Induction of Spermiation in Sterlet Acipenser ruthenus by PLGA Microparticle Delivery with Sustained Alarelin Release


 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish Rearing and Pre-Spawning Water Conditions
2.2. Treatment
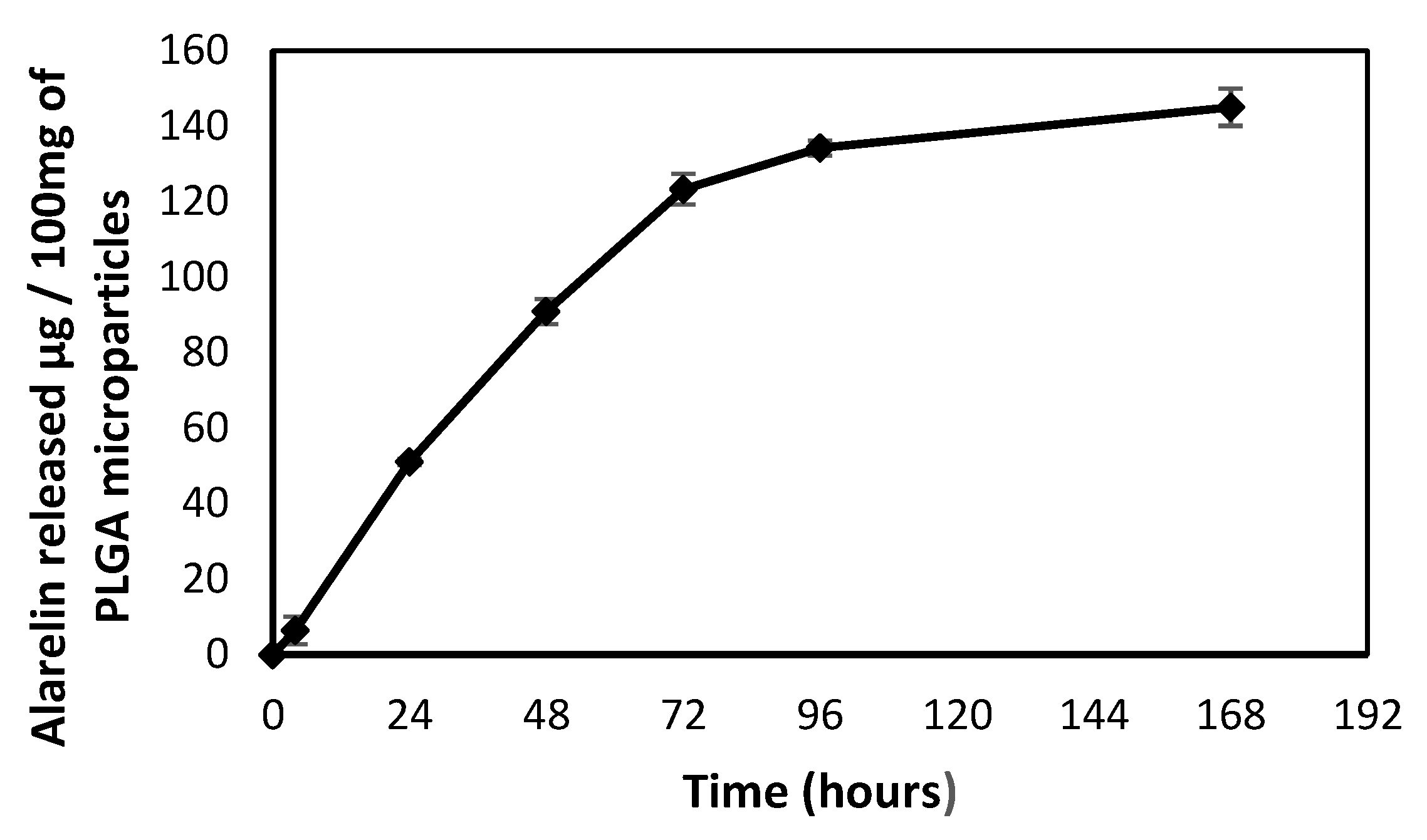
2.2.1. Synthesis of PLGA Microparticles
2.2.2. Treatments
2.3. Sample Collection
2.3.1. Blood Collection
2.3.2. Milt Collection
2.4. Analysis of Samples
2.4.1. 11-Ketotestosterone and Testosterone Analysis
2.4.2. Sperm Quantitative Parameters
2.4.3. Sperm Qualitative Parameters
2.5. Statistical Analysis
2.5.1. Spermiation Rate
2.5.2. Quantitative Sperm Parameters and Androgen Concentrations
2.5.3. Sperm Qualitative Parameters
3. Results
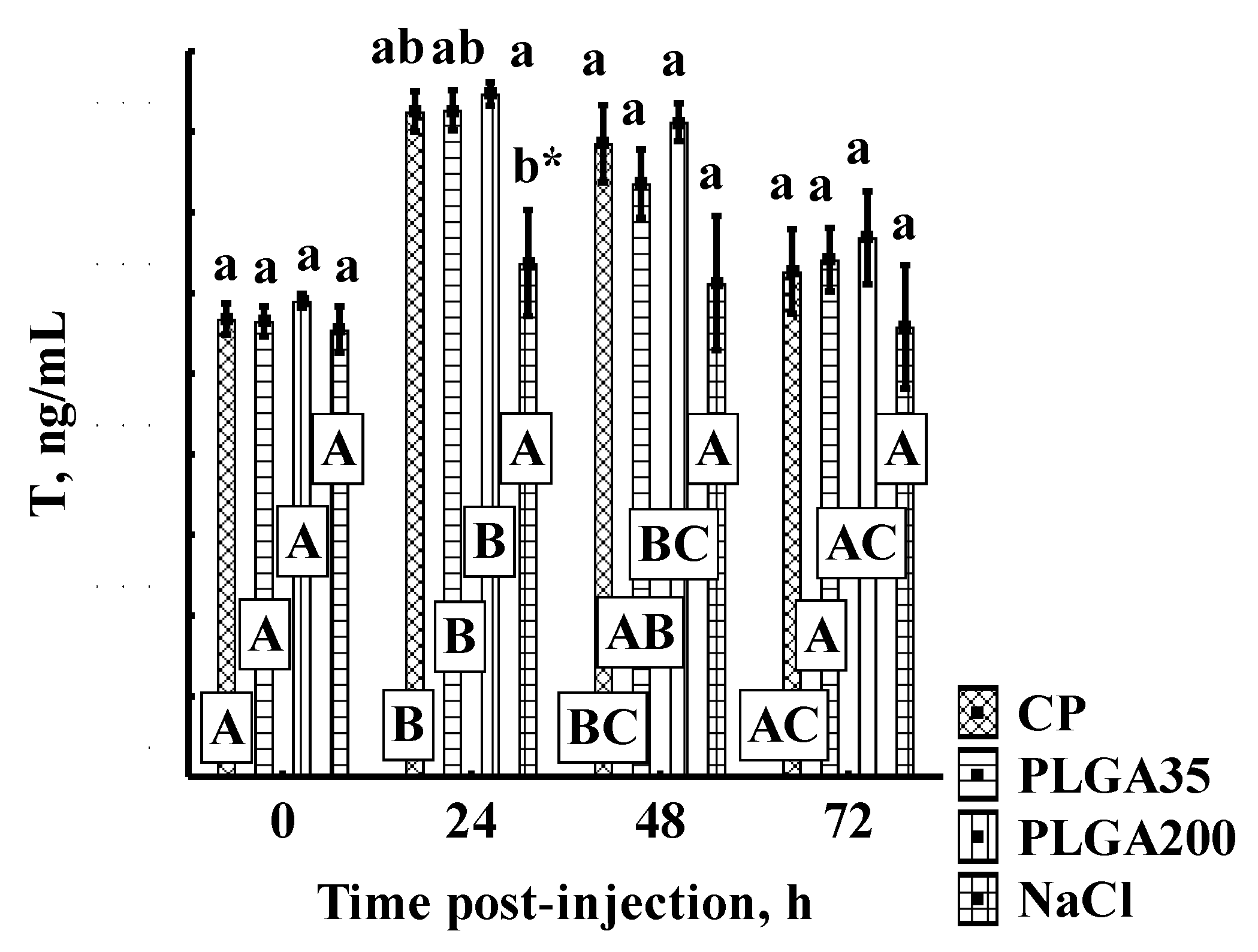
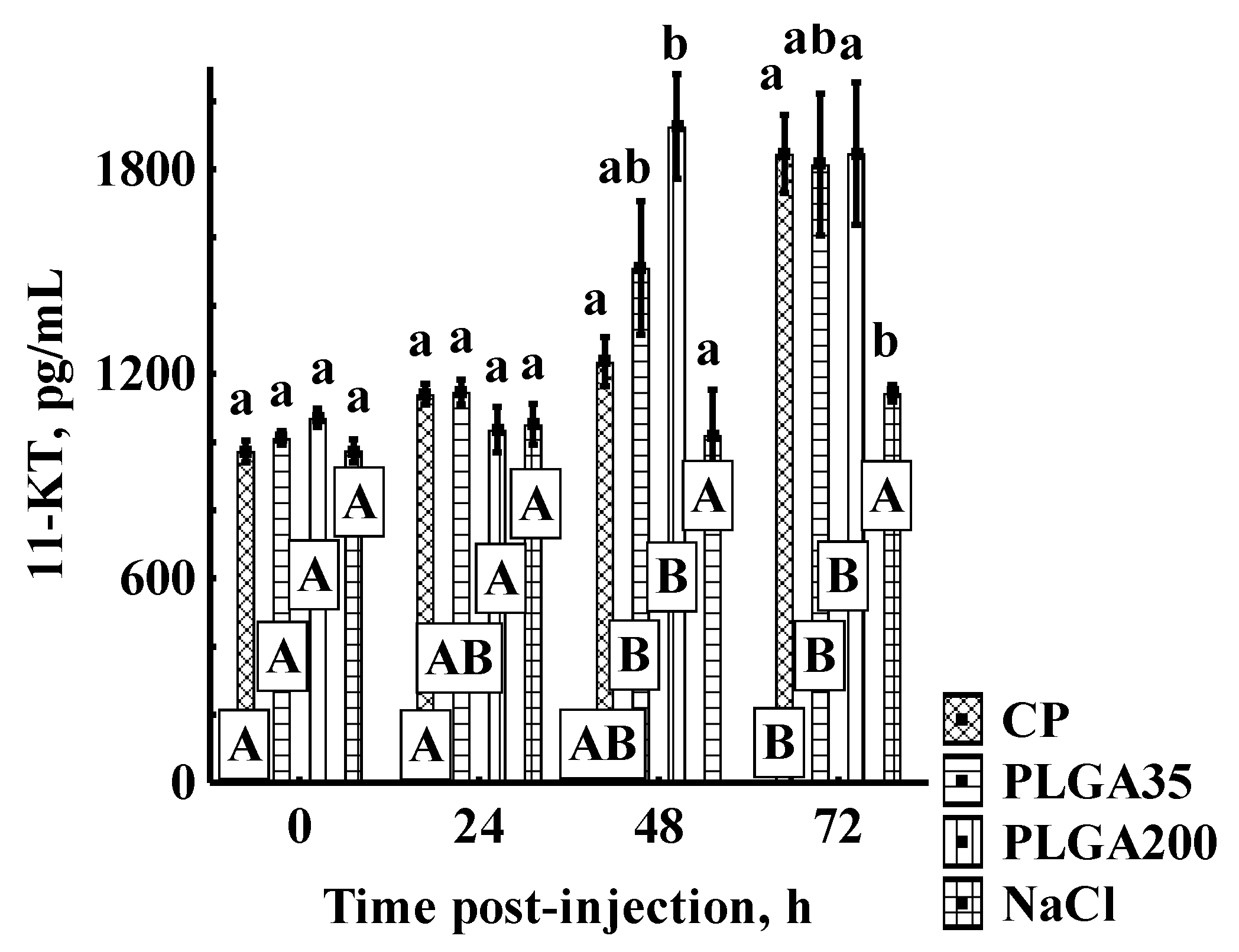
3.1. Androgen Concentrations
3.1.1. Testosterone Concentrations
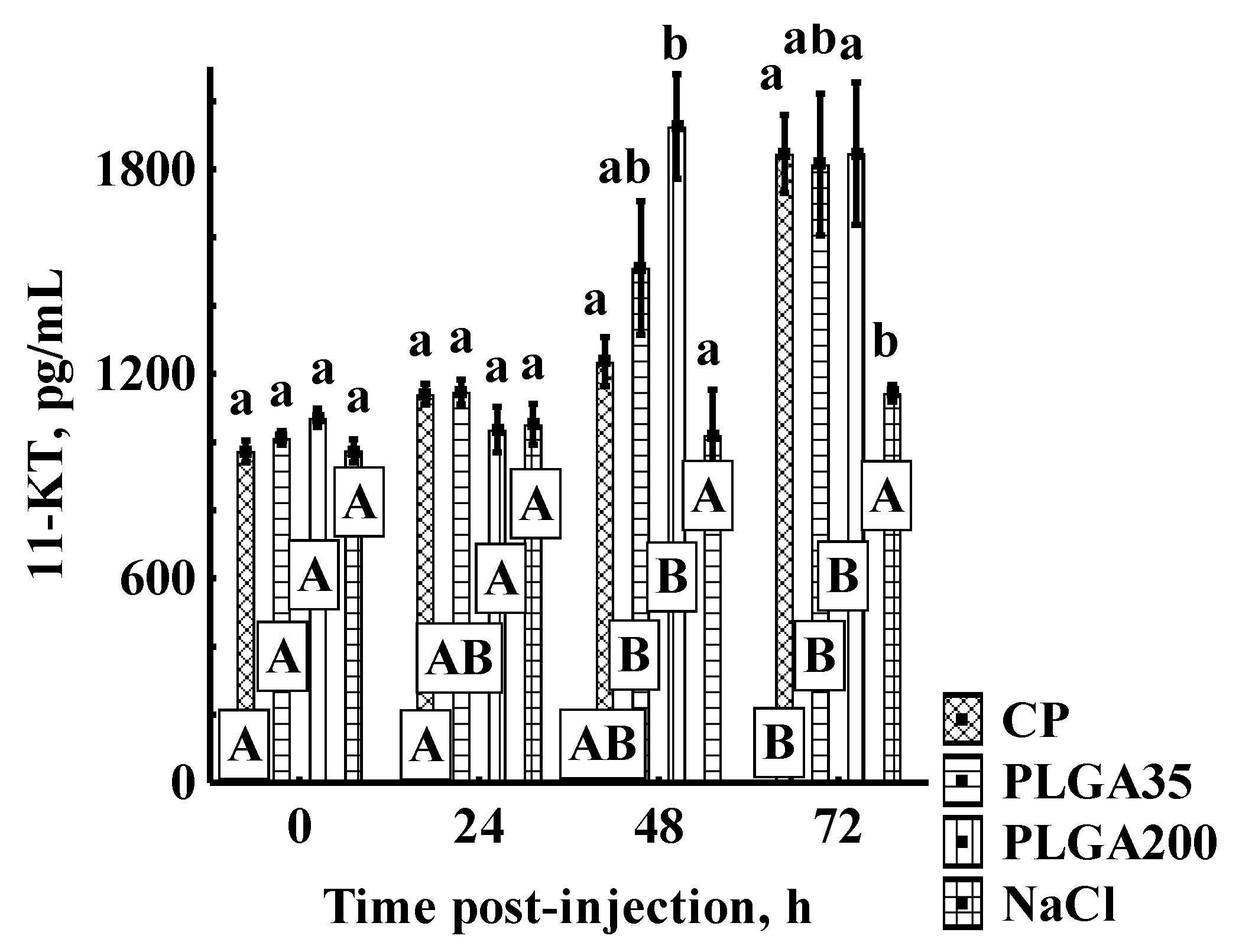
3.1.2. 11-KT Concentrations
3.2. Effect of Hormone Treatment on Sperm Production
3.2.1. Spermiation Rate
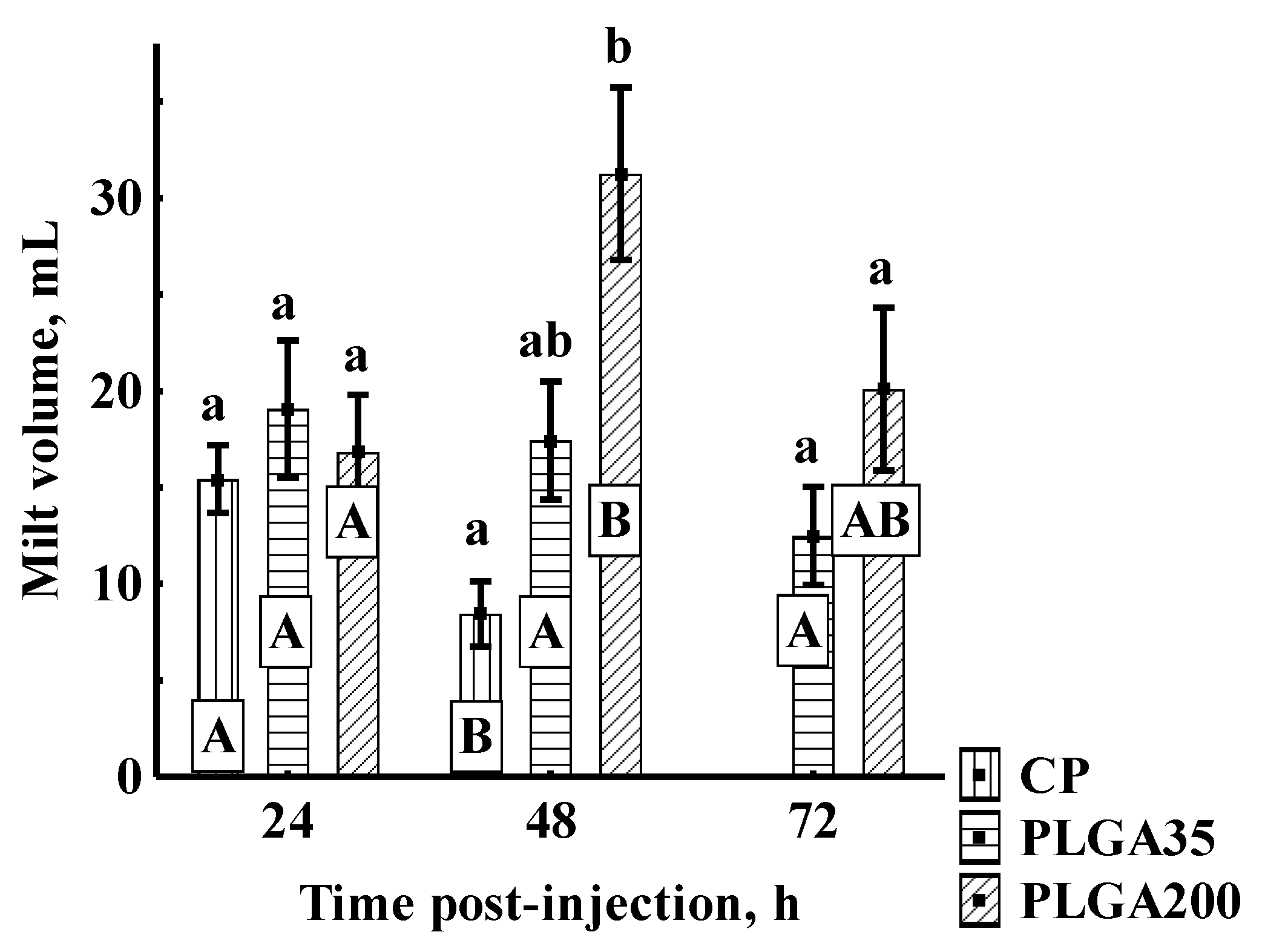
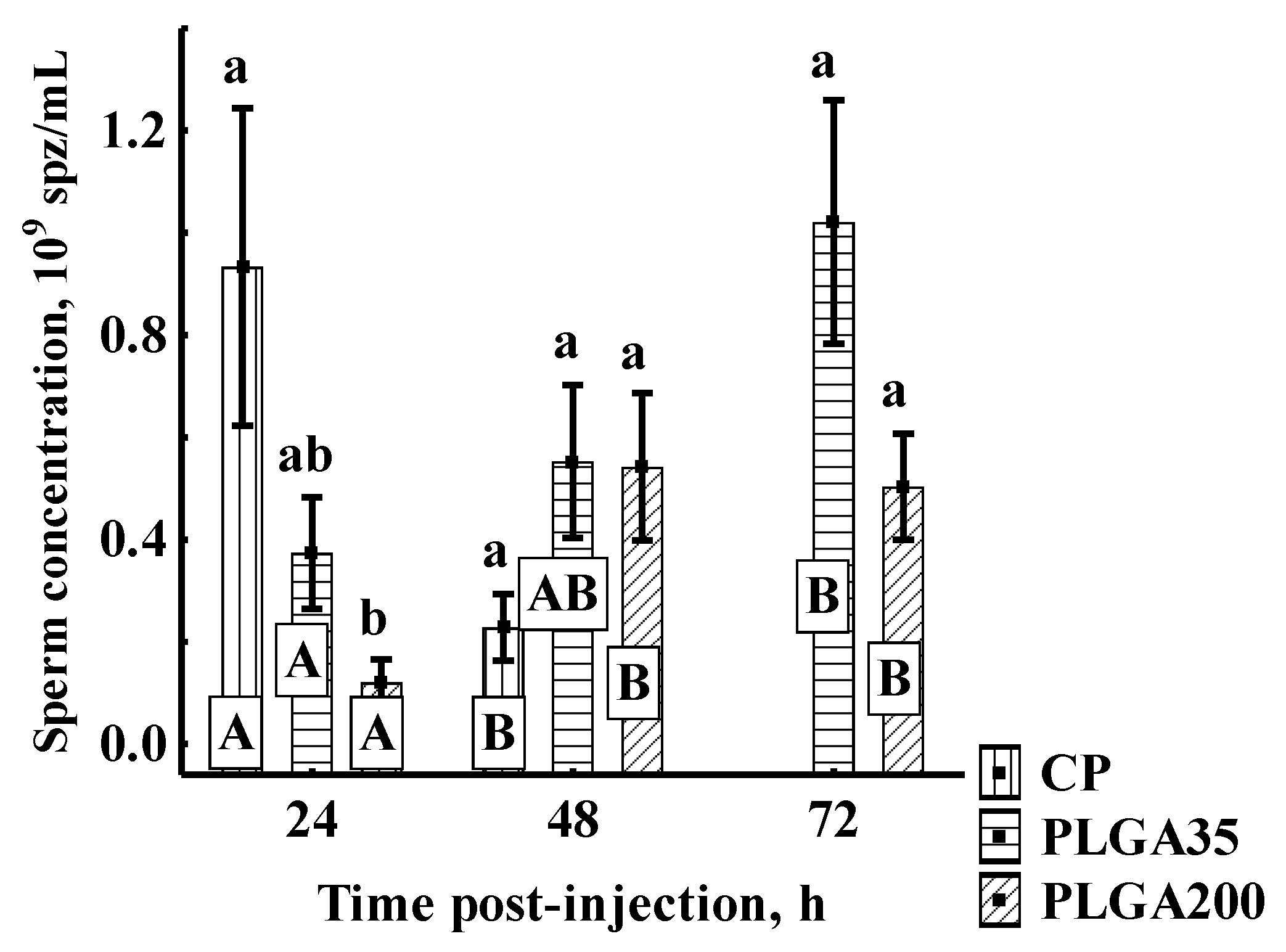
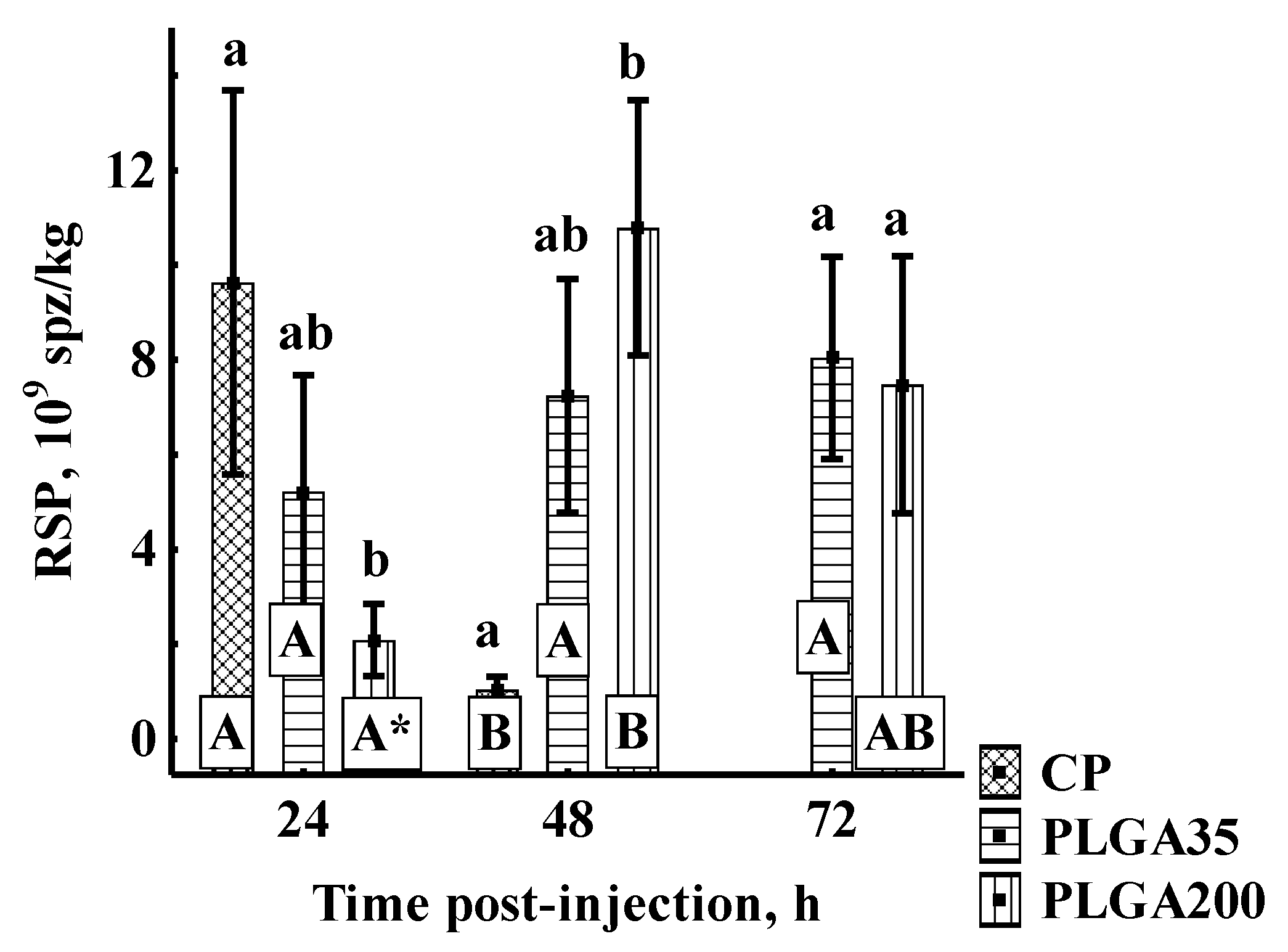
3.2.2. Sperm Quantitative Parameters
Milt Volume
Spermatozoon Concentration
Relative Sperm Production
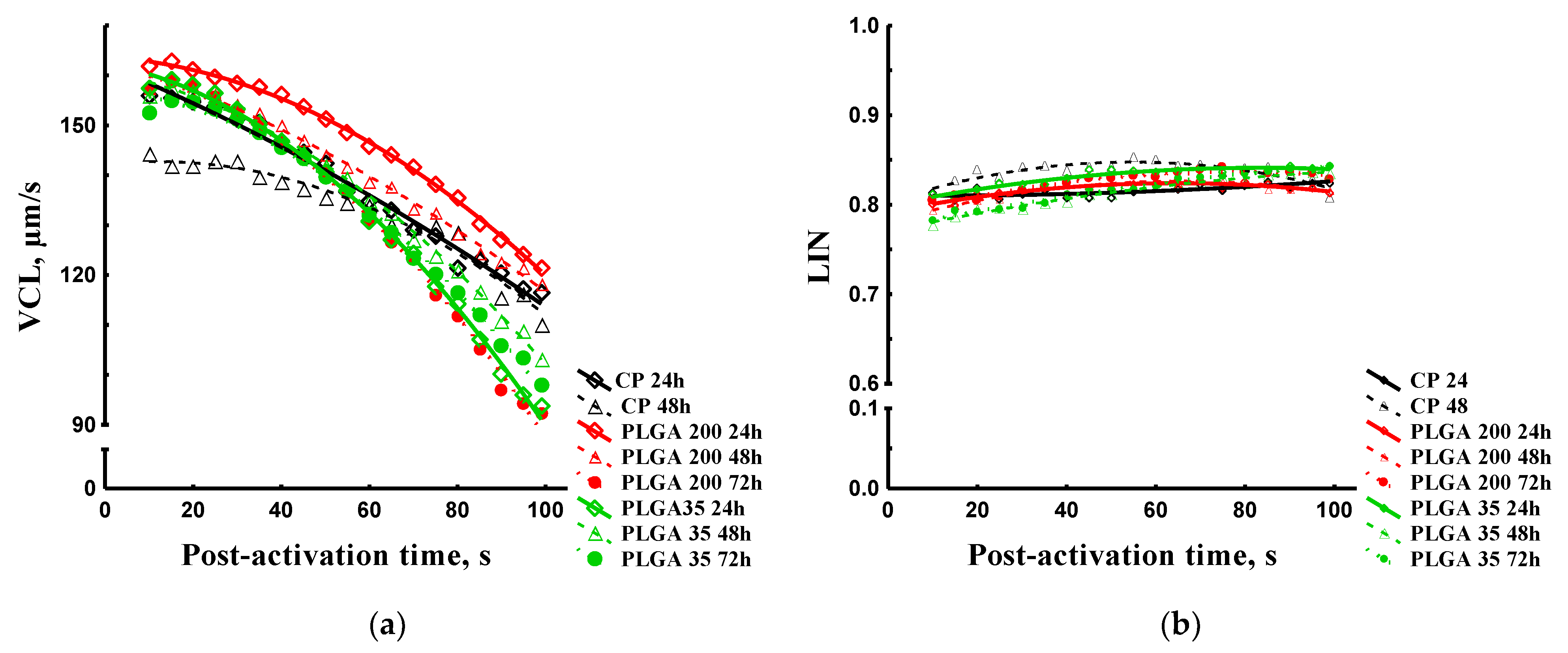
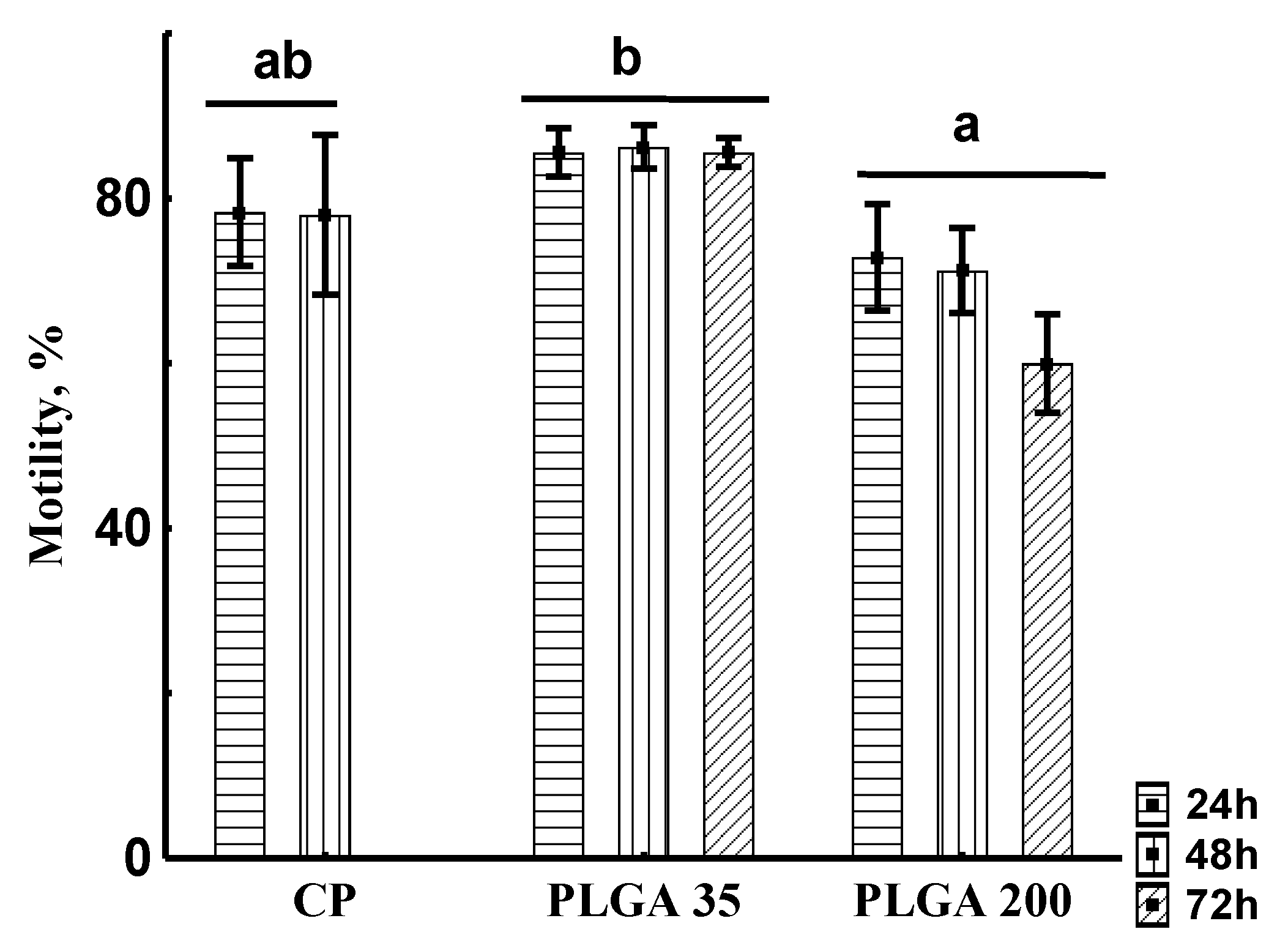
3.2.3. Sperm Qualitative Parameters
Spermatozoon Curvilinear Velocity and Linearity
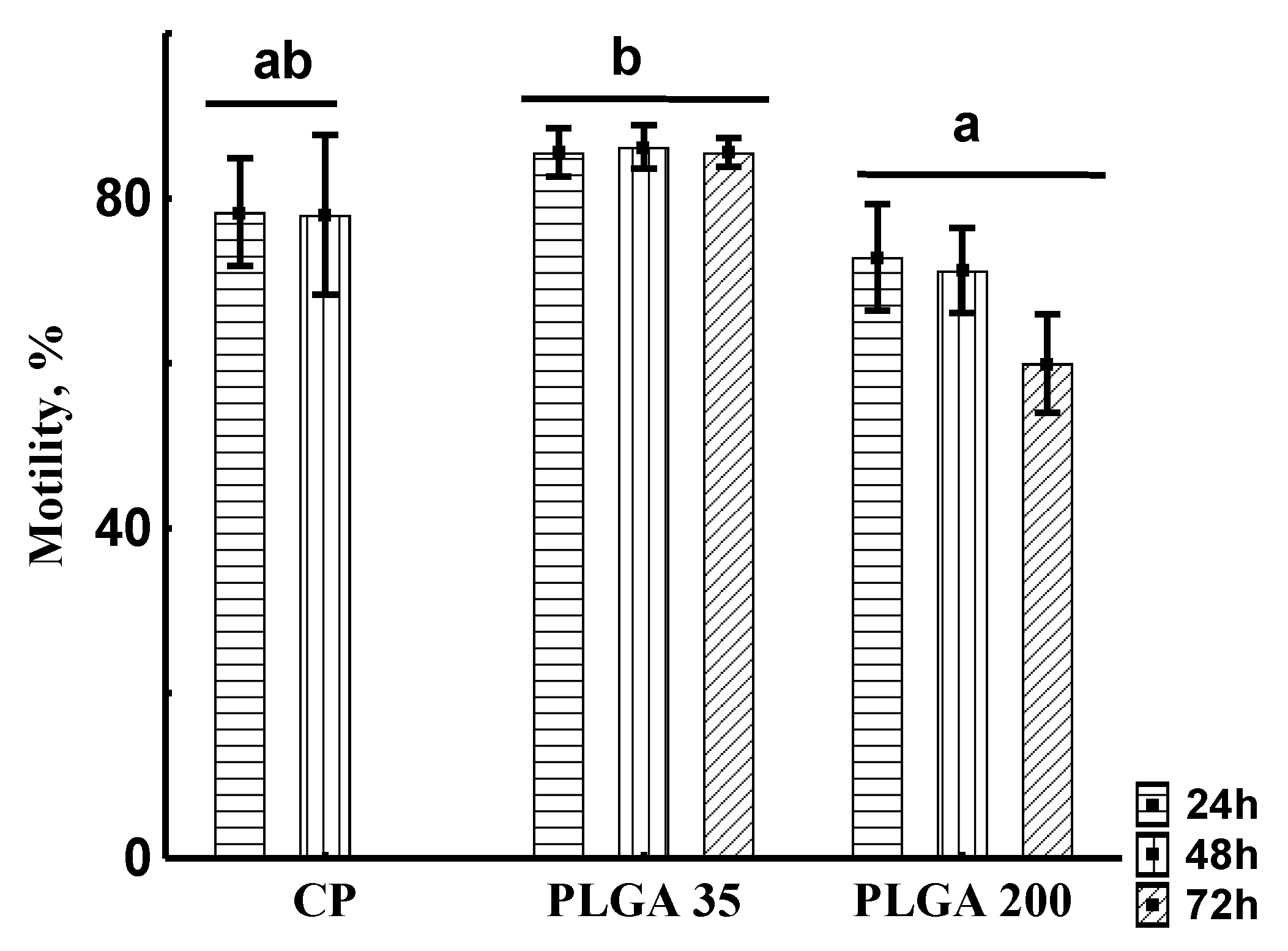
Spermatozoon Motility Rate
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Birstein, V.J.; DeSalle, R. Molecular phylogeny of Acipenserinae. Mol. Phylogenet. Evol. 1998, 9, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Bronzi, P.; Rosenthal, H.; Gessner, J. Global sturgeon aquaculture production: An overview. J. Appl. Ichthyol. 2011, 27, 169–175. [Google Scholar] [CrossRef]
- Podhorec, P.; Kouril, J. Induction of final oocyte maturation in Cyprinidae fish by hypothalamic factors: A review. Vet. Med. 2009, 54, 97–110. [Google Scholar] [CrossRef] [Green Version]
- Alavi, S.M.H.; Hatef, A.; Mylonas, C.C.; Gela, D.; Papadaki, M.; Rodina, M.; Kaspar, V.; Psenicka, M.; Podhorec, P.; Linhart, O. Sperm characteristics and androgens in Acipenser ruthenus after induction of spermiation by carp pituitary extract or GnRHa implants. Fish. Physiol. Biochem. 2012, 38, 1655–1666. [Google Scholar] [CrossRef]
- Yaron, Z.; Bogomolnaya, A.; Drori, S.; Biton, I.; Aizen, J.; Kulikovsky, Z.; Levavi-Sivan, B. Spawning Induction in the Carp: Past Experience and Future Prospects—A Review. Isr. J. Aquac.-Bamid 2009, 61, 5–26. [Google Scholar]
- Burgus, R.; Butcher, M.; Ling, N.; Monahan, M.; Rivier, J.; Fellows, R.; Amoss, M.; Blackwell, R.; Vale, W.; Guillemin, R. Molecular structure of the hypothalamic factor (LRF) of ovine origin monitoring the secretion of pituitary gonadotropic hormone of luteinization (LH). C. R. Acad. Hebd. Seances Acad. Sci. D 1971, 273, 1611–1613. [Google Scholar]
- Mañanós, E.; Duncan, N.; Mylonas, C. Reproduction and control of ovulation, spermiation and spawning in cultured fish. In Methods in Reproductive Aquaculture. Marine and Freshwater Species; Cabrita, E., Robles, V., Herráez, P., Eds.; CRC Press: Boca Raton, FL, USA, 2008; pp. 3–80. [Google Scholar]
- Zohar, Y.; Mylonas, C.C. Endocrine manipulations of spawning in cultured fish: From hormones to genes. Aquaculture 2001, 197, 99–136. [Google Scholar] [CrossRef]
- Williot, P.; Brun, R.; Rouault, T.; Pelard, M.; Mercier, D.; Ludwig, A. Artificial spawning in cultured sterlet sturgeon, Acipenser ruthenus L., with special emphasis on hermaphrodites. Aquaculture 2005, 246, 263–273. [Google Scholar] [CrossRef]
- Mohammadzadeh, S.; Yeganeh, S.; Moradian, F.; Milla, S.; Falahatkar, B. Spawning induction in Sterlet sturgeon (Acipenser ruthenus) with recombinant GnRH: Analysis of hormone profiles and spawning indices. Aquaculture 2021, 533, 736108. [Google Scholar] [CrossRef]
- Mylonas, C.C.; Zohar, Y. Use of GnRHa-delivery systems for the control of reproduction in fish. Rev. Fish. Biol. Fisher 2001, 10, 463–491. [Google Scholar] [CrossRef]
- Mañanós, E.; Carrillo, M.; Sorbera, L.A.; Mylonas, C.C.; Asturiano, J.F.; Bayarri, M.J.; Zohar, Y.; Zanuy, S. Luteinizing hormone and sexual steroid plasma levels after treatment of European sea bass with sustained-release delivery systems for gonadotropin-releasing hormone analogue. J. Fish. Biol. 2002, 60, 328–339. [Google Scholar] [CrossRef]
- Amini, K.; Siraj, S.S.; Amiri, B.M.; Rostami, S.; Sharr, A.; Hossienzadeh, H. Evaluation of LHRH-a acute release implantation on final maturation and spawning in not-fully matured broodstocks of Persian sturgeon (Acipenser persicus Borodin, 1897). Iran. J. Fish. Sci. 2012, 11, 440–459. [Google Scholar]
- Arabacı, M.; Diler, İ.; Sarı, M. Induction and synchronisation of ovulation in rainbow trout, Oncorhynchus mykiss, by administration of emulsified buserelin (GnRHa) and its effects on egg quality. Aquaculture 2004, 237, 475–484. [Google Scholar] [CrossRef]
- Knowles, J.C.; Vyslov~il, J.; Muselík, J.; Stejskal, V.; Kouril, J.; Podhorec, P. Efficacy of poly (lactic-co-glycolic acid) microparticles as a gonadotropin-releasing hormone analogue delivery system to stimulate ovulation of peled Coregonus peled. Czech. J. Anim. Sci. 2021, 66, 331–338. [Google Scholar] [CrossRef]
- Rainis, S.; Mylonas, C.C.; Kyriakou, Y.; Divanach, P. Enhancement of spermiation in European sea bass (Dicentrarchus labrax) at the end of the reproductive season using GnRHa implants. Aquaculture 2003, 219, 873–890. [Google Scholar] [CrossRef]
- Lim, H.K.; Pankhurst, N.W.; Fitzgibbon, Q.P. Effects of slow release gonadotropin releasing hormone analog on milt characteristics and plasma levels of gonadal steroids in greenback flounder, Rhombosolea tapirina. Aquaculture 2004, 240, 505–516. [Google Scholar] [CrossRef]
- Han, F.Y.; Thurecht, K.J.; Whittaker, A.K.; Smith, M.T. Bioerodable PLGA-Based Microparticles for Producing Sustained-Release Drug Formulations and Strategies for Improving Drug Loading. Front. Pharmacol. 2016, 7, 185. [Google Scholar] [CrossRef] [Green Version]
- Rónyai, A. Effect of different synthetic gonadotrop-releasing hormone analogues and their combinations with an anti-dopaminergic compound on the reproduction performance of sterlet (Acipenser ruthenus L.). Aquac. Res. 2009, 40, 315–321. [Google Scholar] [CrossRef]
- Purchase, C.F.; Earle, P.T. Modifications to the imagej computer assisted sperm analysis plugin greatly improve efficiency and fundamentally alter the scope of attainable data. J. Appl. Ichthyol. 2012, 28, 1013–1016. [Google Scholar] [CrossRef]
- Shaliutina, A.; Dzyuba, B.; Hulak, M.; Boryshpolets, S.; Li, P.; Linhart, O. Evaluation of Spermiation Indices with Multiple Sperm Collections in Endangered Sterlet (Acipenser ruthenus). Reprod. Domest. Anim. 2012, 47, 479–484. [Google Scholar] [CrossRef]
- Linhart, O.; Mims, S.D.; Gomelsky, B.; Hiott, A.E.; Shelton, W.L.; Cosson, J.; Rodina, M.; Gela, D. Spermiation of paddlefish (Polyodon spathula, Acipenseriformes) stimulated with injection of LHRH analogue and carp pituitary powder. Aquat. Living Resour. 2000, 13, 455–460. [Google Scholar] [CrossRef]
- Mylonas, C.C.; Duncan, N.J.; Asturiano, J.F. Hormonal manipulations for the enhancement of sperm production in cultured fish and evaluation of sperm quality. Aquaculture 2017, 472, 21–44. [Google Scholar] [CrossRef]
- Rzemieniecki, A.; Domagałła, J.; Glogowski, J.; Ciereszko, A.; Trzebiatowski, R.; Kouřil, J.; Hamáčkova, J.; Babiak, I. Induced spermiation in 3-year-old sterlet, Acipenser ruthenus L. Aquac. Res. 2004, 35, 144–151. [Google Scholar] [CrossRef]
- Zohar, Y.; Goren, A.; Tosky, M.; Pagelson, G.; Leibovitz, D.; Koch, Y. The bioactivity of gonadotropin-releasing hormones and its regulation in the gilthead seabream, Sparus aurata-in vivo and In vitro studies. Fish. Physiol. Biochem. 1989, 7, 59–67. [Google Scholar] [CrossRef]
- Fernandez-Palacios, H.; Schuchardt, D.; Roo, J.; Izquierdo, M.; Hernandez-Cruz, C.; Duncan, N. Dose-dependent effect of a single GnRHa injection on the spawning of meagre (Argyrosomus regius) broodstock reared in captivity. Span. J. Agric. Res. 2014, 12, 11. [Google Scholar] [CrossRef] [Green Version]
- Podhorec, P.; Socha, M.; Ammar, I.B.; Sokolowska-Mikolajczyk, M.; Brzuska, E.; Milla, S.; Gosiewski, G.; Stejskal, V.; Simko, M.; Kouril, J. The effects of GnRHa with and without dopamine antagonist on reproductive hormone levels and ovum viability in tench Tinca tinca. Aquaculture 2016, 465, 158–163. [Google Scholar] [CrossRef]
- Schulz, R.W.; de França, L.R.; Lareyre, J.-J.; LeGac, F.; Chiarini-Garcia, H.; Nobrega, R.H.; Miura, T. Spermatogenesis in fish. Gen. Comp. Endocr. 2010, 165, 390–411. [Google Scholar] [CrossRef]
- Miura, T.; Yamauchi, K.; Takahashi, H.; Nagahama, Y. The role of hormones in the acquisition of sperm motility in salmonid fish. J. Exp. Zool. 1992, 261, 359–363. [Google Scholar] [CrossRef]
- Nagahama, Y. Endocrine regulation of gametogenesis in fish. Int. J. Dev. Biol. 2002, 38, 217–229. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Fish Weight (kg) | Dose |
|---|---|---|
| 0.9% NaCl | 1.57 ± 0.23 | 1 mL/kg |
| Carp pituitary extract | 1.59 ± 0.22 | 4 mg/kg |
| PLGA35 1 | 1.59 ± 0.25 | 35 µg/kg Alarelin |
| PLGA200 1 | 1.58 ± 0.27 | 200 µg/kg Alarelin |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Podhorec, P.; Knowles, J.; Vysloužil, J.; Boryshpolets, S.; Kubová, K.; Rodina, M.; Kholodnyy, V.; Sotnikov, A.; Gela, D.; Dzyuba, B. Induction of Spermiation in Sterlet Acipenser ruthenus by PLGA Microparticle Delivery with Sustained Alarelin Release. Animals 2021, 11, 3305. https://doi.org/10.3390/ani11113305
Podhorec P, Knowles J, Vysloužil J, Boryshpolets S, Kubová K, Rodina M, Kholodnyy V, Sotnikov A, Gela D, Dzyuba B. Induction of Spermiation in Sterlet Acipenser ruthenus by PLGA Microparticle Delivery with Sustained Alarelin Release. Animals. 2021; 11(11):3305. https://doi.org/10.3390/ani11113305
Chicago/Turabian StylePodhorec, Peter, Jindřiška Knowles, Jakub Vysloužil, Sergii Boryshpolets, Kateřina Kubová, Marek Rodina, Vitaliy Kholodnyy, Anatolii Sotnikov, David Gela, and Borys Dzyuba. 2021. "Induction of Spermiation in Sterlet Acipenser ruthenus by PLGA Microparticle Delivery with Sustained Alarelin Release" Animals 11, no. 11: 3305. https://doi.org/10.3390/ani11113305
APA StylePodhorec, P., Knowles, J., Vysloužil, J., Boryshpolets, S., Kubová, K., Rodina, M., Kholodnyy, V., Sotnikov, A., Gela, D., & Dzyuba, B. (2021). Induction of Spermiation in Sterlet Acipenser ruthenus by PLGA Microparticle Delivery with Sustained Alarelin Release. Animals, 11(11), 3305. https://doi.org/10.3390/ani11113305