Differences in the Loin Tenderness of Iberian Pigs Explained through Dissimilarities in Their Transcriptome Expression Profile

 , ,
, ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Material and Phenotypic Data
2.2. Transcriptomic Analyses
2.2.1. RNA Extraction, Library Preparation and Sequencing
2.2.2. Bioinformatics Analyses
Mapping, Assembly and Identification of Novel Isoforms
Differential Expression Analyses
Gene Functional Classification, Network and Pathway Analyses
2.3. RNA-seq Results Validation by Quantitative PCR
3. Results
3.1. Characterization of Longissimus Dorsi Transcriptome
3.2. Differential Expression Analyses
3.3. Gene Functional Analysis
3.3.1. Canonical Pathways Analysis
3.3.2. Transcription Regulatory Factors
3.4. RNA-Seq Validation by qPCR
4. Discussion
4.1. Proteolysis Process
4.2. Skeletal Muscle Tissue Development and Growth
4.3. Lipid Metabolism
4.4. Collagen Metabolic Process
4.5. Transcription Regulatory Factors
4.6. Candidate Genes for an Iberian Pig-Breeding Program
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Maltin, C.; Balcerzak, D.; Tilley, R.; Delday, M. Determinants of Meat Quality: Tenderness. Proc. Nutr. Soc. 2003, 62, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; Choi, B.D.; Kim, B.C.; Park, S.S.; Hong, K.C. Associations of the Variation in the Porcine Myogenin Gene with Muscle Fibre Characteristics, Lean Meat Production and Meat Quality Traits. J. Anim. Breed. Genet. 2009, 126, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Čandek-Potokar, M.; Žlender, B.; Lefaucheur, L.; Bonneau, M. Effects of Age And/or Weight at Slaughter on Longissimus Dorsi Muscle: Biochemical Traits and Sensory Quality in Pigs. Meat Sci. 1998, 48, 287–300. [Google Scholar] [CrossRef]
- Lonergan, S.M.; Huff-Lonergan, E.; Rowe, L.J.; Kuhlers, D.L.; Jungst, S.B. Selection for Lean Growth Efficiency in Duroc Pigs Influences Pork Quality. J. Anim. Sci. 2001, 79, 2075–2085. [Google Scholar] [CrossRef]
- Rosenvold, K.; Andersen, H.J. Factors of Significance for Pork Quality—A Review. Meat Sci. 2003, 64, 219–237. [Google Scholar] [CrossRef]
- Suzuki, K.; Irie, M.; Kadowaki, H.; Shibata, T.; Kumagai, M.; Nishida, A. Genetic Parameter Estimates of Meat Quality Traits in Duroc Pigs Selected for Average Daily Gain, Longissimus Muscle Area, Backfat Thickness, and Intramuscular Fat Content. J. Anim. Sci. 2005, 83, 2058–2065. [Google Scholar] [CrossRef]
- Miar, Y.; Plastow, G.S.; Moore, S.S.; Manafiazar, G.; Charagu, P.; Kemp, R.A.; van Haandel, B.; Huisman, A.E.; Zhang, C.Y.; Mckay, R.M.; et al. Genetic and Phenotypic Parameters for Carcass and Meat Quality Traits in Commercial Crossbred Pigs. J. Anim. Sci. 2014, 92, 2869–2884. [Google Scholar] [CrossRef]
- Fernández-Barroso, M.Á.; Silió, L.; Rodríguez, C.; Palma-Granados, P.; López, A.; Caraballo, C.; García-Casco, J.M.; Sánchez-Esquiliche, F.; Gómez-Carballar, F.; Muñoz, M. Genetic Parameter Estimation and Gene Association Analyses for Meat Quality Traits in Open-Air Free-Range Iberian Pigs. J. Anim. Breed. Genet. 2020, 1–18. [Google Scholar] [CrossRef]
- Lindholm-Perry, A.K.; Rohrer, G.A.; Holl, J.W.; Shackelford, S.D.; Wheeler, T.L.; Koohmaraie, M.; Nonneman, D. Relationships among Calpastatin Single Nucleotide Polymorphisms, Calpastatin Expression and Tenderness in Pork Longissimus. Anim. Genet. 2009, 40, 713–721. [Google Scholar] [CrossRef]
- Gandolfi, G.; Pomponio, L.; Ertbjerg, P.; Karlsson, A.H.; Nanni Costa, L.; Lametsch, R.; Russo, V.; Davoli, R. Investigation on CAST, CAPN1 and CAPN3 Porcine Gene Polymorphisms and Expression in Relation to Post-Mortem Calpain Activity in Muscle and Meat Quality. Meat Sci. 2011, 88, 694–700. [Google Scholar] [CrossRef]
- Honikel, K.O. Reference Methods for the Assessment of Physical Characteristics of Meat. Meat Sci. 1998, 49, 447–457. [Google Scholar] [CrossRef]
- Tejerina, D.; García-Torres, S.; Cava, R. Water-Holding Capacity and Instrumental Texture Properties of M. Longissimus Dorsi and M. Serratus Ventralis from Iberian Pigs as Affected by the Production System. Livest. Sci. 2012, 148, 46–51. [Google Scholar] [CrossRef]
- Damon, M.; Wyszynska-Koko, J.; Vincent, A.; Hérault, F.; Lebret, B. Comparison of Muscle Transcriptome between Pigs with Divergent Meat Quality Phenotypes Identifies Genes Related to Muscle Metabolism and Structure. PLoS ONE 2012, 7, e33763. [Google Scholar] [CrossRef] [PubMed]
- Hamill, R.M.; McBryan, J.; McGee, C.; Mullen, A.M.; Sweeney, T.; Talbot, A.; Cairns, M.T.; Davey, G.C. Functional Analysis of Muscle Gene Expression Profiles Associated with Tenderness and Intramuscular Fat Content in Pork. Meat Sci. 2012, 92, 440–450. [Google Scholar] [CrossRef] [PubMed]
- Horodyska, J.; Wimmers, K.; Reyer, H.; Trakooljul, N.; Mullen, A.M.; Lawlor, P.G.; Hamill, R.M. RNA-Seq of Muscle from Pigs Divergent in Feed Efficiency and Product Quality Identifies Differences in Immune Response, Growth, and Macronutrient and Connective Tissue Metabolism. BMC Genom. 2018, 19, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Ferre, M.G.; Aguilera, J.F.; Nieto, R. Muscle Fractional Protein Synthesis Is Higher in Iberian than in Landrace Growing Pigs Fed Adequate or Lysine-Deficient Diets. J. Nutr. 2005, 135, 469–478. [Google Scholar] [CrossRef]
- Lopez-Bote, C.J. Sustained Utilization of the Iberian Pig Breed. Meat Sci. 1998, 49. [Google Scholar] [CrossRef]
- Alves, E.; Óvilo, C.; Rodríguez, M.C.; Silió, L. Mitochondrial DNA Sequence Variation and Phylogenetic Relationships among Iberian Pigs and Other Domestic and Wild Pig Populations. Anim. Genet. 2003, 34, 319–324. [Google Scholar] [CrossRef]
- Fabuel, E.; Barragán, C.; Silió, L.; Rodríguez, M.C.; Toro, M.A. Analysis of Genetic Diversity and Conservation Priorities in Iberian Pigs Based on Microsatellite Markers. Heredity 2004, 93, 104–113. [Google Scholar] [CrossRef]
- Ollivier, L. European Pig Genetic Diversity: A. Minireview. Animal 2009, 3, 915–924. [Google Scholar] [CrossRef]
- Muñoz, M.; Sánchez-Esquiliche, F.; Caraballo, C.; Gómez, F.; Pariente, J.M.; Silió, L.; Rodríguez, C.; García-Casco, J.M. Animal Breeding Scheme Applied to the Quality of Pure Iberian Montanera Pigs. Arch. Zootec. 2018, 67, 9–11. [Google Scholar] [CrossRef]
- Combes, S.; Lepetit, J.; Darche, B.; Lebas, F. Effect of Cooking Temperature and Cooking Time on Warner-Bratzler Tenderness Measurement and Collagen Content in Rabbit Meat. Meat Sci. 2004, 66, 91–96. [Google Scholar] [CrossRef]
- Legarra, A.; Varona, L.; de Maturana, E.L. TM Threshold Model; INRA-SAGA: Toulouse, France, 2008. [Google Scholar]
- Muñoz, M.; García-Casco, J.M.; Caraballo, C.; Fernández-Barroso, M.Á.; Sánchez-Esquiliche, F.; Gómez, F.; Rodríguez, M.d.C.; Silió, L. Identification of Candidate Genes and Regulatory Factors Underlying Intramuscular Fat Content Through Longissimus Dorsi Transcriptome Analyses in Heavy Iberian Pigs. Front. Genet. 2018, 9, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering Splice Junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential Gene and Transcript Expression Analysis of RNA-Seq Experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef]
- Storey, J.D.; Tibshirani, R. Statistical Significance for Genomewide Studies. Proc. Natl. Acad. Sci. USA 2003, 100, 9440–9445. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING Database in 2017: Quality-Controlled Protein-Protein Association Networks, Made Broadly Accessible. Nucleic Acids Res. 2017, 45, D362–D368. [Google Scholar] [CrossRef]
- Krämer, A.; Green, J.; Pollard, J.; Tugendreich, S. Causal Analysis Approaches in Ingenuity Pathway Analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate Normalization of Real-Time Quantitative RT-PCR Data by Geometric Averaging of Multiple Internal Control Genes. Genome Biol. 2002, 3, research0034-1. [Google Scholar] [CrossRef]
- Miron, M.; Woody, O.Z.; Marcil, A.; Murie, C.; Sladek, R.; Nadon, R. A Methodology for Global Validation of Microarray Experiments. BMC Bioinform. 2006, 7. [Google Scholar] [CrossRef]
- Zappaterra, M.; Gioiosa, S.; Chillemi, G.; Zambonelli, P.; Davoli, R. Muscle Transcriptome Analysis Identifies Genes Involved in Ciliogenesis and the Molecular Cascade Associated with Intramuscular Fat Content in Large White Heavy Pigs. PLoS ONE 2020, 15, e0233372. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.; Pimentel, H.; Trapnell, C.; Pachter, L. Identification of Novel Transcripts in Annotated Genomes Using RNA-Seq. Bioinformatics 2011, 27, 2325–2329. [Google Scholar] [CrossRef] [PubMed]
- Kemp, C.M.; Sensky, P.L.; Bardsley, R.G.; Buttery, P.J.; Parr, T. Tenderness—An Enzymatic View. Meat Sci. 2010, 84, 248–256. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, D.; Taylor. Post-Mortem Muscle Proteolysis and Meat Tenderness. In Muscle Development of Livestock Animals: Physiology, Genetics, and Meat Quality; Centre for Agriculture and Bioscience International: Wallingford, UK, 2004; pp. 363–388. [Google Scholar]
- Li, S.; Xu, X.; Zhou, G. The Roles of the Actin-Myosin Interaction and Proteolysis in Tenderization during the Aging of Chicken Muscle. Poult. Sci. 2012, 91, 150–160. [Google Scholar] [CrossRef] [PubMed]
- D’Andrea, M.; Dal Monego, S.; Pallavicini, A.; Modonut, M.; Dreos, R.; Stefanon, B.; Pilla, F. Muscle Transcriptome Profiling in Divergent Phenotype Swine Breeds during Growth Using Microarray and RT-PCR Tools. Anim. Genet. 2011, 42, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Piórkowska, K.; Żukowski, K.; Ropka-Molik, K.; Tyra, M. Detection of Genetic Variants between Different Polish Landrace and Puławska Pigs by Means of RNA-Seq Analysis. Anim. Genet. 2018, 49, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Casiró, S.; Velez-Irizarry, D.; Ernst, C.W.; Raney, N.E.; Bates, R.O.; Charles, M.G.; Steibel, J.P. Genome-Wide Association Study in an F2 Duroc X Pietrain Resource Population for Economically Important Meat Quality and Carcass Traits. J. Anim. Sci. 2017, 95, 545–558. [Google Scholar] [CrossRef]
- Gad, A.K.B.; Nehru, V.; Ruusala, A.; Aspenström, P. RhoD Regulates Cytoskeletal Dynamics via the Actin Nucleation-Promoting Factor WASp Homologue Associated with Actin Golgi Membranes and Microtubules. Mol. Biol. Cell 2012, 23, 4807–4819. [Google Scholar] [CrossRef]
- Toydemir, R.M.; Chen, H.; Proud, V.K.; Martin, R.; Van Bokhoven, H.; Hamel, B.C.J.; Tuerlings, J.H.; Stratakis, C.A.; Jorde, L.B.; Bamshad, M.J. Trismus-Pseudocamptodactyly Syndrome Is Caused by Recurrent Mutation of MYH8. Am. J. Med. Genet. Part A 2006, 140, 2387–2393. [Google Scholar] [CrossRef]
- Karsch-Mizrachi, I.; Feghali, R.; Shows, T.B.; Leinwand, L.A. Generation of a Full-Length Human Perinatal Myosin Heavy-Chain-Encoding cDNA. Gene 1990, 89, 289–294. [Google Scholar] [CrossRef]
- Liu, S.; Han, W.; Jiang, S.; Zhao, C.; Wu, C. Integrative Transcriptomics and Proteomics Analysis of Longissimus Dorsi Muscles of Canadian Double-Muscled Large White Pigs. Gene 2016, 577, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Sparrow, J.C.; Nowak, K.J.; Durling, H.J.; Beggs, A.H.; Wallgren-Pettersson, C.; Romero, N.; Nonaka, I.; Laing, N.G. Muscle Disease Caused by Mutations in the Skeletal Muscle Alpha-Actin Gene (ACTA1). Neuromusc. Disord. 2003, 519–531. [Google Scholar] [CrossRef]
- Nowak, K.J.; Wattanasirichaigoon, D.; Goebel, H.H.; Wilce, M.; Pelin, K.; Donner, K.; Jacob, R.L.; Hübner, C.; Oexle, K.; Anderson, J.R.; et al. Mutations in the Skeletal Muscle Alpha-Actin Gene in Patients with Actin Myopathy and Nemaline Myopathy. Nat. Genet. 1999, 23, 208–212. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Xu, X.; Li, M.; Ming, H.; Jiang, J. Integrative Analysis of Transcriptomics and Proteomics of Skeletal Muscles of the Chinese Indigenous Shaziling Pig Compared with the Yorkshire Breed. BMC Genet. 2016, 17, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ponsuksili, S.; Siengdee, P.; Du, Y.; Trakooljul, N.; Murani, E.; Schwerin, M.; Wimmers, K. Identification of Common Regulators of Genes in Co-Expression Networks Affecting Muscle and Meat Properties. PLoS ONE 2015, 10, e0123678. [Google Scholar] [CrossRef]
- Jerez-Timaure, N.; Gallo, C.; Ramírez-Reveco, A.; Greif, G.; Strobel, P.; Pedro, A.V.F.; Morera, F.J. Early Differential Gene Expression in Beef Longissimus Thoracis Muscles from Carcasses with Normal (<5.8) and High (>5.9) Ultimate pH. Meat Sci. 2019, 153, 117–125. [Google Scholar] [CrossRef]
- Sentandreu, M.A.; Coulis, G.; Ouali, A. Role of Muscle Endopeptidases and Their Inhibitors in Meat Tenderness. Trends Food Sci. Technol. 2002, 13, 400–421. [Google Scholar] [CrossRef]
- Goll, D.E.; Thompson, V.F.; Li, H.; Wei, W.; Cong, J. The Calpain System. Physiol. Rev. 2003, 1990, 731–801. [Google Scholar] [CrossRef]
- Koohmaraie, M.; Geesink, G.H. Contribution of Postmortem Muscle Biochemistry to the Delivery of Consistent Meat Quality with Particular Focus on the Calpain System. Meat Sci. 2006, 74, 34–43. [Google Scholar] [CrossRef]
- Van Laack, R.L.J.M.; Stevens, S.G.; Stalder, K.J. The Influence of Ultimate pH and Intramuscular Fat Content on Pork Tenderness and Tenderization. J. Anim. Sci. 2001, 79, 392–397. [Google Scholar] [CrossRef]
- Wu, C.; Dedhar, S. Integrin-Linked Kinase (ILK) and Its Interactors: A New Paradigm for the Coupling of Extracellular Matrix to Actin Cytoskeleton and Signaling Complexes. J. Cell Biol. 2001, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Giancotti, F.G.; Ruoslahti, E. Integrin Signaling. Science 1999, 1028–1032. [Google Scholar] [CrossRef] [PubMed]
- Ouali, A.; Herrera-Mendez, C.H.; Coulis, G.; Becila, S.; Boudjellal, A.; Aubry, L.; Sentandreu, M.A. Revisiting the Conversion of Muscle into Meat and the Underlying Mechanisms. Meat Sci. 2006, 74, 44–58. [Google Scholar] [CrossRef] [PubMed]
- Torrado, M.; Nespereira, B.; López, E.; Centeno, A.; Castro-Beiras, A.; Mikhailov, A.T. ANKRD1 Specifically Binds CASQ2 in Heart Extracts and Both Proteins Are Co-Enriched in Piglet Cardiac Purkinje Cells. J. Mol. Cell. Cardiol. 2005, 38, 353–365. [Google Scholar] [CrossRef] [PubMed]
- Ovilo, C.; Benítez, R.; Fernández, A.; Núñez, Y.; Ayuso, M.; Fernández, A.I.; Rodríguez, C.; Isabel, B.; Rey, A.I.; López-Bote, C.; et al. Longissimus Dorsi Transcriptome Analysis of Purebred and Crossbred Iberian Pigs Differing in Muscle Characteristics. BMC Genom. 2014, 15, 413. [Google Scholar] [CrossRef]
- Li, X.J.; Zhou, J.; Liu, L.Q.; Qian, K.; Wang, C.L. Identification of Genes in Longissimus Dorsi Muscle Differentially Expressed between Wannanhua and Yorkshire Pigs Using RNA-Sequencing. Anim. Genet. 2016, 47, 324–333. [Google Scholar] [CrossRef]
- Ponsuksili, S.; Murani, E.; Phatsara, C.; Schwerin, M.; Schellander, K.; Wimmers, K. Porcine Muscle Sensory Attributes Associate with Major Changes in Gene Networks Involving CAPZB, ANKRD1, and CTBP2. Funct. Integr. Genom. 2009, 9, 455–471. [Google Scholar] [CrossRef]
- Velez-Irizarry, D.; Casiro, S.; Daza, K.R.; Bates, R.O.; Raney, N.E.; Steibel, J.P.; Ernst, C.W. Genetic Control of Longissimus Dorsi Muscle Gene Expression Variation and Joint Analysis with Phenotypic Quantitative Trait Loci in Pigs 06 Biological Sciences 0604 Genetics. BMC Genom. 2019, 20, 1–19. [Google Scholar] [CrossRef]
- Zhu, J.; Shi, X.E.; Lu, H.; Xia, B.; Li, Y.; Li, X.; Zhang, Q.; Yang, G. RNA-Seq Transcriptome Analysis of Extensor Digitorum Longus and Soleus Muscles in Large White Pigs. Mol. Genet. Genom. 2016, 291, 687–701. [Google Scholar] [CrossRef]
- Kambadur, R.; Sharma, M.; Smith, T.P.L.; Bass, J.J. Mutations in Myostatin (GDF8) in Double-Muscled Belgian Blue and Piedmontese Cattle. Genome Res. 1997, 7, 910–916. [Google Scholar] [CrossRef]
- Clop, A.; Marcq, F.; Takeda, H.; Pirottin, D.; Tordoir, X.; Bibé, B.; Bouix, J.; Caiment, F.; Elsen, J.M.; Eychenne, F.; et al. A Mutation Creating a Potential Illegitimate microRNA Target Site in the Myostatin Gene Affects Muscularity in Sheep. Nat. Genet. 2006, 38, 813–818. [Google Scholar] [CrossRef] [PubMed]
- Qian, L.; Tang, M.; Yang, J.; Wang, Q.; Cai, C.; Jiang, S.; Li, H.; Jiang, K.; Gao, P.; Ma, D.; et al. Targeted Mutations in Myostatin by Zinc-Finger Nucleases Result in Double-Muscled Phenotype in Meishan Pigs. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Deng, B.; Zhang, F.; Wen, J.; Ye, S.; Wang, L.; Yang, Y.; Gong, P.; Jiang, S. The Function of Myostatin in the Regulation of Fat Mass in Mammals. Nutr. Metab. 2017. [Google Scholar] [CrossRef]
- Haenggi, T.; Fritschy, J.M. Role of Dystrophin and Utrophin for Assembly and Function of the Dystrophin Glycoprotein Complex in Non-Muscle Tissue. Cell. Mol. Life Sci. 2006, 1614–1631. [Google Scholar] [CrossRef] [PubMed]
- Fröhlich, T.; Kemter, E.; Flenkenthaler, F.; Klymiuk, N.; Otte, K.A.; Blutke, A.; Krause, S.; Walter, M.C.; Wanke, R.; Wolf, E.; et al. Progressive Muscle Proteome Changes in a Clinically Relevant Pig Model of Duchenne Muscular Dystrophy. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef]
- Ferrero, G.O.; Velazquez, F.N.; Caputto, B.L. The Kinase c-Src and the Phosphatase TC45 Coordinately Regulate c-Fos Tyrosine Phosphorylation and c-Fos Phospholipid Synthesis Activation Capacity. Oncogene 2012, 31, 3381–3391. [Google Scholar] [CrossRef]
- Reiner, G.; Heinricy, L.; Müller, E.; Geldermann, H.; Dzapo, V. Indications of Associations of the Porcine FOS Proto-Oncogene with Skeletal Muscle Fibre Traits. Anim. Genet. 2002, 33, 49–55. [Google Scholar] [CrossRef]
- Rajan, S.; Dang, H.C.P.; Djambazian, H.; Zuzan, H.; Fedyshyn, Y.; Ketela, T.; Moffat, J.; Hudson, T.J.; Sladek, R. Analysis of Early C2C12 Myogenesis Identifies Stably and Differentially Expressed Transcriptional Regulators Whose Knock-down Inhibits Myoblast Differentiation. Physiol. Genom. 2012, 44, 183–197. [Google Scholar] [CrossRef]
- Ayuso, M.; Fernández, A.; Núñez, Y.; Benitez, R.; Isabel, B.; Barragán, C.; Fernández, A.I.; Rey, A.I.; Medrano, J.F.; Cánovas, Á.; et al. Comparative Analysis of Muscle Transcriptome between Pig Genotypes Identifies Genes and Regulatory Mechanisms Associated to Growth, Fatness and Metabolism. PLoS ONE 2015, 10, e0145162. [Google Scholar] [CrossRef]
- García-Contreras, C.; Madsen, O.; Groenen, M.A.M.; López-García, A.; Vázquez-Gómez, M.; Astiz, S.; Núñez, Y.; Benítez, R.; Fernández, A.; Isabel, B.; et al. Impact of Genotype, Body Weight and Sex on the Prenatal Muscle Transcriptome of Iberian Pigs. PLoS ONE 2020, 15, e0227861. [Google Scholar] [CrossRef]
- Migita, T.; Narita, T.; Asaka, R.; Miyagi, E.; Nagano, H.; Nomura, K.; Matsuura, M.; Satoh, Y.; Okumura, S.; Nakagawa, K.; et al. Role of Insulin-like Growth Factor Binding Protein 2 in Lung Adenocarcinoma: IGF-Independent Antiapoptotic Effect via Caspase-3. Am. J. Pathol. 2010, 176, 1756–1766. [Google Scholar] [CrossRef] [PubMed]
- Hakuno, F.; Takahashi, S.-I. IGF1 Receptor Signaling Pathways. J. Mol. Endocrinol. 2018, 61, T69–T86. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.D. Consequences for Meat Quality of Reducing Carcass Fatness. In Reducing Fat in Meat Animals; Elsevier: Amsterdam, The Netherlands, 1990; pp. 344–397. [Google Scholar]
- Lee, J.H.; Song, K.D.; Lee, H.K.; Cho, K.H.; Park, H.C.; Park, K.D. Genetic Parameters of Reproductive and Meat Quality Traits in Korean Berkshire Pigs. Asian-Australas. J. Anim. Sci. 2015, 28, 1388–1393. [Google Scholar] [CrossRef]
- Sellier, P. Genetics of Meat and Carcass Traits. Genetics of the Pig; Centre for Agriculture and Bioscience International: Wallingford, UK, 1998; pp. 463–510. [Google Scholar]
- Laville, E.; Sayd, T.; Terlouw, C.; Chambon, C.; Damon, M.; Larzul, C.; Leroy, P.; Glénisson, J.; Chérel, P. Comparison of Sarcoplasmic Proteomes between Two Groups of Pig Muscles Selected for Shear Force of Cooked Meat. J. Agric. Food Chem. 2007, 55, 5834–5841. [Google Scholar] [CrossRef] [PubMed]
- Óvilo, C.; Oliver, A.; Noguera, J.L.; Clop, A.; Barragán, C.; Varona, L.; Rodríguez, C.; Toro, M.; Sánchez, A.; Pérez-Enciso, M.; et al. Test for Positional Candidate Genes for Body Composition on Pig Chromosome 6. Genet. Sel. Evol. 2002, 34, 465–479. [Google Scholar] [CrossRef]
- Li, X.; Kim, S.W.; Choi, J.S.; Lee, Y.M.; Lee, C.K.; Choi, B.H.; Kim, T.H.; Choi, Y.I.; Kim, J.J.; Kim, K.S. Investigation of Porcine FABP3 and LEPR Gene Polymorphisms and mRNA Expression for Variation in Intramuscular Fat Content. Mol. Biol. Rep. 2010, 37, 3931–3939. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.N.; Jiang, Y.Z.; Cen, W.M.; Xing, S.H.; Zhu, L.; Tang, G.Q.; Li, M.Z.; Jiang, A.A.; Lou, P.E.; Wen, A.X.; et al. Distribution of H-FABP and ACSL4 Gene Polymorphisms and Their Associations with Intramuscular Fat Content and Backfat Thickness in Different Pig Populations. Genet. Mol. Res. 2014, 13, 6759–6772. [Google Scholar] [CrossRef]
- Handschin, C.; Spiegelman, B.M. Peroxisome Proliferator-Activated Receptor γ Coactivator 1 Coactivators, Energy Homeostasis, and Metabolism. Endocr. Rev. 2006, 27, 728–735. [Google Scholar] [CrossRef]
- Lin, J.; Wu, H.; Tarr, P.T.; Zhang, C.Y.; Wu, Z.; Boss, O.; Michael, L.F.; Puigserver, P.; Isotani, E.; Olson, E.N.; et al. Transcriptional Co-Activator PGC-1α Drives the Formation of Slow-Twitch Muscle Fibres. Nature 2002, 418, 797–801. [Google Scholar] [CrossRef]
- Erkens, T.; De Smet, S.; Van Den Maagdenberg, K.; Stinckens, A.; Buys, N.; Van Zeveren, A.; Peelman, L.J. Association Analysis of PPARGC1A Mutations with Meat Quality Parameters in a Commercial Hybrid Pig Population. Czech J. Anim. Sci. 2010, 55, 200–208. [Google Scholar] [CrossRef]
- Chambon, P. The Nuclear Receptor Superfamily: A Personal Retrospect on the First Two Decades. Mol. Endocrinol. 2005, 1418–1428. [Google Scholar] [CrossRef] [PubMed]
- Hocquette, J.F.; Gondret, F.; Baza, E.; Mdale, F.; Jurie, C.; Pethick, D.W. Intramuscular Fat Content in Meat-Producing Animals: Development, Genetic and Nutritional Control, and Identification of Putative Markers. Animal 2010, 4, 303–319. [Google Scholar] [CrossRef] [PubMed]
- Weston, A.R.; Rogers, R.W.; Althen, T.G. REVIEW: The Role of Collagen in Meat Tenderness. Prof. Anim. Sci. 2002, 18, 107–111. [Google Scholar] [CrossRef]
- Purslow, P.P. Intramuscular Connective Tissue and Its Role in Meat Quality. Meat Sci. 2005, 70, 435–447. [Google Scholar] [CrossRef] [PubMed]
- Torrescano, G.; Sánchez-Escalante, A.; Giménez, B.; Roncalés, P.; Beltrán, J.A. Shear Values of Raw Samples of 14 Bovine Muscles and Their Relation to Muscle Collagen Characteristics. Meat Sci. 2003, 64, 85–91. [Google Scholar] [CrossRef]
- Ngapo, T.M.; Berge, P.; Culioli, J.; Dransfield, E.; De Smet, S.; Claeys, E. Perimysial Collagen Crosslinking and Meat Tenderness in Belgian Blue Double-Muscled Cattle. Meat Sci. 2002, 61, 91–102. [Google Scholar] [CrossRef]
- Dransfield, E.; Martin, J.F.; Bauchart, D.; Abouelkaram, S.; Lepetit, J.; Culioli, J.; Jurie, C.; Picard, B. Meat Quality and Composition of Three Muscles from French Cull Cows and Young Bulls. Anim. Sci. 2003, 76, 387–399. [Google Scholar] [CrossRef]
- Lepetit, J. A Theoretical Approach of the Relationships between Collagen Content, Collagen Cross-Links and Meat Tenderness. Meat Sci. 2007, 76, 147–159. [Google Scholar] [CrossRef]
- Schuppan, D.; Cantaluppi, M.C.; Becker, J.; Veit, A.; Bunte, T.; Troyer, D.; Schuppan, F.; Schmid, M.; Ackermann, R.; Hahn, E.G. Undulin, an Extracellular Matrix Glycoprotein Associated with Collagen Fibrils. J. Biol. Chem. 1990, 265, 8823–8832. [Google Scholar]
- Xu, J.; Wang, C.; Jin, E.; Gu, Y.; Li, S.; Li, Q. Identification of Differentially Expressed Genes in Longissimus Dorsi Muscle between Wei and Yorkshire Pigs Using RNA Sequencing. Genes Genom. 2018, 40, 413–421. [Google Scholar] [CrossRef]
- Hishida, T.; Nishizuka, M.; Osada, S.; Imagawa, M. The Role of C/EBPδ in the Early Stages of Adipogenesis. Biochimie 2009, 91, 654–657. [Google Scholar] [CrossRef] [PubMed]
- Pena, R.N.; Noguera, J.L.; García-Santana, M.J.; González, E.; Tejeda, J.F.; Ros-Freixedes, R.; Ibáñez-Escriche, N. Five Genomic Regions Have a Major Impact on Fat Composition in Iberian Pigs. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Figeac, N.; Mohamed, A.D.; Sun, C.; Schönfelder, M.; Matallanas, D.; Garcia-Munoz, A.; Missiaglia, E.; Collie-Duguid, E.; De Mello, V.; Pobbati, A.V.; et al. VGLL3 Operates via TEAD1, TEAD3 and TEAD4 to Influence Myogenesis in Skeletal Muscle. J. Cell Sci. 2019, 132, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, A.W.; Veerkamp, J.H. New Insights into the Structure and Function of Fatty Acid-Binding Proteins. Cell. Mol. Life Sci. 2002, 59, 1096–1116. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, M.P.; Iannuccelli, N.; Basso, B.; Bidanel, J.P.; Billon, Y.; Gandemer, G.; Gilbert, H.; Larzul, C.; Legault, C.; Riquet, J.; et al. Identification of QTL with Effects on Intramuscular Fat Content Fatty Acid Composition in a Duroc × Large White Cross. BMC Genet. 2007, 8. [Google Scholar] [CrossRef] [PubMed]
- Ayuso, M.; Fernández, A.; Núñez, Y.; Benítez, R.; Isabel, B.; Fernández, A.I.; Rey, A.I.; González-Bulnes, A.; Medrano, J.F.; Cánovas, Á.; et al. Developmental Stage, Muscle and Genetic Type Modify Muscle Transcriptome in Pigs: Effects on Gene Expression and Regulatory Factors Involved in Growth and Metabolism. PLoS ONE 2016, 11, e0167858. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene | Fold Change | Tender | Tough | q-Value |
|---|---|---|---|---|
| Guanylate binding protein 1 (GBP1) | 0.089 | 46.959 | 4.175 | 0.002 |
| Cholinergic Receptor Nicotinic Alpha 9 Subunit (CHRNA9) | 0.360 | 1.666 | 0.600 | 0.002 |
| Ras Homolog Family Member D (RHOD) | 0.438 | 1.414 | 0.620 | 0.025 |
| Calsequestrin 2 (CASQ2) | 0.447 | 15.284 | 6.832 | 0.002 |
| Ankyrin Repeat Domain 1 protein (ANKRD1) | 0.476 | 553.625 | 263.267 | 0.002 |
| Peroxisome Proliferator-Activated Receptor Gamma Coactivator 1-Alpha (PPARGC1A) | 0.508 | 13.825 | 7.024 | 0.015 |
| Cathepsine C (CTSC) | 0.567 | 63.679 | 36.137 | 0.016 |
| Fatty Acid Binding Protein 3 (FABP3) | 0.633 | 352.059 | 222.898 | 0.038 |
| Fos proto-oncogene (FOS) | 1.670 | 24.722 | 41.282 | 0.037 |
| Dystrophin (DMD) | 1.779 | 0.614 | 1.091 | 0.038 |
| Collagen Type XIV Alpha 1 Chain (COL14A1) | 2.017 | 3.035 | 6.122 | 0.057 |
| Myostatin (MSTN) | 2.038 | 2.977 | 6.067 | 0.009 |
| Growth Arrest and DNA Damage Inducible Beta protein (GADD45B) | 2.871 | 12.954 | 37.188 | 0.002 |
| Actin Alpha 1, Skeletal Muscle (ACTC1) | 4.085 | 9.750 | 39.830 | 0.002 |
| Myosin Heavy Chain 8 (MYH8) | 7.541 | 1269.750 | 9574.930 | 0.002 |
| Family With Sequence Similarity 180 Member B (FAM180B) | 8.830 | 1.959 | 17.297 | 0.008 |
| Term | Genes | Adjusted p-Value |
|---|---|---|
| GOBP | ||
| Skeletal muscle tissue development (GO:0007519) | MYLK2, MSTN, FOS, HLF, CXCL10, IGFBP5, ANKRD1, DMD, CXCL9, FOXN2 | 1.02 × 10−9 |
| Muscle cell development (GO:0055001) | CXCL10, ANKRD1, DMD, CXCL9, CASQ2, ACTC1, COL14A1 | 9.94 × 10−6 |
| Skeletal muscle cell differentiation (GO:0035914) | MYLK2, FOS, HLF, ANKRD1, FOXN2 | 9.94 × 10−6 |
| Regulation of muscle system process (GO:0090257) | MYLK2, MSTN, CTGF, DMD, ADRA2C, CASQ2, COL14A1 | 3.72 × 10−5 |
| Collagen metabolic process (GO:0032963) | CTGF, COL1A2, COL1A1, ENG, COL12A1, COL14A1 | 3.72 × 10−5 |
| Response to amino acid (GO:0043200) | CTGF, COL1A2, COL1A1, CDO1, PPARGC1A | 4.07 × 10−5 |
| Cytosolic calcium ion transport (GO:0060401) | CTGF, CXCL10, DMD, CXCL9, THY1, CASQ2 | 6.75 × 10−5 |
| Regulation of muscle tissue development (GO:1901861) | MSTN, PPARGC1A, CXCL10, CXCL9, COL14A1 | 3.70 × 10−4 |
| Regulation of calcium ion transport (GO:0051924) | CXCL10, DMD, CXCL9, ATP2B2, THY1, CASQ2 | 4.40 × 10−4 |
| Actin-myosin filament sliding (GO:0033275) | MYLK2, MYH8, DMD, ACTC1 | 4.53 × 10−4 |
| JNK cascade (GO:0007254) | CTGF, SFRP4, PAK1, TRIB1, DUSP10, GADD45B | 4.53 × 10−4 |
| Negative regulation of protein kinase activity (GO:0006469) | PPP1R1B, THY1, DUSP1, TRIB1, DUSP10, GADD45B | 5.19 × 10−4 |
| Collagen fibril organization (GO:0030199) | COL1A2, COL1A1, COL12A1, COL14A1 | 1.33 × 10−4 |
| Actin-mediated cell contraction (GO:0070252) | MYLK2, MYH8, DMD, ACTC1 | 1.11 × 10−3 |
| Regulation of muscle contraction (GO:0006937) | MYLK2, CTGF, DMD, ADRA2C, CASQ2 | 1.24 × 10−3 |
| Regulation of JNK cascade (GO:0046328) | CTGF, SFRP4, PAK1, DUSP10, GADD45B | 1.27 × 10−3 |
| Regulation of stress-activated MAPK cascade (GO:0032872) | CTGF, SFRP4, PAK1, DUSP10, GADD45B | 2.26 × 10−3 |
| Regulation of stress-activated protein kinase signaling cascade (GO:0070302) | CTGF, SFRP4, PAK1, DUSP10, GADD45B | 2.27 × 10−3 |
| Skeletal muscle tissue growth (GO:0048630) | MSTN, IGFBP5 | 2.97 × 10−3 |
| Regulation of protein kinase B signaling (GO:0051896) | SLC9A3R1, ITSN1, IGFBP5, RASD2 | 3.48 × 10−3 |
| Actin filament bundle organization (GO:0061572) | RHOD, CTGF, PAK1, PFN2 | 4.82 × 10−3 |
| Positive regulation of proteolysis involved in cellular protein catabolic process (GO:1903052) | ZFAND2A, CTSC, TRIB1 | 7.85 × 10−3 |
| Actomyosin structure organization (GO:0031032) | ANKRD1, CASQ2, ACTC1 | 1.61 × 10−2 |
| ID | Molecules in Network | Score | Focus Molecules | Functions |
|---|---|---|---|---|
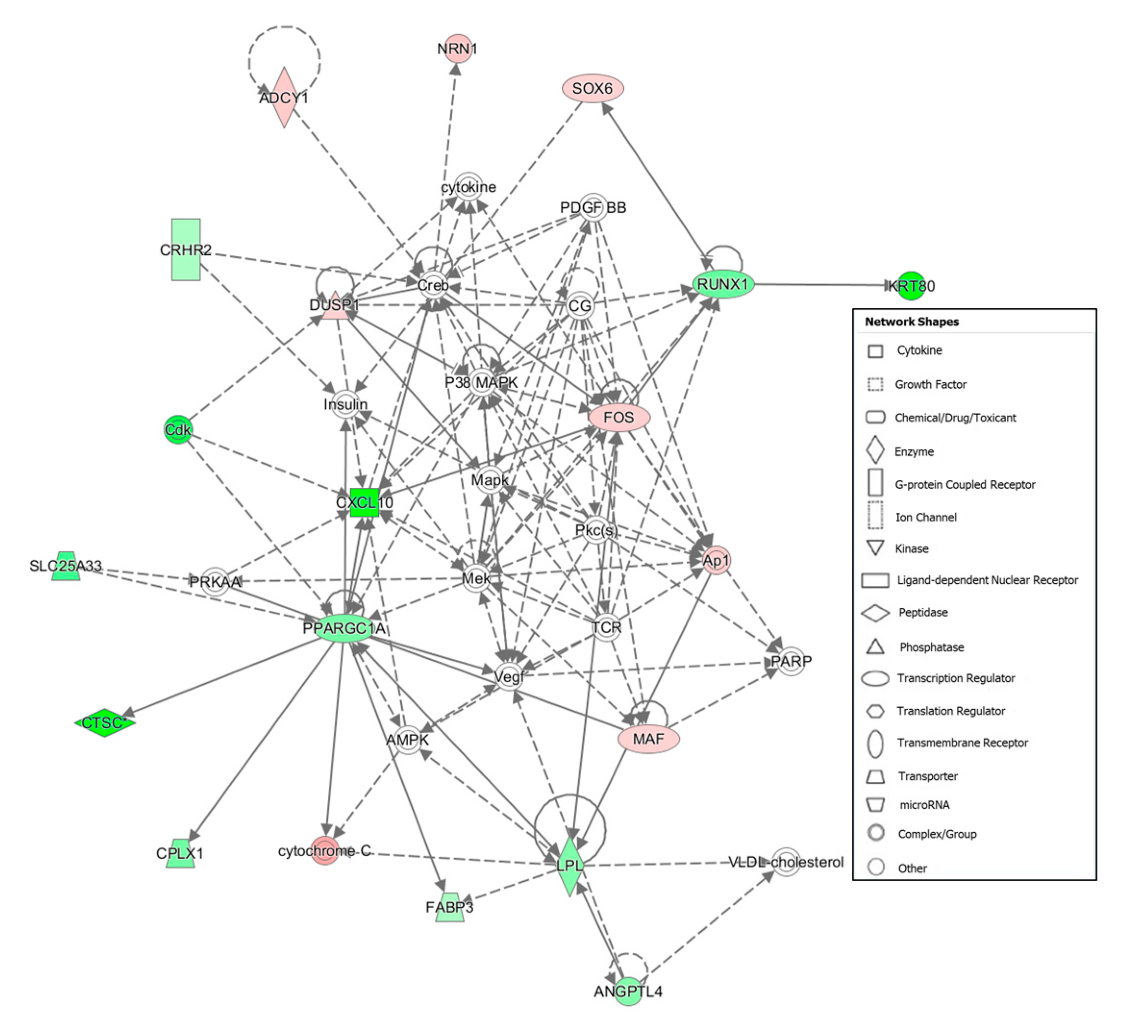
| 5 | ADCY1, AMPK, ANGPTL4, Ap1, Cdk, CG, CPLX1, Creb, CRHR2, CTSC, CXCL10, cytochrome C, cytokine, DUSP1, FABP3, FOS, Insulin, KRT80, LPL, MAF, Mapk, Mek, NRN1, P38 MAPK, PARP, PDGF BB, Pkc(s), PPARGC1A, PRKAA, RUNX1, SLC25A33, SOX6, TCR, Vegf, VLDL-cholesterol | 26 | 17 | Connective Tissue Development and Function, Lipid Metabolism, Tissue Morphology |
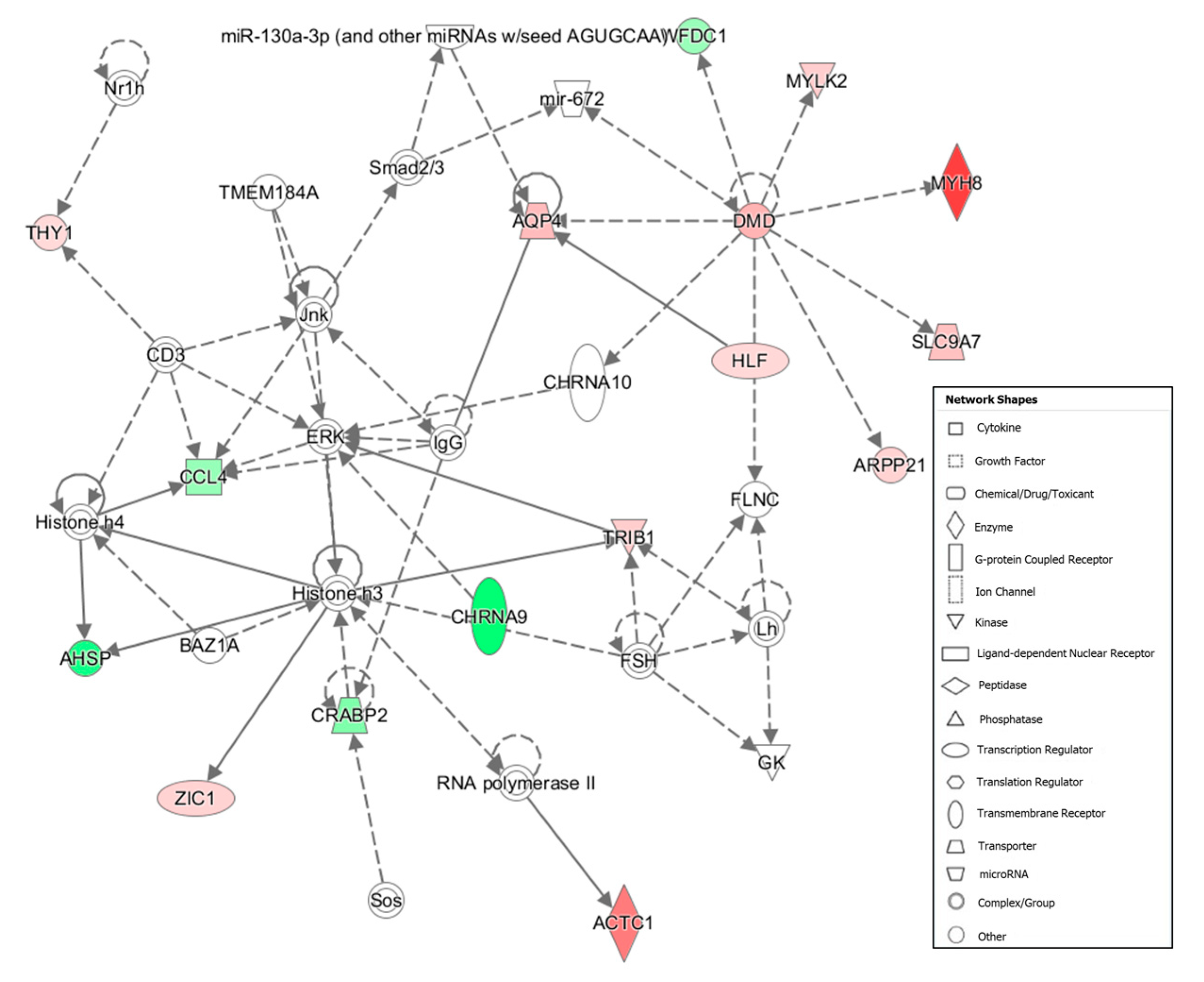
| 6 | ACTC1, AHSP, AQP4, ARPP21, BAZ1A, CCL4, CD3, CHRNA10, CHRNA9, CRABP2, DMD, ERK, FLNC, FSH, GK, Histone h3, Histone h4, HLF, IgG, Jnk, Lh, miR-130a-3p (and other miRNAs w/seed AGUGCAA), mir-672, MYH8, MYLK2, Nr1h, RNA polymerase II, SLC9A7, Smad2/3, Sos, THY1, TMEM184A, TRIB1, WFDC1, ZIC1 | 23 | 16 | Cell Morphology, Cellular Assembly and Organization, Cellular Function and Maintenance |
| Canonical Pathways | p-Value | Ratio | z-Score | Molecules |
|---|---|---|---|---|
| Integrin Signaling | 0.001 | 0.033 | 0.447 | ACTC1, MYLK2, PAK1, PFN2, RASD2, RHOBTB1, RHOD |
| Hepatic Fibrosis Signaling Pathway | 0.001 | 0.027 | 1.265 | CCN2, COL1A1, COL1A2, FOS, MYLK2, RASD2, RHOBTB1, RHOD, TFRC, YAP1 |
| Actin Cytoskeleton Signaling | 0.002 | 0.032 | 1.342 | ACTC1, FN1, MYH8, MYLK2, PAK1, PFN2, RASD2 |
| Synaptogenesis Signaling Pathway | 0.003 | 0.026 | −0.378 | ADCY1, ADCY6, CPLX1, ITSN1, MARCKS, PAK1, RASD2, SNCG |
| PPARα/RXRα Activation | 0.004 | 0.032 | −1.342 | ADCY1, ADCY6, GPD2, LPL, PPARGC1A, RASD2 |
| ILK Signaling | 0.004 | 0.032 | 1.342 | ACTC1, FN1, FOS, MYH8, RHOBTB1, RHOD |
| Tec Kinase Signaling | 0.009 | 0.031 | 1 | ACTC1, FOS, PAK1, RHOBTB1, RHOD |
| Cardiac Hypertrophy Signaling | 0.011 | 0.025 | −1 | ADCY1, ADCY6, ADRA2C, RASD2, RHOBTB1, RHOD |
| GNRH Signaling | 0.011 | 0.029 | 1 | ADCY1, ADCY6, FOS, PAK1, RASD2 |
| Signaling by Rho Family GTPases | 0.012 | 0.025 | 0.447 | ACTC1, CIT, FOS, PAK1, RHOBTB1, RHOD |
| RhoA Signaling | 0.015 | 0.033 | −1 | ACTC1, CIT, MYLK2, PFN2 |
| White Adipose Tissue Browning Pathway | 0.018 | 0.031 | −1 | ADCY1, ADCY6, LDHB, PPARGC1A |
| Upstream Regulator | Molecule Type | PAS | Activation z-Score | p-Value of Overlap | Molecules in Dataset | Related Functions |
|---|---|---|---|---|---|---|
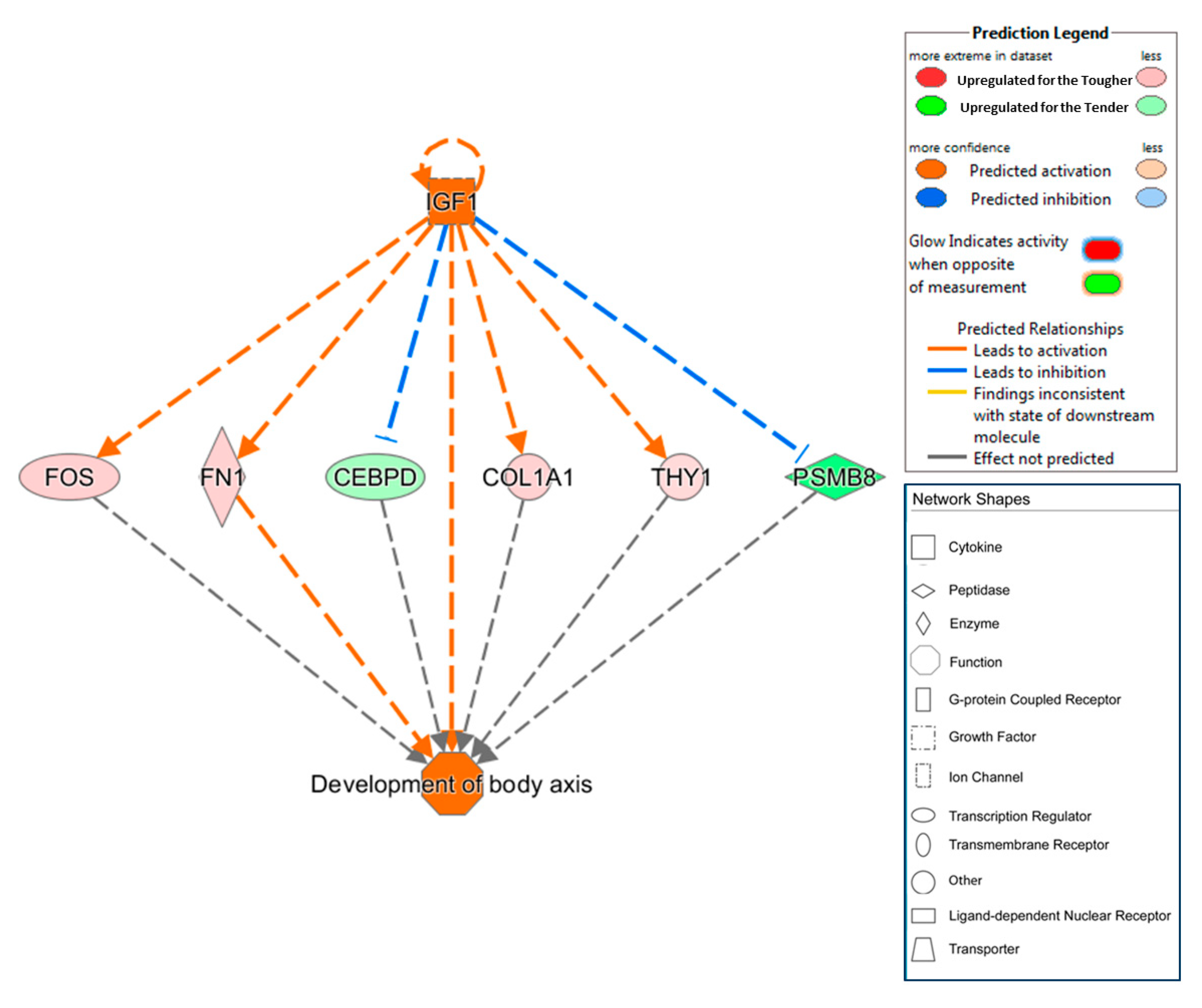
| IGF1 | Growth factor | Activated | 2.947 | 7.29 × 10−8 | CCL4, CEBPD, COL1A1, DUSP1, FN1, FOS, IGFBP5, LPL, MYH8, PSMB8 | Development of body axis |
| VGLL3 | Other | Activated | 2.000 | 1.99 × 10−6 | COL12A1, COL1A1, COL1A2, GADD45B | |
| SEMA7A | Transmembrane receptor | Activated | 2.000 | 3.31 × 10−5 | CCN2, COL1A1, COL1A2, FN1 | |
| PTH | Other | Activated | 2.197 | 4.51 × 10−5 | COL1A1, COL1A2, DUSP1, FOS, IGFBP5, SFRP4 | |
| KLF11 | Transcription regulator | Inhibited | −2.236 | 4.92 × 10−4 | CCN2, COL1A2, CPT2, ENG, FABP3, PPARGC1A | |
| IL4 | Cytokine | Inhibited | −2.331 | 5.93 × 10−4 | ALDOC, CCL26, CCL4, CD163, CXCL10, FOS, LPL, NABP1, PPARGC1A, TFRC | |
| TRIM24 | Transcription regulator | Activated | 2.236 | 1.30 × 10−3 | CXCL10, PSMB10, PSMB8, PSMB9, TAP1 | |
| PPARG | Ligand-dependent nuclear receptor | Inhibited | −2.179 | 1.89 × 10−3 | ANGPTL4, COL1A1, COL1A2, CPT2, CRABP2, FABP3, FN1, IGFBP5, LPL, PPARGC1A | |
| OGT | Enzyme | Inhibited | −2.000 | 2.42 × 10−3 | FOS, LPL, PPARGC1A, THY1 | |
| NOS2 | Enzyme | Inhibited | −2.219 | 3.13 × 10−3 | ACTC1, CCL4, CTSC, CYCS, PPARGC1A, THY1 | |
| SATB1 | Transcription regulator | Activated | 2.000 | 4.18 × 10−2 | GADD45B, HBB, MAF, RUNX1 |
| Gene | Expression Type | qPCR FC | RNAseq FC | r2 | p-Value | CCC |
|---|---|---|---|---|---|---|
| ACTC1 | Tender < Tough | 2.861 | 4.085 | 0.986 | 6.20 × 10−10 | 0.828 |
| MX1 | Tender < Tough | 2.231 | 2.195 | 0.943 | 1.34 × 10−6 | |
| COL1A1 | Tender < Tough | 1.503 | 1.636 | 0.922 | 7.29 × 10−6 | |
| FOS | Tender < Tough | 1.634 | 1.670 | 0.913 | 1.29 × 10−5 | |
| ANKRD1 | Tender > Tough | 0.531 | 0.476 | 0.779 | 0.002 | |
| MSTN | Tender < Tough | 1.753 | 2.038 | 0.527 | 0.064 | |
| IRF1 | NO DE | 0.656 | 0.235 | 0.997 | 1.81 × 10−13 | |
| NOS2 | NO DE | 0.611 | 0.811 | 0.701 | 0.008 | |
| SSH2 | NO DE | 1.037 | 1.625 | 0.584 | 0.036 | |
| ELOVL6 | NO DE | 0.887 | 1.712 | 0.565 | 0.044 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández-Barroso, M.Á.; Caraballo, C.; Silió, L.; Rodríguez, C.; Nuñez, Y.; Sánchez-Esquiliche, F.; Matos, G.; García-Casco, J.M.; Muñoz, M. Differences in the Loin Tenderness of Iberian Pigs Explained through Dissimilarities in Their Transcriptome Expression Profile. Animals 2020, 10, 1715. https://doi.org/10.3390/ani10091715
Fernández-Barroso MÁ, Caraballo C, Silió L, Rodríguez C, Nuñez Y, Sánchez-Esquiliche F, Matos G, García-Casco JM, Muñoz M. Differences in the Loin Tenderness of Iberian Pigs Explained through Dissimilarities in Their Transcriptome Expression Profile. Animals. 2020; 10(9):1715. https://doi.org/10.3390/ani10091715
Chicago/Turabian StyleFernández-Barroso, Miguel Ángel, Carmen Caraballo, Luis Silió, Carmen Rodríguez, Yolanda Nuñez, Fernando Sánchez-Esquiliche, Gema Matos, Juan María García-Casco, and María Muñoz. 2020. "Differences in the Loin Tenderness of Iberian Pigs Explained through Dissimilarities in Their Transcriptome Expression Profile" Animals 10, no. 9: 1715. https://doi.org/10.3390/ani10091715
APA StyleFernández-Barroso, M. Á., Caraballo, C., Silió, L., Rodríguez, C., Nuñez, Y., Sánchez-Esquiliche, F., Matos, G., García-Casco, J. M., & Muñoz, M. (2020). Differences in the Loin Tenderness of Iberian Pigs Explained through Dissimilarities in Their Transcriptome Expression Profile. Animals, 10(9), 1715. https://doi.org/10.3390/ani10091715