3.7.2. Hynobius notialis sp. nov. Min and Borzée, 2020
Synonymy:
“
Hynobius leechii (partim): [
72]: p. 33; [
73]: p. 20”.
“
Hynobius quelpaertensis (partim): [
52]: p. 1166”.
“
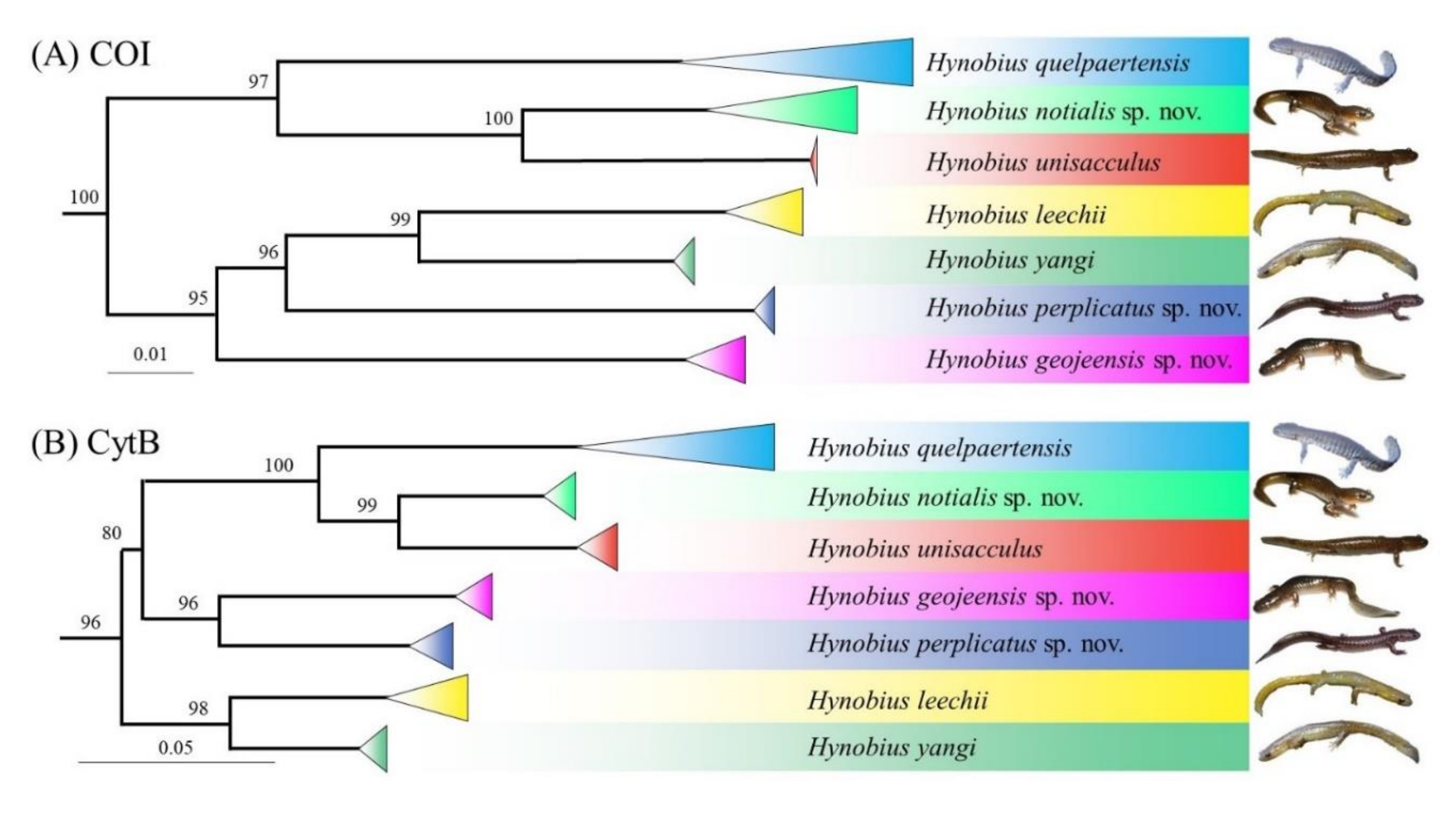
Hynobius leechii Clade HC1: [
53]: pp. 25–32; [
54]: p. 108; [
42]: pp. 475–503; [
55]: pp. 165–178”.
● Holotype
Voucher CGRB15873 (deposited in the Conservation Genome Resources Bank for Korean Wildlife (CGRB), College of Veterinary Medicine, Seoul National University; field ID mms3897); adult collected by Mi-Sook Min, HaeJun Baek and Dong Youn Kim on 14 March 2012 in Jangmok-myeon, Geoje-si, Gyeongsang Nam-do, Republic of Korea (34.986877° N, 128.682033° E;
Figure 8). Measurements and counts in
Table 9.
● Paratypes
Vouchers CGRB15870, CGRB15859, CGRB15884 (deposited in the Conservation Genome Resources Bank for Korean Wildlife (CGRB), College of Veterinary Medicine, Seoul National University; field IDs mms3894, mms3136 and mms4036 respectively). CGRB15870 adult collected by Mi-Sook Min on 14 March 2012 in Jangmok-myeon, Geoje-si, Gyeongsang Nam-do, Republic of Korea (34.986877° N, 128.682033° E). CGRB15859 adult collected by Mi-Sook Min, HaeJun Baek and Dong Yoon Kim on 5 April 2011 in Sicheon-myeon, Sancheong-gun, South Gyeongsang Province, Republic of Korea (35.27849° N, 127.840687° E). CGRB15884 adult collected by Mi-Sook Min, HaeJun Baek and Dong Youn Kim on 21 March 2012 in Jisu-myeon, Jinju-si, Gyeongsang Nam-do, Republic of Korea (35.235064° N, 128.269611° E). Measurements and counts in
Table 9 illustrations in
Figure S3.
● Etymology
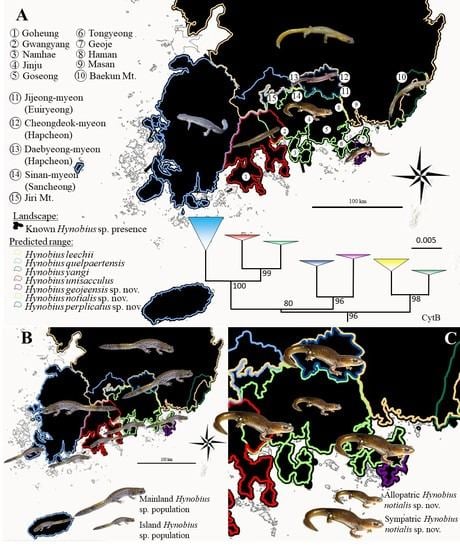
The species was first found in Tongyeong and Namhae (Graphical Abstract), in the central costal region of the Republic of Korea. The name H. notialis sp. nov. comes from the southern location of the holotype, and the majority of the species’ range. The vernacular name of the species, Southern Korean salamander, reflects the scientific name of the special, as does its Korean name: 남방도롱뇽 (pronounced: Nambang Dorongnyong).
● Identity, diagnosis and distribution
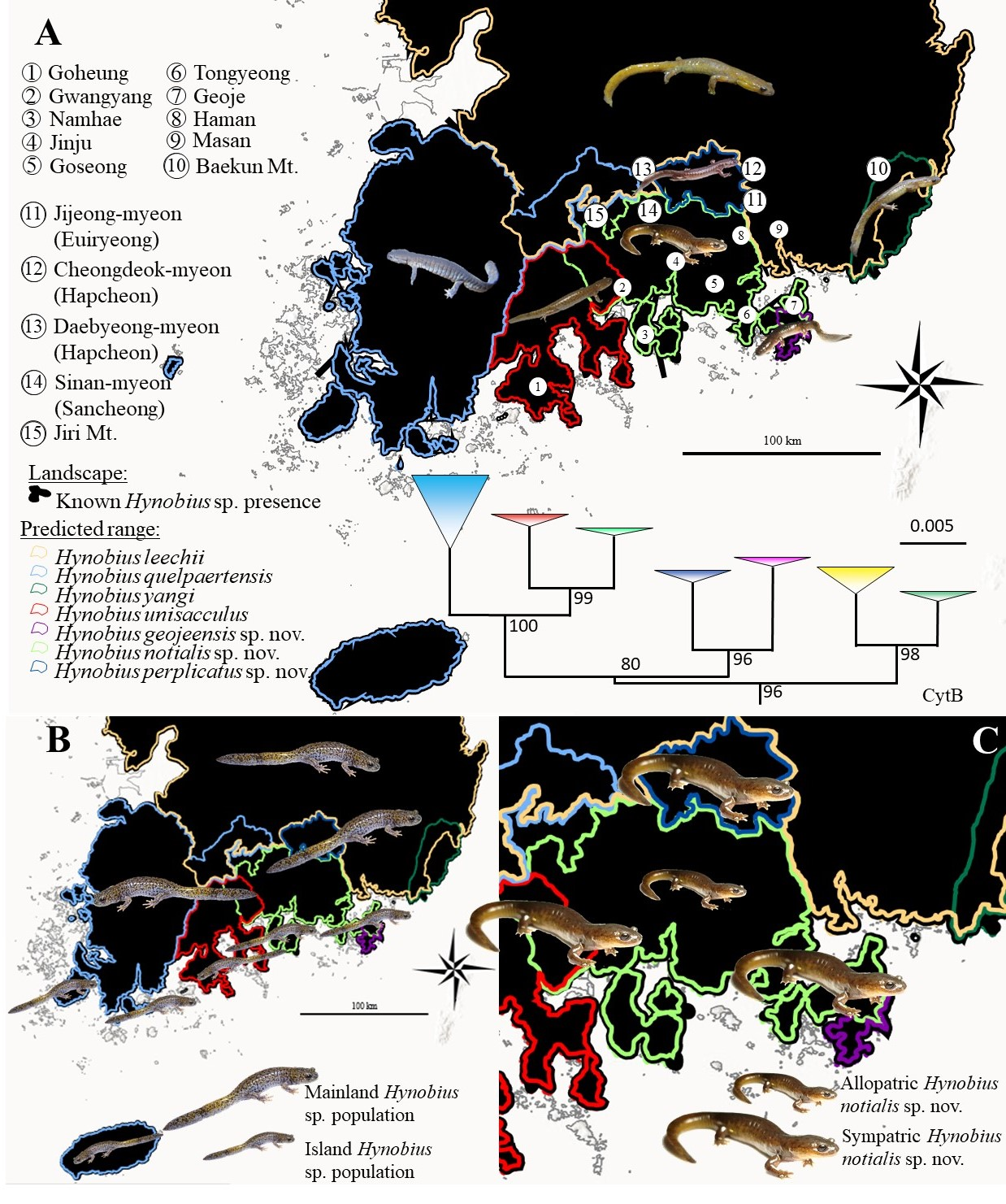
To date, the species is known to range from the feet of Jiri Mountain on the north west, Gwangyang on the south west, Masan on the south east and Haman on the north east (Graphical Abstract). The species is currently known to be present on islands of Namhae, Changseong and Geoje. This species is sympatric with
H. geojeensis sp. nov. on Geoje island, with
H. unisacculus on Baekun Mountain north of Gwangyang and with
H. perplicatus along the southern range of that species (
Figure 1 and Graphical Abstract). Sympatry with the other Korean
Hynobius species has so far not been demonstrated but is not impossible.
The species identification is primarily based on range, although the species is sympatric with
H. perplicatus sp. nov.,
H. geojeensis sp. nov. and
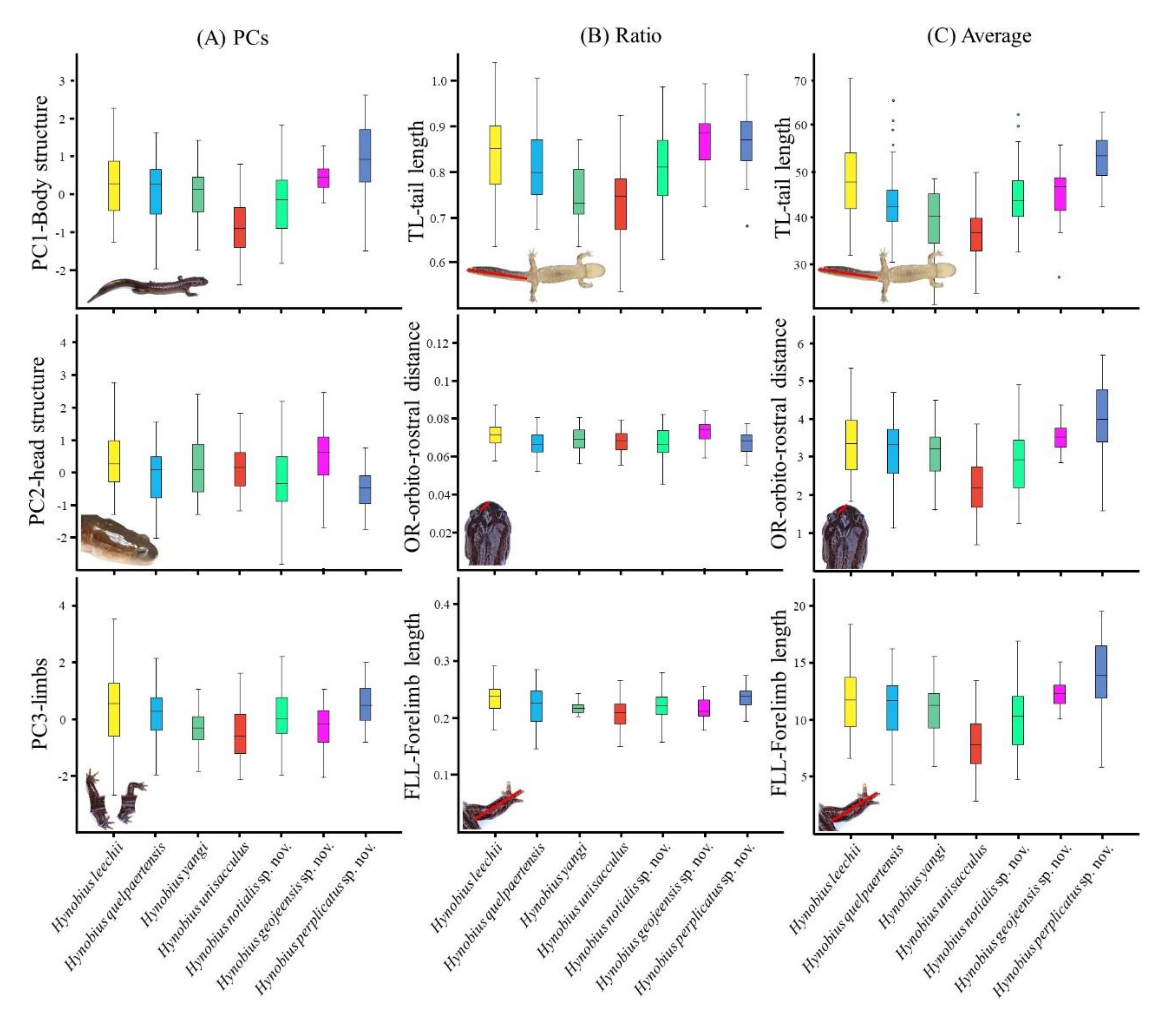
H. unisacculus. The species is generally larger than
H. unisacculus and smaller than
H. perplicatus sp. nov. (
Table 5 and
Table 9). However, size strongly overlaps with that of
H. geojeensis sp. nov., despite a lower average (
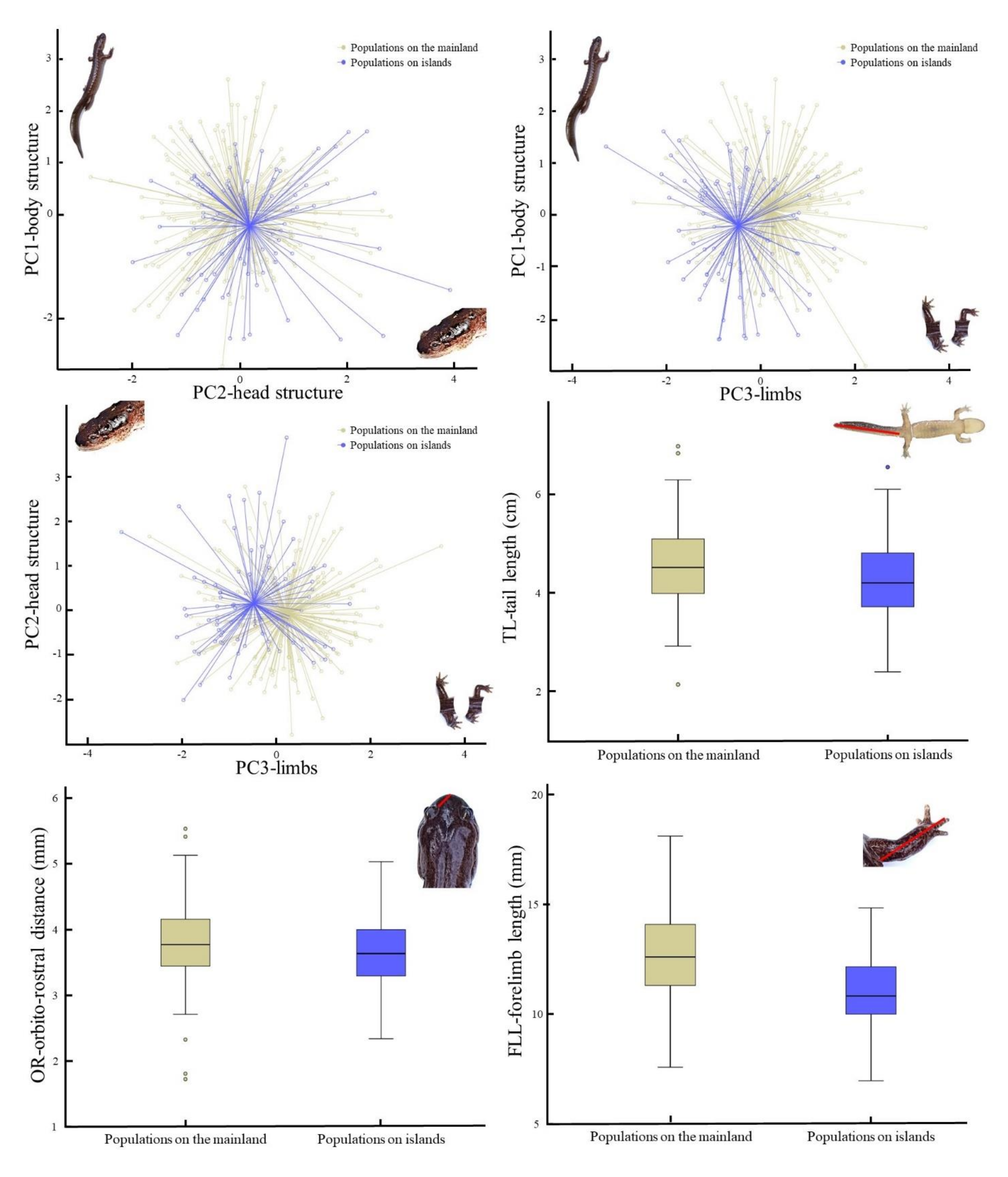
Figure 2;
Figures S1 and S2). When in sympatry with
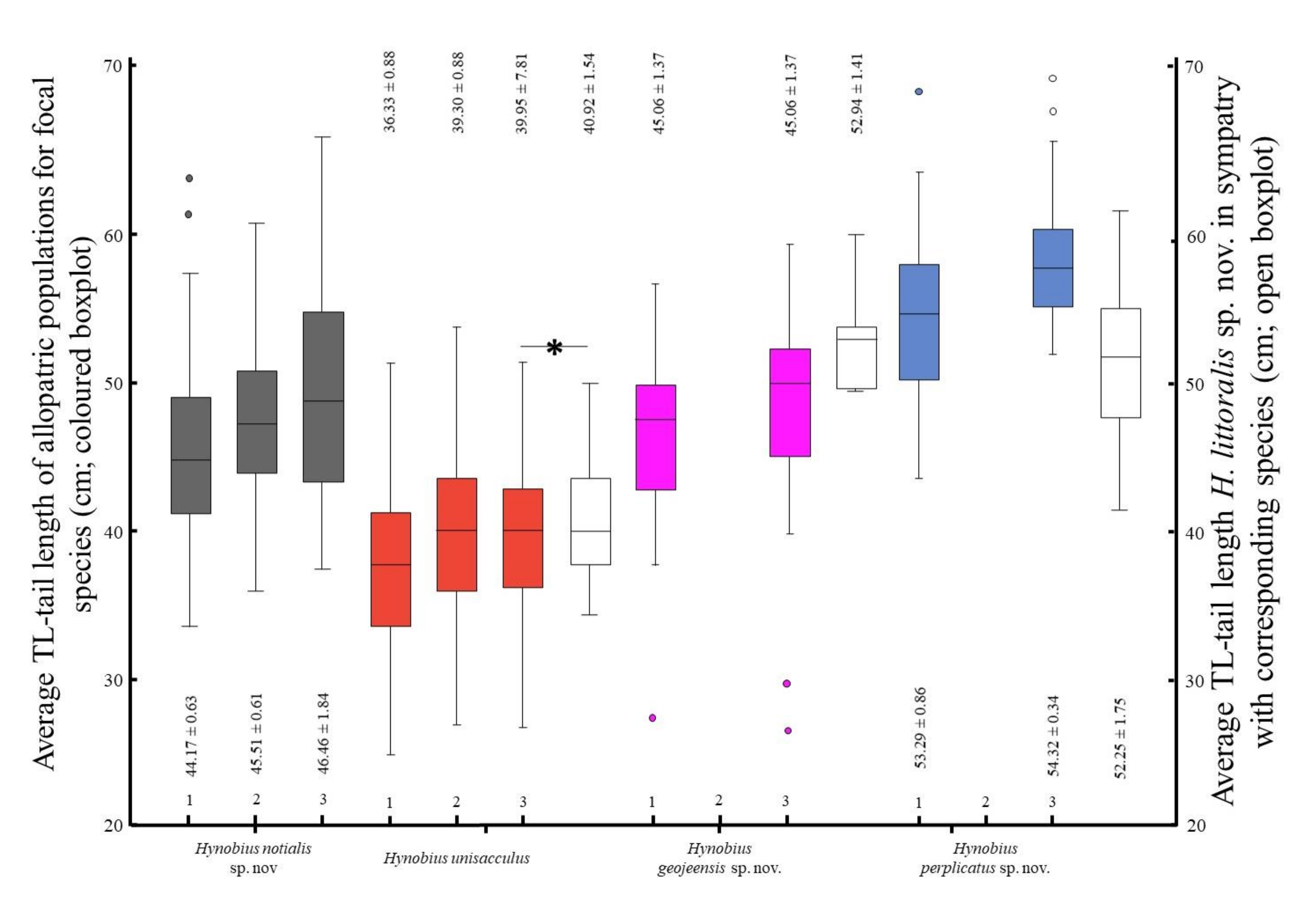
H. unisacculus, the species can be identified if (in mm): SVL > 61.29, TL > 50.15, GA > 31.92, CW > 10. 72, HL > 14.00, HW > 10.83 (
Table 5). For identification when in sympatry with the two other species described in this work, please refer to the matching species description. The species breeds in natural and agricultural wetlands (
Figure 9).
● ZooBank registration
We hereby state that the present paper has been registered to the Official Register of Zoological Nomenclature (ZooBank) under LSID: urn:lsid:zoobank.org:pub:E0E65311-CB76-4AAE-9720-CDF1E6473E71. The new species name Hynobius notialis sp. nov. has been registered under LSID: urn:lsid:zoobank.org:act:82240BF1-7EFB-4DE3-A977-DCB8D4B8E841.
● Nomenclatural acts
The electronic edition of this article conforms to the requirements of the amended International Code of Zoological Nomenclature, and hence the new names contained herein are available under that code from the electronic edition of this article. This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the ICZN. The ZooBank LSIDs (Life Science Identifiers) can be resolved and the associated information viewed through any standard web browser by appending the LSID to the prefix “
http://zoobank.org/”. The LSID for this publication is: urn:lsid:zoobank.org:pub:E0E65311-CB76-4AAE-9720-CDF1E6473E71. The electronic edition of this work was published in a journal with an ISSN, and has been archived and is available from the following digital repositories: PubMed Central.
3.7.3. Hynobius geojeensis sp. nov. Min and Borzée, 2020
Synonymy:
“
Hynobius leechii (partim): [
72]: p. 33; [
73]: p. 20”.
“
Hynobius quelpaertensis (partim): [
52]: p. 1166”.
“
Hynobius leechii Clade HC3: [
53]: pp. 25–32; [
54]: p. 108; [
42]: pp. 475–503; [
55]: pp. 165–178”.
● Holotype
Voucher CGRB15863 (deposited in the Conservation Genome Resources Bank for Korean Wildlife (CGRB), College of Veterinary Medicine, Seoul National University; field ID mms3149); adult collected by Mi-Sook Min, HaeJun Baek and Dong Youn Kim on 6 April 2011 in Geoje-myeon, Geoje-si, Gyeongsang Nam-do, Republic of Korea (34.850992° N, 128.590398° E;
Figure 10). Measurements and counts in
Table 9.
● Paratypes
Vouchers CGRB15866, CGRB15877, CGRB15878 (deposited in the Conservation Genome Resources Bank for Korean Wildlife (CGRB), College of Veterinary Medicine, Seoul National University; field IDs mms3152, mms3904 and mms3909 respectively). CGRB15866 adult collected by Mi-Sook Min, HaeJun Baek and Dong Youn Kim on 6 April 2011 in Geoje-myeon, Geoje-si, Gyeongsang Nam-do, Republic of Korea (34.850992° N, 128.590398° E). CGRB15877 adult collected by Mi-Sook Min on 13 March 2012 in Mundong-dong, Geoje-si, Gyeongsang Nam-do, Republic of Korea (34.857903° N, 128.657295° E). CGRB15878 adult collected by Mi-Sook Min on 13 March 2012 in Mundong-dong, Geoje-si, Gyeongsang Nam-do, Republic of Korea (34.857903° N, 128.657295° E). Measurements and counts in
Table 9 illustrations in
Figure S3.
● Etymology
The species is endemic to Geoje island in the Republic of Korea, and the name H. geojeensis sp. nov. comes from the name of the island where it is restricted. The vernacular name of the species, Geoje salamander, reflects the scientific name of the special, as does its Korean name: 거제도롱뇽 (pronounced: Geoje Dorongnyong).
● Identity, diagnosis and distribution
The species is best identifiable based on range, although it is sympatric with
H. notialis on the northern half of its range, north of the city of Geoje (Graphical Abstract).
Hynobius geojeensis is generally broader and longer than
H. notialis (
Table 5 and
Table 9), with one exception: limbs are shorted than that of
H. notialis based on body size (
Figure S1) and body ratios (
Figure S2). The species can be identified in the area of sympatry if (in mm): SVL < 45.59, TL < 32.67, GA < 22.94, CW < 7.58, HLL < 7.97 and HL < 10.70 (
Table 5). The species breeds in natural and agricultural wetlands (
Figure 11).
● ZooBank registration
We hereby state that the present paper has been registered to the Official Register of Zoological Nomenclature (ZooBank) under LSID: urn:lsid:zoobank.org:pub:E0E65311-CB76-4AAE-9720-CDF1E6473E71. The new species name Hynobius geojeensis sp. nov. has been registered under LSID: urn:lsid:zoobank.org:act:CB0E13D1-4B81-4A59-9548-5A7B6B79856F.
● Nomenclatural acts
The electronic edition of this article conforms to the requirements of the amended International Code of Zoological Nomenclature, and hence the new names contained herein are available under that Code from the electronic edition of this article. This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the ICZN. The ZooBank LSIDs (Life Science Identifiers) can be resolved and the associated information viewed through any standard web browser by appending the LSID to the prefix “
http://zoobank.org/”. The LSID for this publication is: urn:lsid:zoobank.org:pub:E0E65311-CB76-4AAE-9720-CDF1E6473E71. The electronic edition of this work was published in a journal with an ISSN, and has been archived and is available from the following digital repositories: PubMed Central.
3.7.4. Hynobius perplicatus sp. nov. Min and Borzée, 2020
Synonymy:
“
Hynobius leechii (partim): [
72]: p. 33; [
73]: p. 20”.
“
Hynobius quelpaertensis (partim): [
52]: p. 1166”.
“
Hynobius leechii Clade HC4: [
53]: pp. 25–32; [
54]: p. 108; [
55]: pp. 165–178”.
● Holotype
Voucher CGRB15895 (deposited in the Conservation Genome Resources Bank for Korean Wildlife (CGRB), College of Veterinary Medicine, Seoul National University; field ID mms4082); adult collected by Mi-Sook Min, HaeJun Baek and DongYoun Kim on 22 March 2012 in Yongdeok-myeon, Uiryeong-gun, Gyeongsang Nam-do, Republic of Korea (35.348198° N, 128.291133° E;
Figure 12). Measurements and counts in
Table 9.
● Paratypes
Vouchers CGRB15893, CGRB15894, CGRB15896 (deposited in the Conservation Genome Resources Bank for Korean Wildlife (CGRB), College of Veterinary Medicine, Seoul National University; field IDs mms4072, mms4076 and mms4083 respectively). CGRB15893, adult collected by Mi-Sook Min, HaeJun Baek and DongYoun Kim on 22 March 2012 in Yongdeok-myeon, Uiryeong-gun, Gyeongsang Nam-do, Republic of Korea, Republic of Korea (35.348198° N, 128.291133° E). CGRB15894, adult collected by Mi-Sook Min, HaeJun Baek and DongYoun Kim on 22 March 2012in Yongdeok-myeon, Uiryeong-gun, Gyeongsang Nam-do, Republic of Korea, Republic of Korea (35.348198° N, 128.291133° E). CGRB15896, adult collected by Mi-Sook Min, HaeJun Baek and DongYoun Kim on 22 March 2012 in Yongdeok-myeon, Uiryeong-gun, Gyeongsang Nam-do, Republic of Korea, Republic of Korea (35.348198° N, 128.291133° E). Measurements and counts in
Table 9 illustrations in
Figure S3.
● Etymology
The species occurs on a limited area, sandwiched between the ranges of H. leechii and H. notialis. The current range is difficult to explain, and is likely related to glaciation patterns, which are so far unresolved, and thus rendering researchers perplexed about the intricate range of this cryptic species. The name H. perplicatus sp. nov. comes from this yet unresolved origin of the species’ range, the state of mind of researchers, and the cryptic morphology of the species. “Perplicatus” comes from Latin and means “interlaced, entangled, muddled, intricate, cryptic”. The vernacular name of the species, or Cryptic Uiryeong salamander, reflects the scientific name of the special, as does its Korean name: 숨은의령도롱뇽 (pronounced: Sumeun Uiryeong Dorongnyong).
● Identity, diagnosis and distribution
To date, the species is known to range from Daebyeong-myeon in Hapcheon on the north west, Sinan-myeon in Sancheong on the south west, Jijeong-myeon in Uiryeong on the south east and Cheongdeok-myeon in Hapcheon on the north east (Graphical Abstract). The species is most easily identified based on range, despite all known populations being sympatric with
H. notialis.
Hynobius perplicatus is the largest of all Korean
Hynobius species (
Table 5 and
Table 9). While there is no discrete known morphological feature for the species and some individuals may be overlapping with
H. notialis for some characteristics.
Hynobius perplicatus is noticeably larger in term or snout-vent-length, tail length, head length, head width and inter-orbital distance. When in sympatry, the species can be identified if (in mm): SVL > 68.21, TL > 62.59, HL > 15.81, HW > 12.78, HL > 4.39 (
Table 5). Some characteristics are however of higher value for
H. notialis and identification needs to include all variables to be reliable (
Table 5;
Figures S1 and S2). The species breeds in natural and agricultural wetlands (
Figure 13).
● ZooBank registration
We hereby state that the present paper has been registered to the Official Register of Zoological Nomenclature (ZooBank) under LSID: urn:lsid:zoobank.org:pub:E0E65311-CB76-4AAE-9720-CDF1E6473E71. The new species name Hynobius perplicatus sp. nov. has been registered under LSID: urn:lsid:zoobank.org:act:93D07C4B-1334-4370-8FDB-9BD0A2DD9645.
● Nomenclatural acts
The electronic edition of this article conforms to the requirements of the amended International Code of Zoological Nomenclature, and hence the new names contained herein are available under that Code from the electronic edition of this article. This published work and the nomenclatural acts it contains have been registered in ZooBank, the online registration system for the ICZN. The ZooBank LSIDs (Life Science Identifiers) can be resolved and the associated information viewed through any standard web browser by appending the LSID to the prefix “
http://zoobank.org/”. The LSID for this publication is: urn:lsid:zoobank.org:pub:E0E65311-CB76-4AAE-9720-CDF1E6473E71. The electronic edition of this work was published in a journal with an ISSN, and has been archived and is available from the following digital repositories: PubMed Central.
3.7.6. Conservation Status
The assessment presented here follows the IUCN red List criteria and categories (
www.iucnredlist.org/resources/redlistguidelines). All
Hynobius in Korea are facing severe threat, mostly due to habitat destruction and climate change. Most vertebrates have ecological preferences generally matching with those of early humans [
74], resulting in early human settlements, and subsequent urbanisation of the habitat such as large alluvial plains favoured by amphibians [
75,
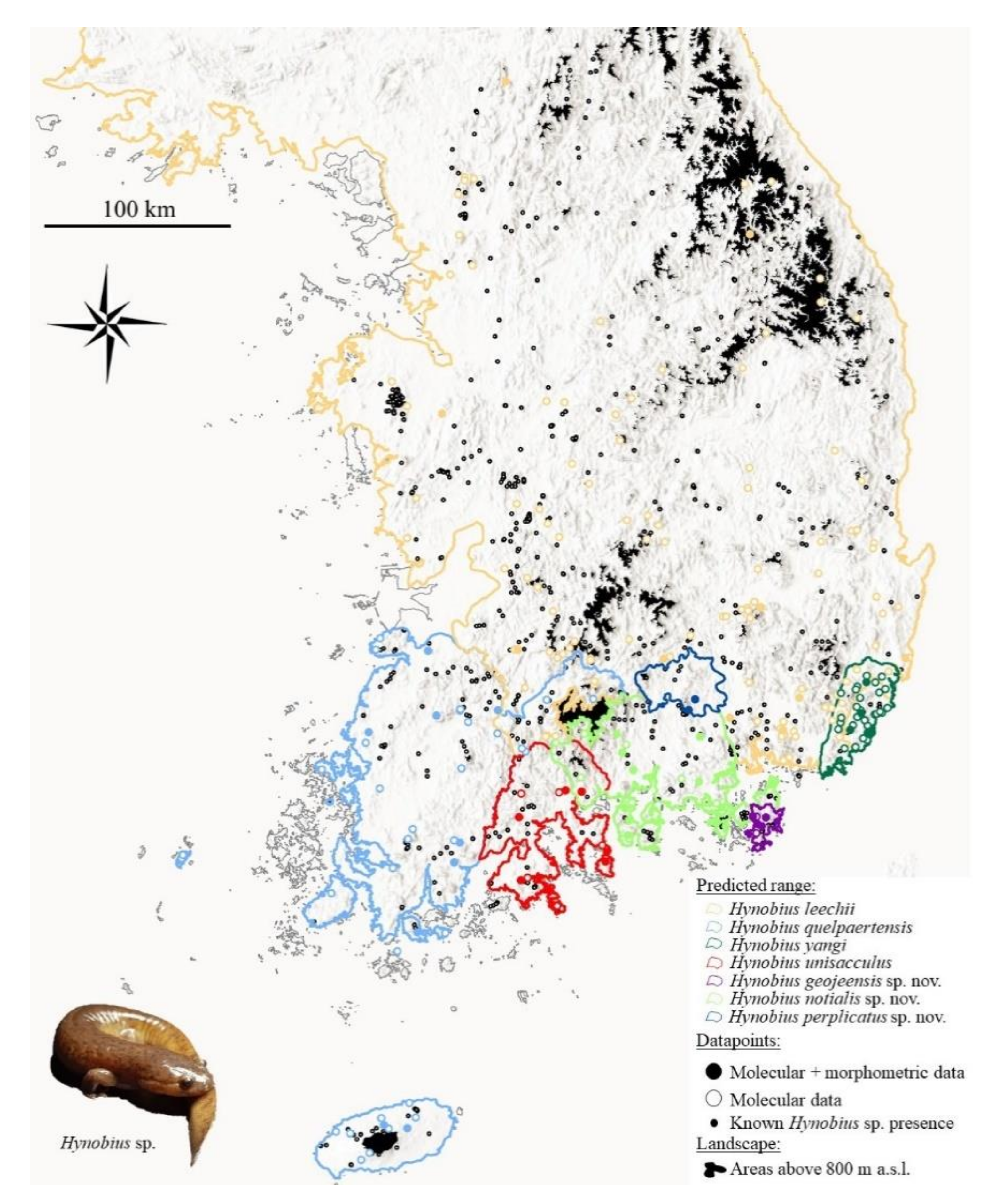
76], matching with zones of species abundance and pushing species towards secondary habitats. Namely, here the city of Gwangyang, Jinju, Goseong, Tongyeong and Haman (Graphical Abstract) have encroached on the range of the species. In addition, modification of the habitat for rice agriculture has a negative impact on the breeding ecology of the species [
77].
All Korean
Hynobius clades are also impacted by climate change in similar ways, with the exception of sea level rise and habitat salinisation not impacting
H. perplicatus. Despite calls to limit the current global warming to 1.5 °C above preindustrial levels by the Paris Climate Agreements [
78], environmental changes to the ecosystems are estimated to severely impact water resources [
79,
80]. In addition, the current environmental predictions based on the respect of the Paris Climate Agreements estimate a 2.3 °C increase in air temperature in Asia, and 2.7 °C for North East Asia (peaking at 7.0 °C under a 4 °C scenario if the Paris Climate Agreements are not held; [
81]). While critical maximal temperatures are not known for the focal species, experiments for other amphibians show a decrease in fitness through lower speed and resistance to chemical [
82]. In addition, the increase in temperature will make rice grows faster [
83] and farmers at low latitude in the Republic of Korea take advantage of this phenomenon to alternate barley and rice agriculture in the same fields. This results in an earlier rice plantation, with ploughing while larvae have not metamorphosed yet.
Climate change results in the rise of sea water level, and some sub-populations of the focal species are under the risk of being submerged by sea water [
84]. A sea level raise by 60 cm would result in the direct loss of habitat for all species at the exception of
H. perplicatus (50 to 70 sea water rise under RCP 4.5 scenario by 2100; [
85]). In addition, with sea level rise, the coastal habitat will become salinised and will not be adequate for the species [
86,
87,
88]. Moreover, climate change may be expected to result in a northern shift of species range to follow adequate ecological requirements [
89], however, and despite the unlikeliness of such dispersal to happen, the current range borders for all three species prevent such shift. Finally the species are under threat because of invasive species such as
Lithobates catesbeianus [
90] emerging diseases such as Batrachochytrids [
91,
92,
93], and the synergy of both type of invasive and pathogens being increasingly released in the environment [
94].
Hynobius yangi, H. unisacculus, H. geojeensis, H. notialis and H. perplicatus have very narrow ranges, <5000 km2, fragmented by expenses of saline water and urban areas. The populations have been observed, inferred and projected to be declining in extent of occurrence, area of occupancy, quality of habitat, number of subpopulations and number of mature individuals. As populations sizes are not known, precise dynamics have not been determined and quantitative analyses have not been conducted for any of the species, so the criteria A, C, D and E cannot be used for threat risk estimates.
The range of H. notialis is estimated to be <5000 km2, which is assigned this threshold to its extent of occurrence (EOO) due to the absence of data on landscape use and presence. The population is known to be present on the mainland and three disconnected islands, and the population is in continuing decline. Therefore, we estimate H. notialis to be in the Endangered category following the criteria of the IUCN Red List of Threatened Species under the criteria B1ab(i,ii,iii,iv,v).
The range of H. geojeensis is estimated to be <500 km2, which is assigned this threshold by the EOO for the species. In addition, the area of occupancy (AOO) of the species, based on island area as a proxy for range is <100 km2. The population is known to be present on the island only, and it is in continuing decline. Therefore, we estimate H. notialis to be in the Critically Endangered category following the criteria of the IUCN Red List of Threatened Species under the criteria B1ab(i,ii,iii,iv,v).
The range of H. perplicatus is estimated to be <1000 km2, which is assigned this threshold to the EOO due to the absence of data on landscape use and presence. The population is known to be present at two connected sites, but it is also likely to occur in linked valleys. The population is in continuing decline. Therefore, we estimate H. perplicatus to be in the Endangered category following the criteria of the IUCN Red List of Threatened Species under the criteria B1ab(i,ii,iii,iv,v).
The range of H. yangi is estimated to be <1000 km2, although the ancestral range may have reached 1200 km2, now reduced by about 150 km2 by the city of Busan and about 100 km2 by the city of Ulsan. Therefore, the species EOO is <1000 km2 and its AOO circa half of this value based on landscape use and the heavy urbanisation of the landscape within the range of the species. The population is in continuous decline, with several extirpated population due to development and the remaining sub-populations are divided into a maximum of four disconnected areas due to urban tracts and motorways. While threats are well understood, they have not ceased, and the range of the species is constantly decreasing due to expansion and urbanisation from the cities of Busan and Ulsan. Therefore, we estimate H. yangi to be in the Endangered category following the criteria of the IUCN Red List of Threatened Species under the criteria B1ab(i,ii,iii,iv,v).
The range of H. unisacculus is estimated to be <2500 km2, which is assigned this threshold by the EOO for the species. In addition, the AOO of the species is <2000 km2 based on landscape and habitat that can be used by the species. The population is severely fragmented due to the species distribution on islands, peninsula and the mainland, and urban areas and highways. In addition, a continuous decline is observed for EOO, AOO, and extant and quality of the habitat. Therefore, we estimate H. unisacculus to be in the Vulnerable category following the criteria of the IUCN Red List of Threatened Species under the criteria B1ab(i,ii,iii).
The range of H. quelpaertensis is estimated to be <20,000 km2, which is assigned this value to the species EOO, and an unknown AOO due to the absence of data on the fine scale distribution and habitat use by the species. In addition, the population is fragmentated due to the coastal and continental distribution of the species. The habitat is continuously decreasing in quality and extent, resulting in decreasing in EOO and AOO. The threats to H. quelpaertensis are known but not mitigated. Therefore, we estimate H. quelpaertensis to be in the Vulnerable category following the criteria of the IUCN Red List of Threatened Species under the criteria B1ab(i,ii,iii).
Finally, the range of H. leechii is estimated to be significantly wider than 20,000 km2, and the northern boundary of the species range is not yet defined. Consequently, the EOO of the species is larger than the threshold used to trigger any of the threatened categories following the criteria of the IUCN Red List of Threatened Species. Therefore, we estimate H. leechii to be falling in the category Least Concern following the criteria of the IUCN Red List of Threatened Species.

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}