsøe, N.B.; Brejnrod, A.; Hoffmann, C.; Cabral, V.P.; Iaucci, J.D.M.; Sales, C.H.; Fisberg, R.M.; Cortez, R.V.; Brix, S.; et al. The Human Milk Microbiota is Modulated by Maternal Diet. Microorganisms 2019, 7, 502. [Google Scholar] [CrossRef] [PubMed]
- Leonard, S.G.; Sweeney, T.; Bahar, B.; Lynch, B.P.; O’Doherty, J.V. Effect of maternal fish oil and seaweed extract supplementation on colostrum and milk composition, humoral immune response, and performance of suckled piglets1. J. Anim. Sci. 2010, 88, 2988–2997. [Google Scholar] [CrossRef]
- Jeurink, P.V.; Knipping, K.; Wiens, F.; Barańska, K.; Stahl, B.; Garssen, J.; Krolak-Olejnik, B. Importance of maternal diet in the training of the infant’s immune system during gestation and lactation. Crit. Rev. Food Sci. Nutr. 2019, 59, 1311–1319. [Google Scholar] [CrossRef]
- Chu, D.M.; Meyer, K.M.; Prince, A.L.; Aagaard, K.M. Impact of maternal nutrition in pregnancy and lactation on offspring gut microbial composition and function. Gut Microbes 2016, 7, 459–470. [Google Scholar] [CrossRef] [PubMed]
- Val-Laillet, D.; Besson, M.; Guérin, S.; Coquery, N.; Randuineau, G.; Kanzari, A.; Quesnel, H.; Bonhomme, N.; Bolhuis, J.E.; Kemp, B.; et al. A maternal Western diet during gestation and lactation modifies offspring’s microbiota activity, blood lipid levels, cognitive responses, and hippocampal neurogenesis in Yucatan pigs. FASEB J. 2017, 31, 2037–2049. [Google Scholar] [CrossRef] [PubMed]
- Laws, J.; Juniper, D.T.; Lean, I.J.; Amusquivar, E.; Herrera, E.; Dodds, P.F.; Clarke, L. Supplementing sow diets with palm oil during late gestation and lactation: Effects on milk production, sow hormonal profiles and growth and development of her offspring. Animal 2018, 12, 2578–2586. [Google Scholar] [CrossRef]
- Wang, C.; Lin, C.; Su, W.; Zhang, Y.; Wang, F.; Wang, Y.; Shi, C.; Lu, Z. Effects of supplementing sow diets with fermented corn and soybean meal mixed feed during lactation on the performance of sows and progeny. J. Anim. Sci 2018. [Google Scholar] [CrossRef]
- Lee, S.D.; Kim, J.H.; Jung, H.J.; Kim, Y.H.; Kim, I.C.; Kim, S.B.; Lim, S.Y.; Jung, W.S.; Lee, S.-H.; Kim, Y.J. The effect of ginger extracts on the antioxidant capacity and IgG concentrations in the colostrum and plasma of neo-born piglets and sows. Livest. Sci. 2013, 154, 117–122. [Google Scholar] [CrossRef]
- Leonard, S.G.; Sweeney, T.; Bahar, B.; Lynch, B.P.; O’Doherty, J.V. Effect of dietary seaweed extracts and fish oil supplementation in sows on performance, intestinal microflora, intestinal morphology, volatile fatty acid concentrations and immune status of weaned pigs. Br. J. Nutr. 2010, 105, 549–560. [Google Scholar] [CrossRef] [PubMed]
- Quesnel, H.; Farmer, C. Review: Nutritional and endocrine control of colostrogenesis in swine. Animal 2019, 13, s26–s34. [Google Scholar] [CrossRef]
- Song, M.; Baidoo, S.K.; Shurson, G.C.; Whitney, M.H.; Johnston, L.J.; Gallaher, D. Dietary effects of distillers dried grains with solubles on performance and milk composition of lactating sows1. J. Anim. Sci. 2010, 88, 3313–3319. [Google Scholar] [CrossRef]
- Alexopoulos, C.; Georgoulakis, I.E.; Tzivara, A.; Kritas, S.K.; Siochu, A.; Kyriakis, S.C. Field evaluation of the efficacy of a probiotic containing Bacillus licheniformis and Bacillus subtilis spores, on the health status and performance of sows and their litters. J. Anim. Physiol. Anim. Nutr. 2004, 88, 381–392. [Google Scholar] [CrossRef]
- Shen, Y.B.; Carroll, J.A.; Yoon, I.; Mateo, R.D.; Kim, S.W. Effects of supplementing Saccharomyces cerevisiae fermentation product in sow diets on performance of sows and nursing piglets1,2. J. Anim. Sci. 2011, 89, 2462–2471. [Google Scholar] [CrossRef]
- Jang, Y.; Kang, K.; Piao, L.; Jeong, T.; Auclair, E.; Jonvel, S.; D’Inca, R.; Kim, Y.Y. Effects of live yeast supplementation to gestation and lactation diets on reproductive performance, immunological parameters and milk composition in sows. Livest. Sci. 2013, 152, 167–173. [Google Scholar] [CrossRef]
- Zanello, G.; Meurens, F.; Serreau, D.; Chevaleyre, C.; Melo, S.; Berri, M.; D’Inca, R.; Auclair, E.; Salmon, H. Effects of dietary yeast strains on immunoglobulin in colostrum and milk of sows. Vet. Immunol. Immunopathol. 2013, 152, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Jurgens, M.H.; A Rikabi, R.; Zimmerman, D.R. The effect of dietary active dry yeast supplement on performance of sows during gestation-lactation and their pigs. J. Anim. Sci. 1997, 75, 593–597. [Google Scholar] [CrossRef] [PubMed]
- Szuba-Trznadel, A.; Rząsa, A.; Lira, R.; Fuchs, B. The influence of (1,3)-(1,6)-β-D-glucan on the production results of sows and their offspring. J. Anim. Feed. Sci. 2014, 23, 228–235. [Google Scholar] [CrossRef][Green Version]
- Lu, H.; Wilcock, P.; Adeola, O.; Ajuwon, K.M. Effect of live yeast supplementation to gestating sows and nursery piglets on postweaning growth performance and nutrient digestibility. J. Anim. Sci. 2019, 97, 2534–2540. [Google Scholar] [CrossRef]
- Mroz, Z.; Partridge, I.G.; Mitchell, G.; Keal, H.D. The effect of oat hulls, added to the basal ration for pregnant sows, on reproductive performance, apparent digestibility, rate of passage and plasma parameters. J. Sci. Food Agric. 1986, 37, 239–247. [Google Scholar] [CrossRef]
- Matte, J.J.; Robert, S.; Girard, C.L.; Farmer, C.; Martineau, G.-P. Effect of bulky diets based on wheat bran or oat hulls on reproductive performance of sows during their first two parities1. J. Anim. Sci. 1994, 72, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Gyawali, R.; Minor, R.C.; Donovan, B.; Ibrahim, S.A. Inclusion of Oat in Feeding Can Increase the Potential Probiotic Bifidobacteria in Sow Milk. Animal 2015, 5, 610–623. [Google Scholar] [CrossRef]
- Stier, H.; Ebbeskotte, V.; Gruenwald, J. Immune-modulatory effects of dietary Yeast Beta-1,3/1,6-D-glucan. Nutr. J. 2014, 13, 38. [Google Scholar] [CrossRef]
- Estrada, A.; Van Kessel, A.; Laarveld, B. Effect of administration of oat beta-glucan on immune parameters of healthy and immunosuppressed beef steers. Can. J. Vet. Res. 1999, 63, 261–268. [Google Scholar]
- Council, N.R. Nutrient Requirements of Swine, 11th ed.; The National Academies Press: Washington, DC, USA, 2012; p. 420. [Google Scholar]
- Harlow, K.; Suarez-Trujillo, A.; Hedrick, V.; Sobreira, T.; Aryal, U.K.; Stewart, K.; Casey, T. Temporal analysis of vaginal proteome reveals developmental changes in lower reproductive tract of gilts across the first two weeks postnatal. Sci. Rep. 2019, 9, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Harlow, K.; Taylor, E.; Casey, T.; Hedrick, V.; Sobreira, T.; Aryal, U.K.; LeMenager, R.P.; Funnell, B.; Stewart, K. Diet Impacts Pre-implantation Histotroph Proteomes in Beef Cattle. J. Proteome Res. 2018, 17, 2144–2155. [Google Scholar] [CrossRef]
- Aryal, U.K.; McBride, Z.; Chen, D.; Xie, J.; Szymanski, D.B. Analysis of protein complexes in Arabidopsis leaves using size exclusion chromatography and label-free protein correlation profiling. J. Proteom. 2017, 166, 8–18. [Google Scholar] [CrossRef]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 12, 1367–1372. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Stephens, R.; Baseler, M.W.; Lane, H.C.; Lempicki, R.A. DAVID gene ID conversion tool. Bioinformation 2008, 2, 428–430. [Google Scholar] [CrossRef]
- Huang da, W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Stelzer, G.; Rosen, N.; Plaschkes, I.; Zimmerman, S.; Twik, M.; Fishilevich, S.; Stein, T.I.; Nudel, R.; Lieder, I.; Mazor, Y.; et al. The GeneCards Suite: From Gene Data Mining to Disease Genome Sequence Analyses. Curr. Protoc. Bioinform. 2016, 54, 1.30.1–1.30.33. [Google Scholar] [CrossRef] [PubMed]
- Apweiler, R.; Bairoch, A.; Wu, C.H. Protein sequence databases. Curr. Opin. Chem. Biol. 2004, 8, 76–80. [Google Scholar] [CrossRef]
- Tuohy, K.M.; Probert, H.M.; Smejkal, C.W.; Gibson, G.R. Using probiotics and prebiotics to improve gut health. Drug Discov. 2003, 8, 692–700. [Google Scholar] [CrossRef]
- Jarrett, S.; Ashworth, C.J. The role of dietary fibre in pig production, with a particular emphasis on reproduction. J. Anim. Sci. Biotechnol. 2018, 9, 1–11. [Google Scholar] [CrossRef]
- Kim, S.W.; Brandherm, M.; Freeland, M.; Newton, B.; Cook, D.; Yoon, I. Effects of Yeast Culture Supplementation to Gestation and Lactation Diets on Growth of Nursing Piglets. Asian-Australas. J. Anim. Sci. 2008, 21, 1011–1014. [Google Scholar] [CrossRef]
- Kim, S.W.; Brandherm, M.; Newton, B.; Cook, D.R.; Yoon, I.; Fitzner, G. Effect of supplementing Saccharomyces cerevisiae fermentation product in sow diets on reproductive performance in a commercial environment. Can. J. Anim. Sci. 2010, 90, 229–232. [Google Scholar] [CrossRef]
- Ekern, A.; Havrevoll, Ø.; Haug, A.; Berg, J.; Lindstad, P.; Skeie, S. Oat and Barley Based Concentrate Supplements for Dairy Cows. Acta Agric. Scand. Sect. A—Anim. Sci. 2003, 53, 65–73. [Google Scholar] [CrossRef]
- Kalmus, P.; Orro, T.; Waldmann, A.; Lindjärv, R.; Kask, K. Effect of yeast culture on milk production and metabolic and reproductive performance of early lactation dairy cows. Acta Vet. Scand. 2009, 51, 32. [Google Scholar] [CrossRef] [PubMed]
- Poppy, G.; Rabiee, A.; Lean, I.; Sanchez, W.; Dorton, K.; Morley, P. A meta-analysis of the effects of feeding yeast culture produced by anaerobic fermentation of Saccharomyces cerevisiae on milk production of lactating dairy cows. J. Dairy Sci. 2012, 95, 6027–6041. [Google Scholar] [CrossRef]
- Hasan, S.; Junnikkala, S.; Peltoniemi, O.; Paulin, L.; Lyyski, A.; Vuorenmaa, J.; Oliviero, C. Dietary supplementation with yeast hydrolysate in pregnancy influences colostrum yield and gut microbiota of sows and piglets after birth. PLoS ONE 2018, 13, e0197586. [Google Scholar] [CrossRef]
- Li, J.; Xing, J.; Li, D.; Wang, X.; Zhao, L.; Lv, S.; Huang, D. Effects of β-glucan extracted fromSaccharomyces cerevisiaeon humoral and cellular immunity in weaned piglets. Arch. Anim. Nutr. 2005, 59, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Vetvicka, V.; Oliveira, C. β(1-3)(1-6)-D-glucans modulate immune status in pigs: Potential importance for efficiency of commercial farming. Ann. Transl. Med. 2014, 2, 16. [Google Scholar]
- Ushida, K.; Kameue, C.; Tsukahara, T.; Fukuta, K.; Nakanishi, N. Decreasing Traits of Fecal Immunoglobulin A in Neonatal and Weaning Piglets. J. Vet. Med Sci. 2008, 70, 849–852. [Google Scholar] [CrossRef]
- Kafkova, L.R.; Brokesova, D.; Krupka, M.; Stehlikova, Z.; Dvorak, J.; Coufal, S.; Fajstova, A.; Srutkova, D.; Stepanova, K.; Hermanova, P.; et al. Secretory IgA N-glycans contribute to the protection against E. coli O55 infection of germ-free piglets. Mucosal Immunol. 2020, 1–12. [Google Scholar] [CrossRef]
- Castro-Dopico, T.; Clatworthy, M.R. IgG and Fcγ Receptors in Intestinal Immunity and Inflammation. Front. Immunol. 2019, 10, 805. [Google Scholar] [CrossRef] [PubMed]
- Niewiesk, S. Maternal Antibodies: Clinical Significance, Mechanism of Interference with Immune Responses, and Possible Vaccination Strategies. Front. Immunol. 2014, 5, 446. [Google Scholar] [CrossRef] [PubMed]
- Werner, C.; Schubbert, A.; Schrödl, W.; Krüger, M.; Sundrum, A. Effects of feeding different roughage components to sows in gestation on bacteriological and immunological parameters in colostrum and immune response of piglets. Arch. Anim. Nutr. 2014, 68, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, S.; Tsukahara, T.; Nishibayashi, R.; Nakatani, M.; Okutani, M.; Nakanishi, N.; Ushida, K.; Inoue, R. Shotgun proteomic analysis of porcine colostrum and mature milk. Anim. Sci. J. 2014, 85, 440–448. [Google Scholar] [CrossRef]
- Dallas, D.C.; Murray, N.M.; Gan, J. Proteolytic Systems in Milk: Perspectives on the Evolutionary Function within the Mammary Gland and the Infant. J. Mammary Gland. Biol. Neoplasia 2015, 20, 133–147. [Google Scholar] [CrossRef] [PubMed]
- Saxena, A.; Belinskaya, T.; Schopfer, L.M.; Lockridge, O. Tetramer organizing polyproline-rich peptides identified by mass spectrometry after release of the peptides from Hupresin-purified butyrylcholinesterase tetramers isolated from milk of domestic pig (Sus scrofa). Data Brief 2018, 20, 1607–1619. [Google Scholar] [CrossRef]
- Legrand, D. Lactoferrin, a key molecule in immune and inflammatory processes1This article is part of Special Issue entitled Lactoferrin and has undergone the Journal’s usual peer review process. Biochem. Cell Biol. 2012, 90, 252–268. [Google Scholar] [CrossRef] [PubMed]
- Perdijk, O.; Van Neerven, R.J.J.; Brink, E.V.D.; Savelkoul, H.F.J.; Brugman, S. Bovine Lactoferrin Modulates Dendritic Cell Differentiation and Function. Nutrients 2018, 10, 848. [Google Scholar] [CrossRef]
- Pammi, M.; Suresh, G. Enteral lactoferrin supplementation for prevention of sepsis and necrotizing enterocolitis in preterm infants. Cochrane Database Syst. Rev. 2017, 6, CD007137. [Google Scholar] [CrossRef]
- Gao, S.; Yang, W.; Yu, H.; Liu, R.; Dong, Z.; Zhang, H.; Xia, C.; Xu, C. Significant changes in ITIH4, AHSG, ORM1, and CD46 content in milk fat globule membrane proteins of ketotic dairy cows. J. Dairy Res. 2017, 84, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Wathes, D.C.; Clempson, A.M.; Pollott, G.E. Associations between lipid metabolism and fertility in the dairy cow. Reprod. Fertil. Dev. 2013, 25, 48–61. [Google Scholar] [CrossRef]
- Ochieng, J.; Nangami, G.; Sakwe, A.; Moye, C.; Alvarez, J.; Whalen, D.; Thomas, P.; Lammers, P. Impact of Fetuin-A (AHSG) on Tumor Progression and Type 2 Diabetes. Int. J. Mol. Sci. 2018, 19, 2211. [Google Scholar] [CrossRef] [PubMed]
- Javed, S.; Ahmad, M.; Ahmad, M.M.; Abdin, M.Z.; Hamid, R.; Khan, M.A.; Musarrat, J. Chitinases: An update. J. Pharm. Bioallied Sci. 2013, 5, 21–29. [Google Scholar] [CrossRef] [PubMed]
- USDA-NASS. Crops and Plants; USDA-NASS: Washington, DC, USA, 2012. Available online: http://www.nass.usda.gov/ (accessed on 6 May 2013).




{kind=link}
{kind=link}
{kind=link}
| Ingredient (%) | Control | YC | Oat | YC + Oat |
|---|---|---|---|---|
| Corn | 81.30 | 80.90 | 66.33 | 66.00 |
| Soybean Meal 48% | 13.85 | 13.78 | 13.84 | 13.77 |
| Limestone | 1.11 | 1.11 | 1.11 | 1.11 |
| MON-CAL 21% P | 2.05 | 2.04 | 2.05 | 2.04 |
| Salt | 0.50 | 0.50 | 0.50 | 0.50 |
| Vitamin Premix | 0.15 | 0.15 | 0.15 | 0.15 |
| Sow Vitamin Premix | 0.04 | 0.04 | 0.04 | 0.04 |
| Poultry Fat | 1.00 | 1.00 | 1.00 | 0.99 |
| Yeast Culture 1 | - | 0.50 | - | 0.50 |
| Ground Whole Oat 2 | - | - | 14.99 | 14.91 |
| Total Calculated composition 3 | 100 | 100 | 100 | 100 |
| ME, kcal/kg | 3300 | 3303 | 3174 | 3177 |
| Crude Protein, % | 13.3 | 13.4 | 13.7 | 13.9 |
| Dry Matter, % | 88.0 | 88.1 | 88.3 | 88.3 |
| Starch, % | 51.1 | 50.9 | 47.6 | 47.4 |
| Amino acids | ||||
| Arginine, % | 0.78 | 0.78 | 0.83 | 0.84 |
| Histamine, % | 0.37 | 0.37 | 0.37 | 0.37 |
| Isoleucine, % | 0.52 | 0.53 | 0.54 | 0.55 |
| Leucine, % | 1.28 | 1.28 | 1.26 | 1.26 |
| Lysine, % | 0.61 | 0.62 | 0.65 | 0.66 |
| Methionine, % | 0.24 | 0.24 | 0.31 | 0.32 |
| Cystine, % | 0.25 | 0.25 | 0.28 | 0.28 |
| Tryptophan, % | 0.14 | 0.14 | 0.15 | 0.15 |
| Valine, % | 0.62 | 0.62 | 0.66 | 0.66 |
| Feed Ingredients (%) | Control | YC | Oat | YC + Oat |
|---|---|---|---|---|
| Corn | 74.06 | 74.06 | 65.91 | 65.91 |
| Soybean Meal 48% | 17.62 | 17.62 | 10.86 | 10.86 |
| Limestone Fine | 1.08 | 1.08 | 1.08 | 1.08 |
| MON-CAL 21% P | 2.39 | 2.39 | 2.39 | 2.39 |
| L-LYS 78% | 0.16 | 0.16 | 0.16 | 0.16 |
| Salt | 0.50 | 0.50 | 0.50 | 0.50 |
| Vitamin Premix | 0.15 | 0.15 | 0.15 | 0.15 |
| Sow Vitamin Premix | 0.04 | 0.04 | 0.04 | 0.04 |
| L-Threonine | 0.01 | 0.01 | 0.01 | 0.01 |
| Poultry Fat | 4.00 | 4.00 | 4.00 | 4.00 |
| Yeast Culture 1 | - | 0.50 | - | 0.50 |
| Ground Whole Oat 2 | - | - | 14.97 | 14.97 |
| Total | 100 | 100 | 100 | 100 |
| Calculated composition 3 | ||||
| ME, kcal/kg | 3436 | 3439 | 3317 | 3320 |
| Crude Protein, % | 14.7 | 14.8 | 12.4 | 12.6 |
| Dry Matter, % | 87.1 | 87.1 | 87.2 | 87.2 |
| Starch, % | 46.7 | 46.7 | 47.2 | 47.0 |
| Amino acids | ||||
| Arginine, % | 0.88 | 0.88 | 0.73 | 0.73 |
| Histamine, % | 0.40 | 0.40 | 0.33 | 0.33 |
| Isoleucine, % | 0.58 | 0.59 | 0.48 | 0.48 |
| Leucine, % | 1.35 | 1.35 | 1.14 | 1.15 |
| Lysine, % | 0.83 | 0.84 | 0.69 | 0.69 |
| Methionine, % | 0.25 | 0.25 | 0.29 | 0.29 |
| Cystine, % | 0.26 | 0.26 | 0.25 | 0.25 |
| Tryptophan, % | 0.16 | 0.16 | 0.13 | 0.13 |
| Valine, % | 0.67 | 0.68 | 0.59 | 0.59 |
| PIGLET GROWER RATION | ||
|---|---|---|
| Feed Ingredient | Amount [kg] | % Inclusion |
| Corn meal | 470.01 | 51.9 |
| Wheat middlings | 153.68 | 16.96 |
| Hominy | 136.08 | 15.0 |
| HI-pro soybean meal | 116.57 | 12.9 |
| Ground limestone | 10.89 | 1.2 |
| Dist. dried grains | 7.48 | 0.82 |
| Phosphate mono/d | 5.22 | 0.57 |
| Salt (plain) | 3.18 | 0.35 |
| Lysine (78.8%) | 0.91 | 0.1 |
| Swine premix 2 | 0.91 | 0.1 |
| Selenium premix (0.06%) | 0.45 | 0.05 |
| Feed preservative (65%) | 0.45 | 0.05 |
| TOTAL | 905.82 | 100 |
| Immunoglobulin Levels (mg/mL) | ||||
|---|---|---|---|---|
| IgG | ||||
| CON | YC | Oat | YC + Oat | |
| Day 0 | 13.2 ± 6.1 ab | 14.7 ± 2.1 a | 6.0 ± 3.1 b | 12.4 ± 5.2 ab |
| Day 1 | 2.9 ± 1.0 | 7.9 ± 2.5 | 3.1 ± 0.9 | 5.0 ± 0.5 |
| Day 4 | 1.9 ± 0.4 | 3.4 ± 1.4 | 2.9 ± 1.3 | 3.5 ± 0.5 |
| Day 7 | 2.4 ± 1.1 | 3.5 ± 0.7 | 2.8 ± 2.0 | 5.0 ± 0.9 |
| Day 14 | 3.2 ± 0.8 | 4.0 ± 0.4 | 3.7 ± 1.5 | 3.6 ± 1.1 |
| IgM | ||||
| CON | YC | Oat | YC + Oat | |
| Day 0 | 6.7 ± 4.0 | 5.5 ± 3.1 | 4.6 ± 0.4 | 9.2 ± 5.4 |
| Day 1 | 2.4 ± 0.9 | 2.8 ± 1.6 | 1.9 ± 0.6 | 2.9 ± 0.9 |
| Day 4 | 1.6 ± 0.6 | 1.4 ± 0.3 | 1.9 ± 0.5 | 2.0 ± 1.2 |
| Day 7 | 1.9 ± 0.5 | 2.2 ± 0.6 | 4.7 ± 1.1 | 2.9 ± 1.1 |
| Day 14 | 1.9 ± 1.0 | 1.5 ± 0.3 | 3.3 ± 1.5 | 1.9 ± 0.5 |
| IgA | ||||
| CON | YC | Oat | YC + Oat | |
| Day 0 | 3.2 ± 1.7 | 3.4 ± 1.9 | 4.6 ± 1.9 | 6.2 ± 2.8 |
| Day 1 | 4.3 ± 1.3 | 6.8 ± 2.9 | 3.8 ± 1.3 | 5.2 ± 0.9 |
| Day 4 | 1.7 ± 1.3 | 3.3 ± 1.8 | 1.7 ± 0.2 | 1.7 ± 0.7 |
| Day 7 | 2.4 ± 1.1 | 3.5 ± 0.7 | 2.8 ± 2.0 | 5.0 ± 0.9 |
| Day 14 | 3.2 ± 0.8 | 4.0 ± 0.7 | 3.7 ± 1.5 | 3.6 ± 1.1 |
| GO Biological Process | ||||
|---|---|---|---|---|
| Term | Count | % | Benjamini | FDR |
| Platelet degranulation | 26 | 12.7 | 3.95 × 10−23 | 4.16 × 10−23 |
| Negative regulation of endopeptidase activity | 18 | 8.8 | 4.18 × 10−11 | 8.81 × 10−11 |
| GO Cellular Components | ||||
| Term | Count | % | Benjamini | FDR |
| Extracellular exosome | 165 | 80.5 | 7.33 × 10−98 | 3.04 × 10−97 |
| Extracellular space | 106 | 51.7 | 5.99 × 10−64 | 4.98 × 10−63 |
| Blood microparticle | 42 | 20.5 | 2.67 × 10−44 | 3.33 × 10−43 |
| KEGG Pathways | ||||
| Term | Count | % | Benjamini | FDR |
| Complement and coagulation cascades | 11 | 5.4 | 4.08 × 10−5 | 2.70 × 10−4 |
| Gene | Protein Names | CON-Oat | CON-YC | Oat-CON | YC-CON |
|---|---|---|---|---|---|
| BChE | Cholinesterase | −5.9 | −5.4 | 5.9 | 5.4 |
| SERPIND1 | Serpin family D member 1 | −5.5 | −6 | 5.5 | 6 |
| CHID1 | Chitinase domain-containing protein 1 precursor | −5.4 | −5.4 | 5.4 | 5.4 |
| CES1 | Carboxylic ester hydrolase | −0.9 | −0.4 | 0.9 | 0.4 |
| LTF | Lactoferrin | −0.8 | −1.1 | 0.8 | 1.1 |
| AHSG | Alpha-2-HS-glycoprotein | −0.8 | −0.5 | 0.8 | 0.5 |
| RAB7A | RAB7A; member RAS oncogene family | −0.4 | −0.5 | 0.4 | 0.5 |
| PPIC | Peptidyl-prolyl cis-trans isomerase | −0.1 | 0.3 | 0.1 | −0.3 |
| Uncharacterized protein | 0.3 | −0.7 | −0.3 | 0.7 | |
| ST13 | Hsp70 Interacting Protein | 1.5 | 1.7 | −1.5 | −1.7 |
| CALM2 | Calmodulin | 1.6 | 1.6 | −1.6 | −1.6 |
| RPLP0 | 60S acidic ribosomal protein P0 | 1.6 | 3.3 | −1.6 | −3.3 |
| TUBA1A | Tubulin alpha chain | 2 | 2.5 | −2 | −2.5 |
| EIF5A | Eukaryotic translation initiation factor 5A | 2.1 | 2.5 | −2.1 | −2.5 |
| RPS7 | 40S ribosomal protein S7 | 2.6 | 3 | −2.6 | −3 |
| VCP | Transitional endoplasmic reticulum ATPase | 2.7 | 2.9 | −2.7 | −2.9 |
| EEF2 | Eukaryotic translation elongation factor 2 | 2.7 | 2.6 | −2.7 | −2.6 |
| HSP90B1 | Endoplasmin | 2.7 | 3.7 | −2.7 | −3.7 |
| PDIA3 | Protein disulfide-isomerase | 3.1 | 4.6 | −3.1 | −4.6 |
| MDH2 | Malate dehydrogenase; mitochondrial | 3.3 | 4.1 | −3.3 | −4.1 |
| CKB | Creatine kinase B-type | 3.3 | 3.8 | −3.3 | −3.8 |
| RPSA | 40S ribosomal protein SA | 3.3 | 3.7 | −3.3 | −3.7 |
| RPLP2 | 60S acidic ribosomal protein P2 | 3.4 | 3.7 | −3.4 | −3.7 |
| RACK1 | Receptor of activated protein C kinase 1 | 3.4 | 4.6 | −3.4 | −4.2 |
| ACTN4 | Actinin alpha 4 | 3.7 | 2.1 | −3.7 | −2.1 |
| SRM | Spermidine Synthase | 3.7 | 3.8 | −3.7 | −3.8 |
| HSP90AA1 | Heat shock protein HSP 90-alpha | 3.8 | 3 | −3.8 | −3 |
| RNASE4 | Ribonuclease 4 | 4.3 | 0.5 | −4.3 | −0.5 |
| ATP5B | ATP synthase subunit beta | 4.5 | 6.3 | −4.5 | −6.3 |
| SERPINB1 | Leukocyte elastase inhibitor | 6.7 | −0.2 | −6.7 | 0.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Donovan, B.; Suarez-Trujillo, A.; Casey, T.; Aryal, U.K.; Conklin, D.; Williams, L.L.; Minor, R.C. Inclusion of Oat and Yeast Culture in Sow Gestational and Lactational Diets Alters Immune and Antimicrobial Associated Proteins in Milk. Animals 2021, 11, 497. https://doi.org/10.3390/ani11020497
Donovan B, Suarez-Trujillo A, Casey T, Aryal UK, Conklin D, Williams LL, Minor RC. Inclusion of Oat and Yeast Culture in Sow Gestational and Lactational Diets Alters Immune and Antimicrobial Associated Proteins in Milk. Animals. 2021; 11(2):497. https://doi.org/10.3390/ani11020497
Chicago/Turabian StyleDonovan, Barry, Aridany Suarez-Trujillo, Theresa Casey, Uma K. Aryal, Dawn Conklin, Leonard L. Williams, and Radiah C. Minor. 2021. "Inclusion of Oat and Yeast Culture in Sow Gestational and Lactational Diets Alters Immune and Antimicrobial Associated Proteins in Milk" Animals 11, no. 2: 497. https://doi.org/10.3390/ani11020497
APA StyleDonovan, B., Suarez-Trujillo, A., Casey, T., Aryal, U. K., Conklin, D., Williams, L. L., & Minor, R. C. (2021). Inclusion of Oat and Yeast Culture in Sow Gestational and Lactational Diets Alters Immune and Antimicrobial Associated Proteins in Milk. Animals, 11(2), 497. https://doi.org/10.3390/ani11020497