
Successful Inclusion of High Vegetable Protein Sources in Feed for Rainbow Trout without Decrement in Intestinal Health

 ,
,  ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Production System
2.2. Fish and Experimental Design
2.3. Diets and Feeding
2.4. Analysis of Intestinal Health Status
2.4.1. Intestinal Histology
2.4.2. Gene Expression
RNA Extraction
2.4.3. Quantitative Real-Time RT-PCR (qPCR)
Reference and Target Genes
RT-qPCR Conditions and Gene Expression Quantification
2.5. Statistical Analysis
3. Results
3.1. Growth, Survival and Food Intake
3.2. Intestinal Health Status
3.2.1. Intestinal Histology
3.2.2. Gene Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Martínez-Llorens, S.; Moñino, A.V.; Tomás Vidal, A.; Salvador, V.J.M.; Pla Torres, M.; Jover Cerdá, M. Soybean meal as a protein source in gilthead sea bream (Sparus aurata L.) diets: Effects on growth and nutrient utilization. Aquac. Res. 2007, 38, 82–90. [Google Scholar] [CrossRef]
- Gomes, E.F.; Rema, P.; Kaushik, S.J. Replacement of fish meal by plant proteins in the diet of rainbow trout (Oncorhynchus mykiss): Digestibility and growth performance. Aquaculture 1995, 130, 177–186. [Google Scholar] [CrossRef]
- Heikkinen, J.; Vielma, J.; Kemiläinen, O.; Tiirola, M.; Eskelinen, P.; Kiuru, T.; Navia-Paldanius, D.; von Wright, A. Effects of soybean meal based diet on growth performance, gut histopathology and intestinal microbiota of juvenile rainbow trout (Oncorhynchus mykiss). Aquaculture 2006, 261, 259–268. [Google Scholar] [CrossRef]
- Palmegiano, G.B.; Daprà, F.; Forneris, G.; Gai, F.; Gasco, L.; Guo, K.; Peiretti, P.G.; Sicuro, B.; Zoccarato, I. Rice protein concentrate meal as a potential ingredient in practical diets for rainbow trout (Oncorhynchus mykiss). Aquaculture 2006, 258, 357–367. [Google Scholar] [CrossRef]
- Drew, M.D.; Ogunkoya, A.E.; Janz, D.M.; Van Kessel, A.G. Dietary influence of replacing fish meal and oil with canola protein concentrate and vegetable oils on growth performance, fatty acid composition and organochlorine residues in rainbow trout (Oncorhynchus mykiss). Aquaculture 2007, 267, 260–268. [Google Scholar] [CrossRef]
- Bureau, D.; Harris, A.; Bevan, D.; Simmons, L.; Azevedo, P.; Cho, C. Feather meals and meat and bone meals from different origins as protein sources in rainbow trout (Oncorhynchus mykiss) diets. Aquaculture 2000, 181, 281–291. [Google Scholar] [CrossRef]
- Yoshitomi, B.; Aoki, M.; Oshima, S.; Hata, K. Evaluation of krill (Euphausia superba) meal as a partial replacement for fish meal in rainbow trout (Oncorhynchus mykiss) diets. Aquaculture 2006, 261, 440–446. [Google Scholar] [CrossRef]
- Aas, T.S.; Grisdale-Helland, B.; Terjesen, B.F.; Helland, S.J. Improved growth and nutrient utilisation in Atlantic salmon (Salmo salar) fed diets containing a bacterial protein meal. Aquaculture 2006, 259, 365–376. [Google Scholar] [CrossRef]
- Santigosa, E.; Sánchez, J.; Médale, F.; Kaushik, S.; Pérez-Sánchez, J.; Gallardo, M.A. Modifications of digestive enzymes in trout (Oncorhynchus mykiss) and sea bream (Sparus aurata) in response to dietary fish meal replacement by plant protein sources. Aquaculture 2008, 282, 68–74. [Google Scholar] [CrossRef]
- Lee, K.-J.; Powell, M.S.; Barrows, F.T.; Smiley, S.; Bechtel, P.; Hardy, R.W. Evaluation of supplemental fish bone meal made from Alaska seafood processing byproducts and dicalcium phosphate in plant protein based diets for rainbow trout (Oncorhynchus mykiss). Aquaculture 2010, 302, 248–255. [Google Scholar] [CrossRef]
- Storebakken, T.; Refstie, S.; Ruyter, B. Soy products as fat and protein sources in fish feeds for intensive aquaculture. In Soy In Animal Nutrition; Drackley, J.K., Ed.; Federation of Animal Science Societies: Savoy, IL, USA, 2000; pp. 127–170. [Google Scholar]
- Buttle, L.G.; Burrells, A.C.; Good, J.E.; Williams, P.D.; Southgate, P.J.; Burrells, C. The binding of soybean agglutinin (SBA) to the intestinal epithelium of Atlantic salmon, Salmo salar and Rainbow trout, Oncorhynchus mykiss, fed high levels of soybean meal. Vet. Immunol. Immunopathol. 2001, 80, 237–244. [Google Scholar] [CrossRef]
- Rumsey, G.L.; Siwicki, A.K.; Anderson, D.P.; Bowser, P.R. Effect of soybean protein on serological response, non-specific defense mechanisms, growth, and protein utilization in rainbow trout. Vet. Immunol. Immunopathol. 1994, 41, 323–339. [Google Scholar] [CrossRef]
- Lusas, E.W.; Riaz, M.N. Soy protein products: Processing and use. J. Nutr. 1995, 125, 573S–580S. [Google Scholar] [CrossRef]
- Kaushik, S.J.; Cravedi, J.P.; Lalles, J.P.; Sumpter, J.; Fauconneau, B.; Laroche, M. Partial or total replacement of fish-meal by soybean protein on growth, protein-utilization, potential estrogenic or antigenic effects, cholesterolemia and flesh quality in rainbow-trout, Oncorhynchus mykiss. Aquaculture 1995, 133, 257–274. [Google Scholar] [CrossRef]
- Gatlin, D.M., III; Barrows, F.T.; Brown, P.; Dabrowski, K.; Gaylord, T.G.; Hardy, R.W.; Herman, E.; Hu, G.; Krogdahl, Å.; Nelson, R.; et al. Expanding the utilization of sustainable plant products in aquafeeds: A review. Aquac. Res. 2007, 38, 551–579. [Google Scholar] [CrossRef]
- Oliva-Teles, A.; Enes, P.; Peres, H. 8—Replacing fishmeal and fish oil in industrial aquafeeds for carnivorous fish. In Feed and Feeding Practices in Aquaculture; Davis, D.A., Ed.; Woodhead Publishing: Sawston, UK, 2015; pp. 203–233. ISBN 978-0-08-100506-4. [Google Scholar]
- Burrells, C.; Williams, P.D.; Southgate, P.J.; Crampton, V.O. Immunological, physiological and pathological responses of rainbow trout (Oncorhynchus mykiss) to increasing dietary concentrations of soybean proteins. Vet. Immunol. Immunopathol. 1999, 72, 277–288. [Google Scholar] [CrossRef]
- Estruch, G.; Martínez-Llorens, S.; Tomás-Vidal, A.; Monge-Ortiz, R.; Jover-Cerdá, M.; Brown, P.B.; Peñaranda, D.S. Impact of high dietary plant protein with or without marine ingredients in gut mucosa proteome of gilthead seabream (Sparus aurata, L.). J. Proteom. 2020, 216, 103672. [Google Scholar] [CrossRef]
- Sitjà-Bobadilla, A.; Peña-Llopis, S.; Gómez-Requeni, P.; Médale, F.; Kaushik, S.; Pérez-Sánchez, J. Effect of fish meal replacement by plant protein sources on non-specific defence mechanisms and oxidative stress in gilthead sea bream (Sparus aurata). Aquaculture 2005, 249, 387–400. [Google Scholar] [CrossRef] [Green Version]
- Batista, S.; Ozório, R.O.A.; Kollias, S.; Dhanasiri, A.K.; Lokesh, J.; Kiron, V.; Valente, L.M.P.; Fernandes, J.M.O. Changes in intestinal microbiota, immune- and stress-related transcript levels in Senegalese sole (Solea senegalensis) fed plant ingredient diets intercropped with probiotics or immunostimulants. Aquaculture 2016, 458, 149–157. [Google Scholar] [CrossRef]
- Ye, G.; Dong, X.; Yang, Q.; Chi, S.; Liu, H.; Zhang, H.; Tan, B.; Zhang, S. Low-gossypol cottonseed protein concentrate used as a replacement of fish meal for juvenile hybrid grouper (Epinephelus fuscoguttatus ♀ × Epinephelus lanceolatus ♂): Effects on growth performance, immune responses and intestinal microbiota. Aquaculture 2020, 524, 735309. [Google Scholar] [CrossRef]
- Hernández, M.D.; Martínez, F.J.; Jover, M.; García García, B. Effects of partial replacement of fish meal by soybean meal in sharpsnout seabream (Diplodus puntazzo) diet. Aquaculture 2007, 263, 159–167. [Google Scholar] [CrossRef]
- Baeza-Ariño, R.; Martínez-Llorens, S.; Nogales-Mérida, S.; Jover-Cerda, M.; Tomás-Vidal, A. Study of liver and gut alterations in sea bream, Sparus aurata L., fed a mixture of vegetable protein concentrates. Aquac. Res. 2016, 47, 460–471. [Google Scholar] [CrossRef]
- Estruch, G.; Collado, M.C.; Peñaranda, D.S.; Tomás Vidal, A.; Jover Cerdá, M.; Pérez Martínez, G.; Martinez-Llorens, S. Impact of fishmeal replacement in DIETS for gilthead sea bream (Sparus aurata) on the gastrointestinal microbiota determined by pyrosequencing the 16S rRNA gene. PLoS ONE 2015, 10, e0136389. [Google Scholar] [CrossRef]
- Santigosa, E.; García-Meilán, I.; Valentin, J.M.; Pérez-Sánchez, J.; Médale, F.; Kaushik, S.; Gallardo, M.A. Modifications of intestinal nutrient absorption in response to dietary fish meal replacement by plant protein sources in sea bream (Sparus aurata) and rainbow trout (Onchorynchus mykiss). Aquaculture 2011, 317, 146–154. [Google Scholar] [CrossRef] [Green Version]
- Dimitroglou, A.; Merrifield, D.L.; Spring, P.; Sweetman, J.; Moate, R.; Davies, S.J. Effects of mannan oligosaccharide (MOS) supplementation on growth performance, feed utilisation, intestinal histology and gut microbiota of gilthead sea bream (Sparus aurata). Aquaculture 2010, 300, 182–188. [Google Scholar] [CrossRef]
- Davies, S.J.; Morris, P.C. Influence of multiple amino acid supplementation on the performance of rainbow trout, Oncorhynchus mykiss (Walbaum), fed soya based diets. Aquac. Res. 1997, 28, 65–74. [Google Scholar] [CrossRef]
- Yue, Y.; Zou, Z.; Zhu, J.; Li, D.; Xiao, W.; Han, J.; Yang, H. Effects of dietary arginine on growth performance, feed utilization, haematological parameters and non-specific immune responses of juvenile Nile tilapia (Oreochromis niloticus L.). Aquac. Res. 2015, 46, 1801–1809. [Google Scholar] [CrossRef]
- Lin, H.; Tan, X.; Zhou, C.; Niu, J.; Xia, D.; Huang, Z.; Wang, J.; Wang, Y. Effect of dietary arginine levels on the growth performance, feed utilization, non-specific immune response and disease resistance of juvenile golden pompano Trachinotus ovatus. Aquaculture 2015, 437, 382–389. [Google Scholar] [CrossRef]
- Cheng, Z.; Buentello, A.; Gatlin, D.M. Effects of dietary arginine and glutamine on growth performance, immune responses and intestinal structure of red drum, Sciaenops ocellatus. Aquaculture 2011, 319, 247–252. [Google Scholar] [CrossRef]
- Coutinho, F.; Simões, R.; Monge-Ortiz, R.; Furuya, W.M.; Pousão-Ferreira, P.; Kaushik, S.; Oliva-Teles, A.; Peres, H. Effects of dietary methionine and taurine supplementation to low-fish meal diets on growth performance and oxidative status of European sea bass (Dicentrarchus labrax) juveniles. Aquaculture 2017, 479, 447–454. [Google Scholar] [CrossRef]
- Feng, L.; Xiao, W.-W.; Liu, Y.; Jiang, J.; Hu, K.; Jiang, W.-D.; Li, S.-H.; Zhou, X.-Q. Methionine hydroxy analogue prevents oxidative damage and improves antioxidant status of intestine and hepatopancreas for juvenile Jian carp (Cyprinus carpio var. Jian). Aquac. Nutr. 2011, 17, 595–604. [Google Scholar] [CrossRef]
- Yang, Y.-H.; Wang, Y.-Y.; Lu, Y.; Li, Q.-Z. Effect of replacing fish meal with soybean meal on growth, feed utilization and nitrogen and phosphorus excretion on rainbow trout (Oncorhynchus mykiss). Aquac. Int. 2011, 19, 405–419. [Google Scholar] [CrossRef]
- Lee, S.; Masagounder, K.; Hardy, R.W.; Small, B.C. Effects of lowering dietary fishmeal and crude protein levels on growth performance, body composition, muscle metabolic gene expression, and chronic stress response of rainbow trout (Oncorhynchus mykiss). Aquaculture 2019, 513, 734435. [Google Scholar] [CrossRef]
- Burr, G.S.; Wolters, W.R.; Barrows, F.T.; Hardy, R.W. Replacing fishmeal with blends of alternative proteins on growth performance of rainbow trout (Oncorhynchus mykiss), and early or late stage juvenile Atlantic salmon (Salmo salar). Aquaculture 2012, 334–337, 110–116. [Google Scholar] [CrossRef]
- Watanabe, T.; Pongmaneerat, J.; Sato, S.; Takeuchi, T. Replacement of fish meal by alternative protein sources in rainbow trout diets. Nippon Suisan Gakkaishi 1993, 59, 1573–1579. [Google Scholar] [CrossRef]
- Aksnes, A.; Hope, B.; Jönsson, E.; Björnsson, B.T.; Albrektsen, S. Size-fractionated fish hydrolysate as feed ingredient for rainbow trout (Oncorhynchus mykiss) fed high plant protein diets. I: Growth, growth regulation and feed utilization. Aquaculture 2006, 261, 305–317. [Google Scholar] [CrossRef]
- Ostaszewska, T.; Dabrowski, K.; Palacios, M.E.; Olejniczak, M.; Wieczorek, M. Growth and morphological changes in the digestive tract of rainbow trout (Oncorhynchus mykiss) and pacu (Piaractus mesopotamicus) due to casein replacement with soybean proteins. Aquaculture 2005, 245, 273–286. [Google Scholar] [CrossRef]
- Bureau, D.P.; Harris, A.M.; Young Cho, C. The effects of purified alcohol extracts from soy products on feed intake and growth of chinook salmon (Oncorhynchus tshawytscha) and rainbow trout (Oncorhynchus mykiss). Aquaculture 1998, 161, 27–43. [Google Scholar] [CrossRef]
- Romarheim, O.H.; Skrede, A.; Penn, M.; Mydland, L.T.; Krogdahl, Å.; Storebakken, T. Lipid digestibility, bile drainage and development of morphological intestinal changes in rainbow trout (Oncorhynchus mykiss) fed diets containing defatted soybean meal. Aquaculture 2008, 274, 329–338. [Google Scholar] [CrossRef]
- Jalili, R.; Tukmechi, A.; Agh, N.; Noori, F.; Ghasemi, A. Replacement of dietary fish meal with plant sources in rainbow trout (Oncorhynchus mykiss); effect on growth performance, immune responses, blood indices and disease resistance. Iran. J. Fish. Sci. 2013, 12, 577–591. [Google Scholar]
- Penn, M.H.; Bendiksen, E.Å.; Campbell, P.; Krogdahl, Å. High level of dietary pea protein concentrate induces enteropathy in Atlantic salmon (Salmo salar L.). Aquaculture 2011, 310, 267–273. [Google Scholar] [CrossRef]
- Venou, B.; Alexis, M.N.; Fountoulaki, E.; Haralabous, J. Effects of extrusion and inclusion level of soybean meal on diet digestibility, performance and nutrient utilization of gilthead sea bream (Sparus aurata). Aquaculture 2006, 261, 343–356. [Google Scholar] [CrossRef]
- Hardy, R.W. Rainbow trout, Oncorhynchus mykiss. In Nutrient Requirements and Feeding of Finfish for Aquaculture; Webster, C.D., Lim, C., Eds.; CABI Publishing: New York, NY, USA, 2002; pp. 184–202. [Google Scholar]
- National Research Council (NRC). Nutrient Requirements of Fish and Shrimp; The National Academic Press: Washington, DC, USA, 2011. [Google Scholar]
- Brouwer, E. Report of Sub-Committee on Constants and Factors. Proceedings of the 3rd Symposium on Energy Metabolism of Farm Animals; EAAP Publ. No. 11; Blaxter, K.L., Ed.; Academic Press: London, UK, 1965; pp. 441–443. [Google Scholar]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper—Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Angosto, D.; López-Castejón, G.; López-Muñoz, A.; Sepulcre, M.P.; Arizcun, M.; Meseguer, J.; Mulero, V. Evolution of inflammasome functions in vertebrates: Inflammasome and caspase-1 trigger fish macrophage cell death but are dispensable for the processing of IL-1β. Innate Immun. 2012, 18, 815–824. [Google Scholar] [CrossRef]
- Peddie, S.; Zou, J.; Cunningham, C.; Secombes, C.J. Rainbow trout (Oncorhynchus mykiss) recombinant IL-1β and derived peptides induce migration of head-kidney leucocytes in vitro. Fish Shellfish Immunol. 2001, 11, 697–709. [Google Scholar] [CrossRef]
- Elsässer-Beile, U.; Von Kleist, S. Cytokines as therapeutic and diagnostic agents. Tumor Biol. 1993, 14, 69–94. [Google Scholar] [CrossRef]
- Sporn, M.B. TGF-β: 20 years and counting. Microbes Infect. 1999, 1, 1251–1253. [Google Scholar] [CrossRef]
- Kim, D.-H.; Austin, B. Cytokine expression in leucocytes and gut cells of rainbow trout, Oncorhynchus mykiss Walbaum, induced by probiotics. Vet. Immunol. Immunopathol. 2006, 114, 297–304. [Google Scholar] [CrossRef]
- Bullen, J.J.; Rogers, H.J.; Griffiths, E. Role of iron in bacterial infection. Curr. Top. Microbiol. Immunol. 1978, 80, 1–35. [Google Scholar] [CrossRef]
- Zhang, Y.-A.; Salinas, I.; Li, J.; Parra, D.; Bjork, S.; Xu, Z.; LaPatra, S.E.; Bartholomew, J.; Sunyer, J.O. IgT, a primitive immunoglobulin class specialized in mucosal immunity. Nat. Immunol. 2010, 11, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Secombes, C. The function of fish cytokines. Biology 2016, 5, 23. [Google Scholar] [CrossRef] [PubMed]
- Evenhuis, J.P.; Cleveland, B.M. Modulation of rainbow trout (Oncorhynchus mykiss) intestinal immune gene expression following bacterial challenge. Vet. Immunol. Immunopathol. 2012, 146, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Kania, P.W.; Chettri, J.K.; Buchmann, K. Characterization of serum amyloid A (SAA) in rainbow trout using a new monoclonal antibody. Fish Shellfish Immunol. 2014, 40, 648–658. [Google Scholar] [CrossRef]
- Talbot, A.T.; Pottinger, T.G.; Smith, T.J.; Cairns, M.T. Acute phase gene expression in rainbow trout (Oncorhynchus mykiss) after exposure to a confinement stressor: A comparison of pooled and individual data. Fish Shellfish Immunol. 2009, 27, 309–317. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Sánchez, T.; Balcázar, J.L.; Merrifield, D.L.; Carnevali, O.; Gioacchini, G.; de Blas, I.; Ruiz-Zarzuela, I. Expression of immune-related genes in rainbow trout (Oncorhynchus mykiss) induced by probiotic bacteria during Lactococcus garvieae infection. Fish Shellfish Immunol. 2011, 31, 196–201. [Google Scholar] [CrossRef]
- Weltzien, F.-A.; Pasqualini, C.; Vernier, P.; Dufour, S. A quantitative real-time RT-PCR assay for European eel tyrosine hydroxylase. Gen. Comp. Endocrinol. 2005, 142, 134–142. [Google Scholar] [CrossRef]
- Snedecor, G.; Cochran, W. Statistical Methods, 6th ed.; Iowa State University Press: Ames, IA, USA, 1971. [Google Scholar]
- Sagner, G.; Goldstein, C. Principles, workflows and advantages of the new LightCycler Relative Quantification Software. Biochemica 2001, 3, 15–17. [Google Scholar]
- Goff, J.B.; Gatlin, D.M. Evaluation of different sulfur amino acid compounds in the diet of red drum, Sciaenops ocellatus, and sparing value of cystine for methionine. Aquaculture 2004, 241, 465–477. [Google Scholar] [CrossRef]
- Grimble, R.F. The effects of sulfur amino acid intake on immune function in humans. J. Nutr. 2006, 136, 1660S–1665S. [Google Scholar] [CrossRef] [Green Version]
- Choi, D.G.; He, M.; Fang, H.; Wang, X.L.; Li, X.Q.; Leng, X.J. Replacement of fish meal with two fermented soybean meals in diets for rainbow trout (Oncorhynchus mykiss). Aquac. Nutr. 2020, 26, 37–46. [Google Scholar] [CrossRef]
- Sanz, A.; Morales, A.E.; de la Higuera, M.; Gardenete, G. Sunflower meal compared with soybean meals as partial substitutes for fish meal in rainbow trout (Oncorhynchus mykiss) diets: Protein and energy utilization. Aquaculture 1994, 128, 287–300. [Google Scholar] [CrossRef]
- Haghbayan, S.; Shamsaie Mehrgan, M. The effect of replacing fish meal in the diet with enzyme-treated soybean meal (HP310) on growth and body composition of rainbow trout fry. Molecules 2015, 20, 21058–21066. [Google Scholar] [CrossRef] [Green Version]
- Alami-Durante, H.; Médale, F.; Cluzeaud, M.; Kaushik, S.J. Skeletal muscle growth dynamics and expression of related genes in white and red muscles of rainbow trout fed diets with graded levels of a mixture of plant protein sources as substitutes for fishmeal. Aquaculture 2010, 303, 50–58. [Google Scholar] [CrossRef]
- Hua, K.; Cobcroft, J.M.; Cole, A.; Condon, K.; Jerry, D.R.; Mangott, A.; Praeger, C.; Vucko, M.J.; Zeng, C.; Zenger, K.; et al. The future of aquatic protein: Implications for protein sources in aquaculture diets. One Earth 2019, 1, 316–329. [Google Scholar] [CrossRef] [Green Version]
- Krogdahl, Å.; Penn, M.; Thorsen, J.; Refstie, S.; Bakke, A.M. Important antinutrients in plant feedstuffs for aquaculture: An update on recent findings regarding responses in salmonids. Aquac. Res. 2010, 41, 333–344. [Google Scholar] [CrossRef]
- Davis, D.A.; Hardy, R.W. Chapter 14—Feeding and Fish Husbandry, 4th ed.; Hardy, R.W., Kaushik, S.J.B.T.-F.N., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 857–882. ISBN 978-0-12-819587-1. [Google Scholar]
- Cheng, Z.J.; Hardy, R.W.; Usry, J.L. Effects of lysine supplementation in plant protein-based diets on the performance of rainbow trout (Oncorhynchus mykiss) and apparent digestibility coefficients of nutrients. Aquaculture 2003, 215, 255–265. [Google Scholar] [CrossRef]
- Takeuchi, T.; Park, G.-S.; Seikai, T.; Yokoyama, M. Taurine content in Japanese flounder Paralichthys olivaceus T. & S. and red sea bream Pagrus major T. & S. during the period of seed production. Aquac. Res. 2001, 32, 244–248. [Google Scholar] [CrossRef]
- Yokoyama, M.; Kaneniwa, M.; Sakaguchi, M. Metabolites of L-[35S]Cysteine injected into the peritoneal cavity of rainbow trout. Fish. Sci. 1997, 63. [Google Scholar] [CrossRef] [Green Version]
- Gibson Gaylord, T.; Barrows, F.T.; Teague, A.M.; Johansen, K.A.; Overturf, K.E.; Shepherd, B. Supplementation of taurine and methionine to all-plant protein diets for rainbow trout (Oncorhynchus mykiss). Aquaculture 2007, 269, 514–524. [Google Scholar] [CrossRef]
- Gaylord, T.G.; Teague, A.M.; Barrows, F.T. Taurine supplementation of all-plant protein diets for rainbow trout (Oncorhynchus mykiss). J. World Aquac. Soc. 2006, 37, 509–517. [Google Scholar] [CrossRef]
- Francis, G.; Makkar, H.P.; Becker, K. Antinutritional factors present in plant-derived alternate fish feed ingredients and their effects in fish. Aquaculture 2001, 199, 197–227. [Google Scholar] [CrossRef]
- Øverland, M.; Sørensen, M.; Storebakken, T.; Penn, M.; Krogdahl, Å.; Skrede, A. Pea protein concentrate substituting fish meal or soybean meal in diets for Atlantic salmon (Salmo salar)—Effect on growth performance, nutrient digestibility, carcass composition, gut health, and physical feed quality. Aquaculture 2009, 288, 305–311. [Google Scholar] [CrossRef]
- Baeverfjord, G.; Krogdahl, A. Development and regression of soybean meal induced enteritis in Atlantic salmon, Salmo salar L., distal intestine: A comparison with the intestines of fasted fish. J. Fish Dis. 1996, 19, 375–387. [Google Scholar] [CrossRef]
- Van den Ingh, T.S.G.A.M.; Krogdahl, Å.; Olli, J.J.; Hendriks, H.G.C.J.M.; Koninkx, J.G.J.F. Effects of soybean-containing diets on the proximal and distal intestine in Atlantic salmon (Salmo salar): A morphological study. Aquaculture 1991, 94, 297–305. [Google Scholar] [CrossRef]
- Tomás-Almenar, C.; Toledo-Solís, F.J.; Larrán, A.M.; de Mercado, E.; Alarcón, F.J.; Rico, D.; Martín-Diana, A.B.; Fernández, I. Effects and safe inclusion of narbonne vetch (Vicia narbonensis) in rainbow trout (Oncorhynchus mykiss) Diets: Towards a more sustainable aquaculture. Animals 2020, 10, 2175. [Google Scholar] [CrossRef]
- Urán, P.A.; Schrama, J.W.; Rombout, J.H.W.M.; Taverne-Thiele, J.J.; Obach, A.; Koppe, W.; Verreth, J.A.J. Time-related changes of the intestinal morphology of Atlantic salmon, Salmo salar L., at two different soybean meal inclusion levels. J. Fish Dis. 2009, 32, 733–744. [Google Scholar] [CrossRef]
- Martínez-Llorens, S.; Baeza-Ariño, R.; Nogales-Mérida, S.; Jover-Cerdá, M.; Tomás-Vidal, A. Carob seed germ meal as a partial substitute in gilthead sea bream (Sparus aurata) diets: Amino acid retention, digestibility, gut and liver histology. Aquaculture 2012, 338–341, 124–133. [Google Scholar] [CrossRef]
- Olsen, R.E.; Hansen, A.-C.; Rosenlund, G.; Hemre, G.-I.; Mayhew, T.M.; Knudsen, D.L.; Tufan Eroldoğan, O.; Myklebust, R.; Karlsen, Ø. Total replacement of fish meal with plant proteins in diets for Atlantic cod (Gadus morhua L.) II—Health aspects. Aquaculture 2007, 272, 612–624. [Google Scholar] [CrossRef]
- Reyes-Becerril, M.; Guardiola, F.; Rojas, M.; Ascencio-Valle, F.; Esteban, M.Á. Dietary administration of microalgae Navicula sp. affects immune status and gene expression of gilthead seabream (Sparus aurata). Fish Shellfish Immunol. 2013, 35, 883–889. [Google Scholar] [CrossRef]
- Secombes, C.J.; Wang, T.; Hong, S.; Peddie, S.; Crampe, M.; Laing, K.J.; Cunningham, C.; Zou, J. Cytokines and innate immunity of fish. Dev. Comp. Immunol. 2001, 25, 713–723. [Google Scholar] [CrossRef]
- Bird, S.; Zou, J.; Wang, T.; Munday, B.; Cunningham, C.; Secombes, C.J. Evolution of interleukin-1β. Cytokine Growth Factor Rev. 2002, 13, 483–502. [Google Scholar] [CrossRef]
- Laing, K.J.; Secombes, C.J. Chemokines. Dev. Comp. Immunol. 2004, 28, 443–460. [Google Scholar] [CrossRef]
- Calduch-Giner, J.A.; Sitjà-Bobadilla, A.; Davey, G.C.; Cairns, M.T.; Kaushik, S.; Pérez-Sánchez, J. Dietary vegetable oils do not alter the intestine transcriptome of gilthead sea bream (Sparus aurata), but modulate the transcriptomic response to infection with Enteromyxum leei. BMC Genom. 2012, 13, 470. [Google Scholar] [CrossRef] [Green Version]
- Bernard, D.; Six, A.; Rigottier-Gois, L.; Messiaen, S.; Chilmonczyk, S.; Quillet, E.; Boudinot, P.; Benmansour, A. Phenotypic and functional similarity of gut intraepithelial and systemic T cells in a teleost fish. J. Immunol. 2006, 176, 3942–3949. [Google Scholar] [CrossRef]
- Rombout, J.H.W.M.; Picchietti, S.; Kiron, V. Teleost intestinal immunology. Fish Shellfish Immunol. 2011, 31, 616–626. [Google Scholar] [CrossRef] [Green Version]
- Letterio, J.J.; Roberts, A.B. Regulation of immune responses by TGF-β. Annu. Rev. Immunol. 1998, 16, 137–161. [Google Scholar] [CrossRef] [Green Version]
- Cerezuela, R.; Guardiola, F.A.; Meseguer, J.; Esteban, M.Á. Enrichment of gilthead seabream (Sparus aurata L.) diet with microalgae: Effects on the immune system. Fish Physiol. Biochem. 2012, 38, 1729–1739. [Google Scholar] [CrossRef]
- Vendrell, D.; Luis Balcázar, J.; de Blas, I.; Ruiz-Zarzuela, I.; Gironés, O.; Luis Múzquiz, J. Protection of rainbow trout (Oncorhynchus mykiss) from lactococcosis by probiotic bacteria. Comp. Immunol. Microbiol. Infect. Dis. 2008, 31, 337–345. [Google Scholar] [CrossRef]

{kind=link}
| Experimental Diets | |||
|---|---|---|---|
| FM0 | FM10 | FM20 | |
| Ingredients (g kg−1) | |||
| Fishmeal 1 | 0 | 100 | 200 |
| Wheat meal 2 | 116 | 158 | 201 |
| Wheat gluten 3 | 214 | 201 | 180 |
| Soybean meal 4 | 400 | 300 | 200 |
| Soybean oil | 88 | 89 | 90 |
| Fish oil | 100 | 91 | 82 |
| Calcium phosphate | 38 | 33 | 28 |
| Taurine 5 | 20 | 10 | 5 |
| Valine 5 | 2 | 0 | 0 |
| L- Methionine 5 | 4 | 2 | 0 |
| L-Lysine Clh 5 | 8 | 6 | 4 |
| Vitamin-mineral mix 6 | 10 | 10 | 10 |
| Proximate composition (g kg−1 on dry matter) | |||
| Dry matter (DM) | 913 | 912 | 912 |
| Crude protein (CP) | 446 | 440 | 434 |
| Crude lipid (CL) | 191 | 190 | 192 |
| Ash | 42 | 68 | 75 |
| Calculated values | |||
| Carbohydrates (CHO) 7 | 322 | 302 | 299 |
| Calculated GE (MJ kg−1) 8 | 22.5 | 22.6 | 23.3 |
| Category | Gen/Protein Description | Abbrev. | Primer Sequence | GenBank | BP Size | Reference | |
|---|---|---|---|---|---|---|---|
| Forward | Reverse | ||||||
| Housekeeping genes | Beta-actin | β-actin | GCCGGCCGCGACCTCACAGACTAC | CGGCCGTGGTGGTGAAGCTGTAAC | EZ908974 | 73 | Evenhuis and Cleveland, [58] |
| Elongation factor 1-alpha | ELF-1α | ACCCTCCTCTTGGTCGTTTC | TGATGACACCAACAGCAACA | AF498320 | 63 | Kania et al. [59] | |
| Transferrin | 5’CCACCTCCAGGGCCATTAAATG3’ | 5’ATCCACCGCTATGGCATCTGCC3’ | D89083 | Talbot et al. [60] | |||
| Immune | Immunoglobulin T | IgT | AACATCACCTGGCACATCAA | TTCAGGTTGCCCTTTGATTC | AY870265 | 80 | Evenhuis and Cleveland, [58] |
| Interferon gamma | IFN-γ | CTGTTCAACGGAAACCCTGT | AACACCCTCCGATCACTGTC | NM 001160503 | 62 | Evenhuis and Cleveland, [58] | |
| Interleukin-1 beta | IL-1β | ACATTGCCAACCTCATCATCG | TTGAGCAGGTCCTTGTCCTTG | AJ223954 | 91 | Pérez-Sánchez et al. [61] | |
| Inflammatory | Interleukin 8 | IL-8 | CTCGCAACTGGACTGACAAA | TGGCTGACATTCTGATGCTC | AJ279069 | 148 | Evenhuis and Cleveland, [58] |
| Transforming growth factor beta | TGF-β | TCCGCTTCAAAATATCAGGG | TGATGGCATTTTCATGGCTA | X99303 | 71 | Evenhuis and Cleveland, [58] | |
| Experimental Diets | ||||
|---|---|---|---|---|
| FM0 | FM10 | FM20 | p-Value | |
| Initial weight (g) | 13.60 ± 0.23 | 13.43 ± 0.23 | 13.13 ± 0.23 | 0.3911 |
| Final weight (g) | 49.07 b ± 1.69 | 72.94 a ± 1.69 | 72.24 a ± 1.69 | 0.0001 |
| Survival (%) | 51.11 b ± 5.62 | 81.67 a ± 5.62 | 93.33 a ± 5.62 | 0.0046 |
| SGR 1 (% day−1). | 1.73 b ± 0.04 | 2.24 a ± 0.04 | 2.21 a ± 0.04 | 0.0043 |
| FI (g 100 g fish−1 day−1) 2 | 2.10 a ± 0.06 | 1.85 b ± 0.06 | 1.79 b ± 0.06 | 0.0188 |
| FCR 3 | 1.52 b ± 0.06 | 1.09 a ± 0.06 | 1.01 a ± 0.06 | 0.0018 |
| PER 4 | 1.62 b ± 0.09 | 2.31 a ± 0.09 | 2.49 a ± 0.09 | 0.0025 |
| Experimental Diets | ||||
|---|---|---|---|---|
| FM0 | FM10 | FM20 | p-Value | |
| Proximal Intestine | ||||
| VL (µm) | 782.8 ± 77.6 | 831.6 ± 56.0 | 715.2 ± 47.5 | 0.2850 |
| VT (µm) | 209.9 b ± 14.1 | 164.6 a ± 10.2 | 149.8 a ± 8.6 | 0.0026 |
| LP (µm) | 53.7 b ± 4.5 | 35.4 a ± 3.3 | 33.7 a ± 2.8 | 0.0012 |
| GC | 2.7 a ± 1.5 | 8.9 b ± 1.0 | 15.2 c ± 0.9 | 0.0000 |
| SL (µm) | 67.8 ± 5.3 | 55.3 ± 4.7 | 62.7 ± 4.6 | 0.2184 |
| ML (µm) | 152.7 ± 13.0 | 117.3 ± 11.6 | 140.6 ± 11.4 | 0.1214 |
| SML (µm) | 60.8 ± 5.0 | 59.3 ±4.5 | 57.5 ± 4.4 | 0.8834 |
| Distal Intestine | ||||
| VL (µm) | 547.5 ± 134.3 | 754.2 ± 54.8 | 756.8 ± 60.0 | 0.3396 |
| VT (µm) | 135.91 ± 26.9 | 160.88 ± 11.1 | 168.4 ± 11.8 | 0.5404 |
| LP (µm) | 26.9 ± 3.7 | 31.0 ± 1.5 | 33.2 ± 1.6 | 0.2633 |
| GC | 2.1 a ± 2.8 | 9.4 b ± 1.1 | 14.0 b ± 1.2 | 0.0006 |
| SL (µm) | 70.9 ± 12.6 | 80.0 ± 7.0 | 67.3 ± 9.6 | 0.5428 |
| ML (µm) | 93.3 ± 15.8 | 110.1 ± 8.7 | 110.4 ± 11.9 | 0.6266 |
| SML (µm) | 66.2 ± 7.1 | 50.1 ± 3.9 | 56.3 ± 5.3 | 0.1437 |
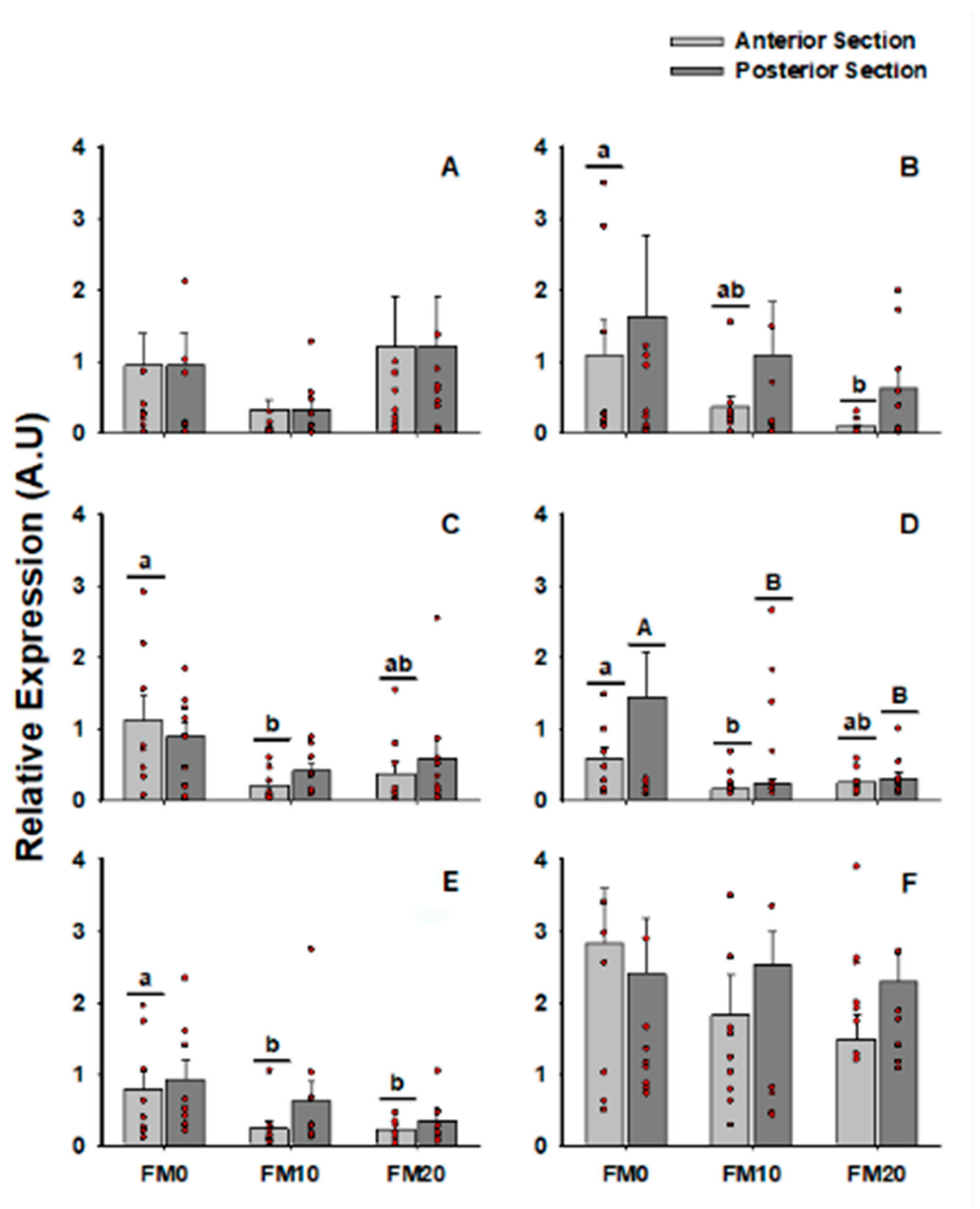
| IgT | Transferrin | IFN-γ | IL-8 | IL-1β | TGF-β | |
|---|---|---|---|---|---|---|
| Function | Immune system | Immune system | Immune system | Inflammatory | Inflammatory | Inflammatory |
| Section | 0.1966 | 0.2457 | 0.7111 | 0.1358 | 0.201 | 0.4442 |
| Diet | 0.1905 | 0.2857 | 0.0042 * | 0.0088 * | 0.018 * | 0.4428 |
| Interaction (section × diet) | ||||||
| FM0-Ant vs. FM10-Ant vs. FM20-Ant | 0.2137 | 0.0608 | 0.0177 * | 0.014 * | 0.0271 * | 0.2446 |
| FM0-Pos vs. FM10-Pos vs. FM20-Pos | 0.4101 | 0.6864 | 0.2198 | 0.0512 | 0.2386 | 0.9588 |
| FM0-Ant vs. FM0-Pos | 0.8031 | 0.8249 | 0.5844 | 0.2244 | 0.7244 | 0.6994 |
| FM10-Ant vs. FM10-Pos | 0.1086 | 0.3628 | 0.1074 | 0.1703 | 0.2292 | 0.3474 |
| FM20-Ant vs. FM20-Pos | 0.2350 | 0.0620 | 0.5153 | 0.6035 | 0.3455 | 0.1546 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vélez-Calabria, G.; Peñaranda, D.S.; Jover-Cerdá, M.; Llorens, S.M.; Tomás-Vidal, A. Successful Inclusion of High Vegetable Protein Sources in Feed for Rainbow Trout without Decrement in Intestinal Health. Animals 2021, 11, 3577. https://doi.org/10.3390/ani11123577
Vélez-Calabria G, Peñaranda DS, Jover-Cerdá M, Llorens SM, Tomás-Vidal A. Successful Inclusion of High Vegetable Protein Sources in Feed for Rainbow Trout without Decrement in Intestinal Health. Animals. 2021; 11(12):3577. https://doi.org/10.3390/ani11123577
Chicago/Turabian StyleVélez-Calabria, Glenda, David Sánchez Peñaranda, Miguel Jover-Cerdá, Silvia Martínez Llorens, and Ana Tomás-Vidal. 2021. "Successful Inclusion of High Vegetable Protein Sources in Feed for Rainbow Trout without Decrement in Intestinal Health" Animals 11, no. 12: 3577. https://doi.org/10.3390/ani11123577
APA StyleVélez-Calabria, G., Peñaranda, D. S., Jover-Cerdá, M., Llorens, S. M., & Tomás-Vidal, A. (2021). Successful Inclusion of High Vegetable Protein Sources in Feed for Rainbow Trout without Decrement in Intestinal Health. Animals, 11(12), 3577. https://doi.org/10.3390/ani11123577