1. Introduction
Enzymes have been widely used in animal production for optimizing nutrient digestibility and growth performance in all animal species. Exogenous enzymes supplemented with monogastric diets can contribute to the removal of antinutritional factors, modulating the intestinal microbiota, increasing the digestibility of nutrients and their utilization leading to improvements in intestinal health and growth performance [
1,
2,
3].
Around 61–70% of the phosphorus (P) in cereal grains and oilseeds used in monogastric diet formulation are present in the form of phytic acid [
4]. However, monogastric animals do not produce endogenous phytase and, consequently, the P content in the phytic acid form is not available for utilization by poultry [
4,
5]. Furthermore, phytic acid can also bind to proteins and enzymes like trypsin and α-amylase, inhibiting their activity and lowering protein and carbohydrate digestibility [
6,
7,
8]. The undigested nutrients would be eliminated via excreta by the animals, raising an environmental concern [
9]. When phytase is included in feeds there is a reduction in supplemental levels of calcium (Ca) and P to compensate for the increased uptake of these minerals provided by phytase supplementation [
10,
11]. A high supplementation of Ca alter the physical and chemical properties of the digesta in the gastrointestinal tract and lead to alterations in pH and solubility, which can affect the relative abundance and diversity of the intestinal microbiota [
12,
13].
The dietary inclusion of phytase can release the P in the phytic acid form, implying the degradation of insoluble antinutritional inositol hexaphosphoric acid and subsequent generation of lower inositol phosphates into inositol. [
14,
15,
16]. In principle, phytase can hydrolyze phytate and convert it to lower myo-inositol phosphate esters, inositol, and inorganic P through a sequence of stepwise dephosphorylation reactions [
16,
17,
18]. Part of the benefit of supplementing phytase could be attributed to catalyzing hydrolysis reaction to release inorganic P from phytate and consequently the generation of inositol from the hydrolysis of phytate [
14,
16,
19].
The supplementation of phytase for poultry has been widely studied for different benefits, such as bone parameters, growth performance, nutrient digestibility, and intestinal health [
14,
20,
21,
22]. However, some studies reported that the supplementation of phytase can also play an important role in modulating the intestinal microbiota by reducing harmful and increasing beneficial bacteria, which in turn can exert benefits associated with nutrient digestibility, intestinal morphology, and bone parameters of broiler chickens [
23,
24,
25]. The role of dietary phytase on the intestinal microbiota can be associated with the buffering property and availability of Ca and P for microbial fermentation [
23,
24,
25]. According to Ptak et al. [
23], the Ca and P released by hydrolysis of phytate by phytase increased the proliferation of lactic acid bacteria in broiler fed low Ca and P diet. The higher concentration of short-chain fatty acids and lactic acid can improve the intestinal environment, consequently, enhancing intestinal health and nutrient digestibility [
23,
25]. In addition, by hydrolyzing the phytate other minerals and nutrients that were complexed with the phytate molecule can be released further improving bone parameters and intestinal morphology [
23,
24,
25].
Thus, it was hypothesized that the supplementation of phytase could positively modulate the mucosa-associated microbiota in the jejunum by increasing beneficial and reducing pathogen bacteria and consequently enhancing intestinal morphology nutrient digestibility and bone parameters in broiler chickens. Therefore, the objectives of this study were to determine the effects of increasing doses of phytase on the modulation of the mucosa-associated microbiota in the jejunum and the impacts on the nutrient digestibility, intestinal morphology, bone parameters, and growth performance of broiler chickens.
4. Discussion
Phytase supplementation for broiler diets has been associated with improvements in growth performance, nutrient digestibility, bone parameters, and its efficacy has been established with a large number of studies over time [
8,
22,
34,
35]. Recently, the study of the mucosa-associated microbiota in the jejunum has gained more attention due to the relevance for the metabolism of nutrients, stimulation of immune response, protection from pathogens, and stimulation of epithelium cell proliferation of the animals [
25,
36]. Modulation of the microbiota toward a more healthy one may reflect improvements in the health and productive performance of monogastric animals [
3,
23,
24,
37]. The microbiota composition can vary among species and sections of the organism, for example, the chicken’s intestinal microbiota is composed of more than 900 species [
24,
38].
In the present study, the mucosa-associated microbiota in the jejunum was mainly composed of four phyla with
Firmicutes,
Proteobacteria,
Cyanobacteria, and
Actinobacteria accounting for more than 85% of the total mucosa-associated microbiota in the jejunum. The intestinal microbiota can play an essential role in nutrient digestibility, hydrolysis of antinutritional factors and toxins, decrease in pathogens, stimulation, and modulation of the immune system and metabolism [
24,
39]. The solubility and stability of phytate-mineral complexes are both pH dependents [
12,
40,
41], and most of these complexes are soluble at a low pH lower than 4 and insoluble between a pH 4 and 7 [
12]. The formation of phytate-mineral complexes can impact phytase efficacy [
12,
42]. A low solubility may have an impact on phytate P digestibility and availability because is directly influenced by the hydrolysis of the phytate-mineral complexes [
12,
41] and because the pH of the broilers’ small intestinal is between 5.5 and 6.6, it can favor the formation of these complexes [
12,
13].
Mineral cations such as Zn
2+, Fe
2+, Mn
2+, Fe
3+, Ca
2+, and Mg
2+ have different properties regarding the formation and hydrolysis of phytate-mineral complexes [
40]. According to Maenz et al. [
40], zinc and iron have been reported with higher affinity and Ca and magnesium with lower affinity with the phytate molecule. Even though Ca ions have a low affinity with the phytate molecule, it may cause more impact on the hydrolysis of the phytate molecule because traditionally in poultry diets, Ca is a mineral added in a high concentration. Different authors attributed the negative effects of a high Ca supplementation on the hydrolysis of the phytate molecule due to a combination of factors [
12,
22]. It may be due to the precipitation of phytate by Ca through Ca-phytate-complex formation [
12,
40] or by an increase in the intestinal pH caused by the supplementation of Ca which will lead to a reduction of the mineral solubility and consequently availability [
12,
43]. A high supplementation of Ca can alter the physical and chemical properties of the digesta in the gastrointestinal tract [
12,
22]. Shafey et al. [
43], found that by increasing Ca the pH of the digesta in the crop and ileum was increased, although alterations on the digesta pH may reflect on different shifts in the microbiota population.
A lower pH in the small intestine may provide a bacteriostatic effect, which is positive for intestinal integrity and the characteristics of the microbiota population by reducing the occurrence of pathogenic bacteria such as
Enterobacteriaceae and
Heliboacteraceae, and increasing beneficial bacteria, such as lactic acid bacteria populations [
23,
44]. These findings are in agreement with the results of the present study, where phytase supplementation tended to reduce the relative abundance of
Enterobacteriaceae and
Heliboacteraceae and to increase the relative abundance of
Lactobacillus. Also, it tended to reduce the relative abundance of
Pelomonas,
Microbacterium,
Pseudomonas,
Methylobacterium,
Sphingomonas. Furthermore, phytase supplementation tended to increase the relative abundance of
Lactobacillus reuteri, which is a Gram-positive bacteria also known as lactic acid bacteria that can play a role as a probiotic for different species [
45,
46]. Different associated benefits were reported with a high abundance of this bacteria, such as the production of antimicrobial components (organic acids, ethanol, and reuterin) that can lead to inhibition of the colonization of pathogenic microbes and remodeling the commensal microbiota composition [
45,
47,
48]. Additionally, it can strengthen the intestinal barrier and may decrease the microbial translocation from the gut lumen to the tissues [
45,
49]. However, it is important to keep in mind that different diet compositions can affect and interact with different microbial communities throughout the different parts of the digestive tract of the birds, which can primarily account for a large variation among the replicates [
24].
A low microbial diversity can be associated with diets containing high levels of Ca and may lead to a decrease in the growth performance [
24] and the energy value of the diet [
50]. The bacterial growth can be affected by different Ca levels in a wide range of factors such as, by reducing acidic fermentation and precipitation of cytotoxic components in the intestinal lumen [
23,
51], by increasing gastric acid secretion [
23,
52], and by reducing pathogenic adhesion on the intestinal mucosal walls [
23,
53,
54]. In the present study where we found that phytase supplementation increased the alpha diversity of the mucosa-associated microbiota in the jejunum at the family and tended to increase at the genus level estimated with the Chao1 index. Therefore, it may correlate the microbiota results and a positive effect of the phytase supplementation observed on the overall ADG of the birds. Furthermore, the insoluble complexes formed with Ca with phytate [
24,
55] will interact in the intestinal lumen with inorganic P resulting in the formation of Ca-orthophosphate [
24,
56], which can lead to a decrease in the productive performance of the birds due to the decreased solubility and availability of the P [
24,
57]. The positive effects of phytase supplementation for broiler chickens are well established and are considered as an alternative for reducing the use of inorganic P, reducing P excretion in the environment, improving nutrient digestibility and utilization, and the growth performance [
20,
21,
58,
59].
The primary established role of phytase was to increase the bioavailability and utilization of phytate-bound P, nevertheless, it may also provide new insights into the anti-nutritive properties of phytate. The intestinal jejunal morphology parameters, such as villus height (VH), villus width (VW), crypt depth (CD), and villus height crypt depth ratio (VH:CD), can be interpreted as indicatives of intestinal health in broiler chickens [
60] since the intestinal jejunum is considered the main site of absorption in the small intestine [
61]. According to Nari et al. [
60], an increase in the villus height and VH:CD are linked to an enhanced epithelial cells turnover, and improvements on these parameters are correlated with activation of cell mitosis [
62] and greater nutrient absorption and utilization. Birds with longer villi may be an indicator of a greater absorptive capacity and mature enterocytes in the lumen [
63,
64], whereas birds with higher crypt depth may represent a high cell turnover, which can be considered a response to epithelial destruction and inflammation [
63,
65]. In the present study, phytase supplementation increased the VH and VW of broiler chickens, which are in agreement with other studies that reported positive effects of phytase on the intestinal morphology parameters [
60,
66]. The increased VH and CD may also be related to the positive modulation of the mucosa-associated microbiota observed in the present study. On the other hand, the intestinal morphology data of crypt depth indicated that a higher supplementation might have deleterious effects on the intestinal morphology, however, it might be indirectly correlated with the health status and wellbeing of the birds, as previously described by Paiva et al. [
63]. Supplementing phytase exogenously will express their positive effects regardless of the bird’s health status, although when a bird is not healthy their ability to digest and absorb nutrients will be compromised and consequently nutrients released by phytase will available for pathogenic bacteria leading to issues on the microbial population of the small intestine [
63,
67,
68]. The intestinal morphology results of the present study may be an indicator of greater capacity of digestion, absorption, and utilization of nutrients by the birds supplemented with increasing levels of phytase.
Phytase supplementation is also involved with protein and amino acid digestion and availability for the animal due to the formation of protein-phytate complexes. The pH can be determinant for the formation of those complexes, which was first reported by Breese Jones and Csonka [
69] and had that adverse effects for monogastric animals studied by Rojas and Scott [
70]. According to Selle et al. [
71], binary protein phytate-complexes are present at acidic pH, and ternary protein mineral phytate-complexes are formed via a cationic bridge as pH approaches neutrality. In theory, the hydrolysis of phytate will release phytate-bound proteins and P for animal utilization [
41,
71]. Phytase is capable of increasing the utilization of dietary amino acids and N by decreasing these antinutritional effects of the phytate molecule [
41]. In addition, a reduction in the dietary Ca may improve the protein and amino acid digestibility and utilization by facilitating a decrease in gastric pH and consequently improving the pepsin efficiency [
23,
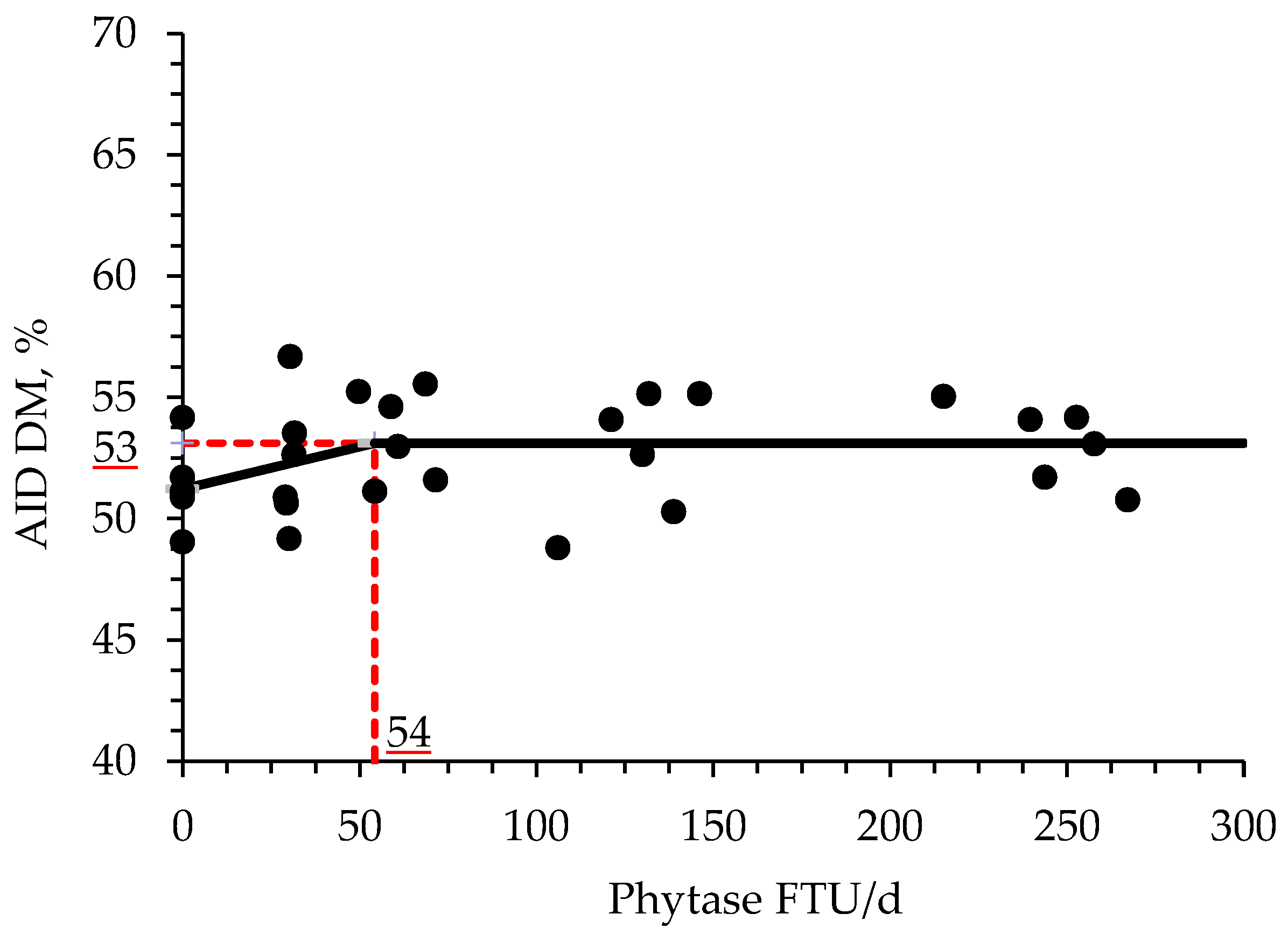
72]. The results of the present study are in agreement with these findings, where the AID of CP and P increased, whereas numerically increased the AID of Ca in broiler chickens supplemented with increasing levels of phytase. Moreover, it was found by broken-line analysis that the optimal supplemental levels of phytase for improving AID of CP, DM, and P are 33, 54, and 59 FTU/d or 541, 885, and 952 FTU/kg feed, respectively. During the preparation of enzymes, usually, the microorganisms used are genetically modified generating enzymatic side activities by the presence of other enzymes, such as proteases, in their structure [
41]. Based on Selle et al. [
71] findings, any enzymatic side activities present in the phytase structure are secondary, nevertheless, they may increase the hydrolysis of the phytate molecule, either by their own phosphoric effects or by improving the accessibility of phytase to its substrate.
Furthermore, phytase supplementation also shows potential benefits regarding bone parameters, such as the breaking strength, minerals, and ash content for monogastric animals, especially broiler chickens [
14,
22,
57,
59,
73,
74]. According to Sebastian et al. [
75], the improvement of the ash percentage in the tibia may indicate an increase in bone mineralization consequent to an increase in mineral availability released by phytase from the phytate-mineral complexes. The results of the present study showed that the bone-breaking strength, ash, and P content of the bone increased, whereas the Ca content numerically increase in broiler chickens supplemented with increasing levels of phytase. Increasing the mineral content of the bone, especially P content, may reflect on a greater breaking strength indicating that phytase plays an important role in bone mineralization. These positive effects can be attributed to the efficacy of phytase in hydrolyzing the phytate molecule and consequently releasing more P and other nutrients bound to the phytate complexes.
Supplemental phytase can provide benefits that go beyond improving nutrient digestibility, bone parameters, and growth performance. In this study, the supplementation of increasing levels of phytase played an important role in the intestinal health of broiler chickens by potentially reducing pathogenic and increasing beneficial bacteria, which in turn may reflect in benefits on nutrient digestibility, intestinal morphology, and bone parameters. The supplementation of phytase may be an interesting alternative for improving intestinal health and subsequent performance of broiler chickens.







{kind=link}
{kind=link}
{kind=link}