On the Behavioural Biology of the Mainland Serow: A Comparative Study



Simple Summary
Abstract
1. Introduction
2. Materials and Methods
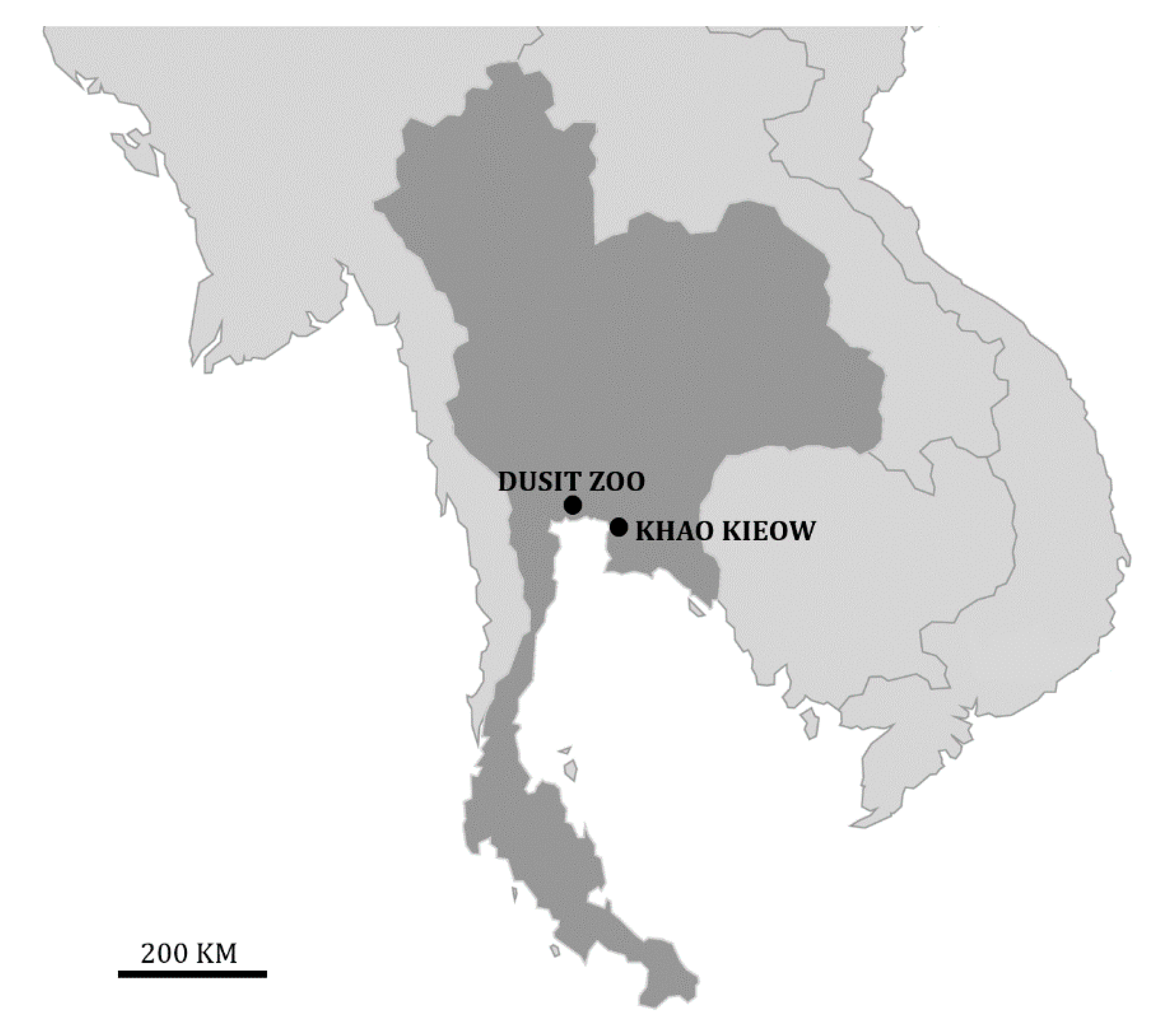
2.1. Study Area
2.2. Data Collection
2.3. Data Analysis
3. Results
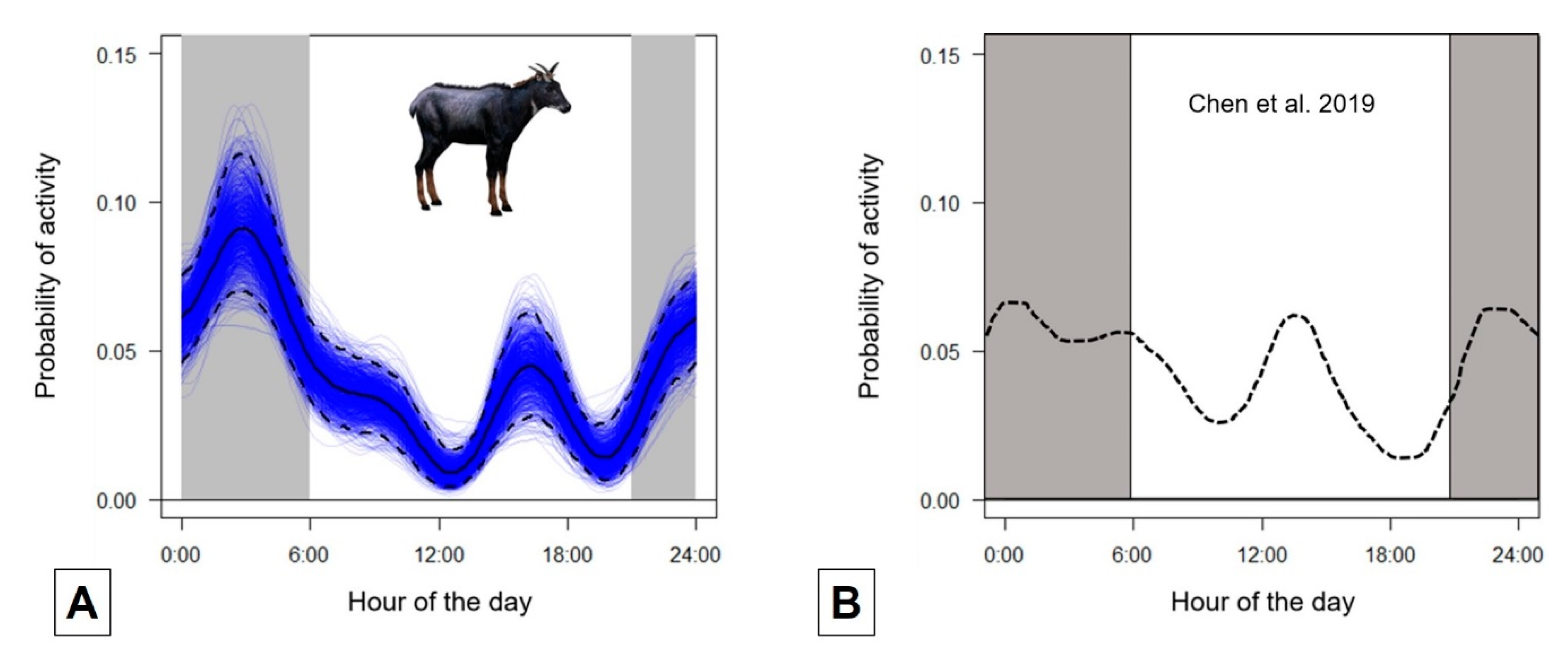
3.1. Activity Rhythms and Marking Behaviour
3.2. Behavioural Repertoire
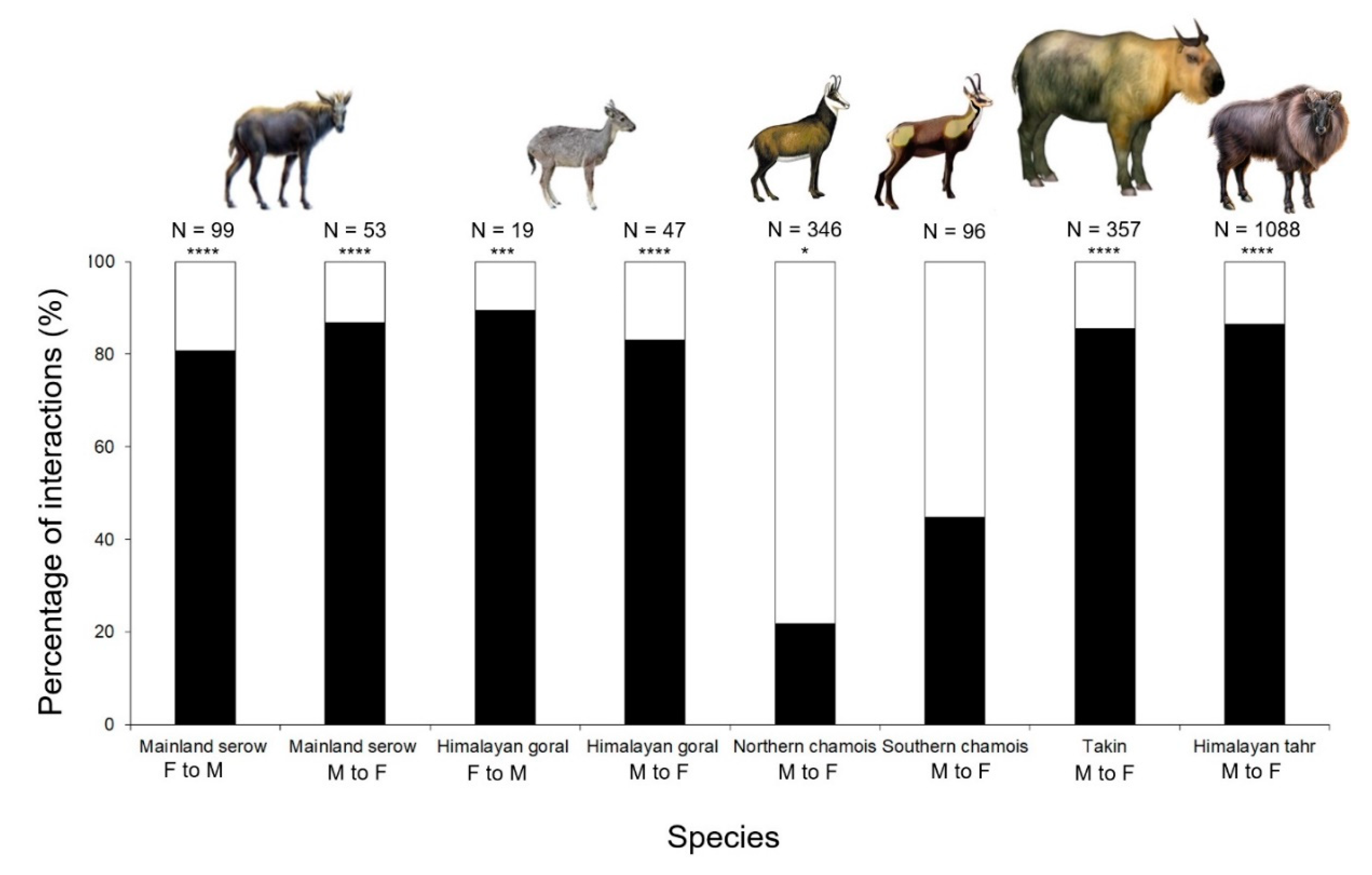
3.3. Direct and Indirect Forms
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Alcock, J. Animal Behavior: An Evolutionary Perspective; Sinauer Editions: Sunderland, MA, USA, 1984. [Google Scholar]
- Noble, G.K. The role of dominance in the social life of birds. Auk 1939, 56, 263–273. [Google Scholar] [CrossRef]
- Owen-Smith, N. On territoriality in ungulates and an evolutionary model. Quart. Rev. Biol. 1977, 52, 1–38. [Google Scholar] [CrossRef]
- Mead, G.H. Concerning animal perception. Psychol. Rev. 1907, 14, 383. [Google Scholar] [CrossRef]
- Kleiman, D.G.; Eisenberg, J.F. Comparisons of canid and felid social systems from an evolutionary perspective. Anim. Behav. 1973, 21, 637–659. [Google Scholar] [CrossRef]
- Johnson, R.P. Scent marking in mammals. Anim. Behav. 1973, 21, 521–535. [Google Scholar] [CrossRef]
- Barrette, C. Scent-marking in captive muntjacs, Muntiacus reevesi. Anim. Behav. 1977, 25, 536–541. [Google Scholar] [CrossRef]
- Gosling, L.M. A reassessment of the function of scent marking in territories. Zeitschr. Tierpsychol. 1982, 60, 89–118. [Google Scholar] [CrossRef]
- Walther, F.R. Communication and Expression in Hoofed Mammals; Indiana University Press: Bloomington, IN, USA, 1984. [Google Scholar]
- Gosling, L.M. The evolution of mating strategies in male antelopes. In Ecological Aspects of Social Evolution; Rubestein, D.I., Wrangham, R.W., Eds.; Princeton University Press: Princeton, NJ, USA, 1986; pp. 244–281. [Google Scholar]
- Schaller, G.B. Mountain Monarchs. Wild Sheep and Goats of the Himalaya; University of Chicago Press: Chicago/London, UK, 1977. [Google Scholar]
- Geist, V. On the evolution of the Caprinae. In The Biology and Management of Capricornis and Related Mountain Antelopes; Soma, H., Ed.; Croom-Helm: London, UK, 1987; pp. 134–144. [Google Scholar]
- Von Hardenberg, A.; Bassano, B.; Peracino, A.; Lovari, S. Male Alpine chamois occupy territories as hotspots before the mating season. Ethology 2000, 106, 617–630. [Google Scholar] [CrossRef]
- Corlatti, L.; Béthaz, S.; von Hardenberg, A.; Bassano, B.; Palme, R.; Lovari, S. Hormones, parasites and male mating tactics in Alpine chamois: Identifying the mechanisms of life history trade-offs. Anim. Behav. 2012, 84, 1061–1070. [Google Scholar] [CrossRef]
- Munton, P.N. The ecology of the Arabian Tahr (Hemitragus jayakari Thomas, 1894) and a strategy for the conservation of the species. J. Oman Stud. 1985, 8, 11–48. [Google Scholar]
- Akasaka, T.; Maruyama, N. Social organization and habitat use of Japanese serow in Kasabori. J. Mammal. Soc. Jpn. 1977, 7, 87–102. [Google Scholar]
- Kishimoto, R.; Kawamichi, T. Territoriality and monogamous pairs in a solitary ungulate, the Japanese serow, Capricornis crispus. Anim. Behav. 1996, 52, 673–682. [Google Scholar]
- Myslenkov, A.I.; Voloshina, I.V. Sexual behavior of Amur goral. Proc. Sec. World Conf. Mt. Ungulates 1998, 1, 75–80. [Google Scholar]
- Lovari, S.; Apollonio, M. On the rutting behaviour of the Himalayan goral Nemorhaedus goral (Hardwicke, 1825). J. Ethol. 1994, 12, 25–34. [Google Scholar]
- Mori, E.; Nerva, L.; Lovari, S. Reclassification of the serows and the gorals: The end of a never-ending story? Mammal Rev. 2019, 49, 256–262. [Google Scholar]
- Phan, T.D.; Nijhawan, S.; Li, S.; Xiao, L. Mainland serow Capricornis sumatraensis. IUCN Glob. Mammal Assess. 2020. [Google Scholar] [CrossRef]
- Lekagul, B.; McNeely, J.A. Mammals of Thailand. Association for the Conservation of Wildlife; Kurusapa Ladproa Press Editions: Bangkok, Thailand, 1977; pp. 715–719. [Google Scholar]
- Chen, Y.; Xiao, Z.; Zhang, L.; Wang, X.; Li, M.; Xiang, Z. Activity rhythms of coexisting red serow and Chinese serow at Mt. Gaoligong as identified by camera traps. Animals 2019, 9, 1071. [Google Scholar] [CrossRef]
- Davis, E.O.; Glikman, J.A. An assessment of wildlife use by Northern Laos nationals. Animals 2020, 10, 685. [Google Scholar] [CrossRef]
- Lovari, S.; Locati, M. Site features of territorial dung-marking in mainland serow. Mammalia 1994, 58, 153–156. [Google Scholar]
- Aryal, A. Habitat ecology of Himalayan serow (Capricornis sumatraensis ssp. thar) in Annapurna Conservation area of Nepal. Tiger Pap. 2009, 34, 12–20. [Google Scholar]
- Chen, W.; Chu Hu, J.; Li, X. Habitat use and separation between the Chinese serow (Capricornis milneedwardsi) and the Chinese goral (Naemorhedus griseus) in winter. Mammalia 2009, 73, 249–252. [Google Scholar] [CrossRef]
- Giri, S.; Aryal, A.; Koirala, R.K.; Adhikari, B.; Raubenheimer, D. Feeding ecology and distribution of Himalayan serow (Capricornis thar) in Annapurna Conservation Area, Nepal. World J. Zool. 2011, 6, 80–85. [Google Scholar]
- Bhattacharya, T.; Bashir, T.; Poudyal, K.; Sathyakumar, S.; Saha, G.K. Distribution, occupancy and activity patterns of goral (Nemorhaedus goral) and serow (Capricornis thar) in Khangchendzonga Biosphere Reserve, Sikkim, India. Mammal Study 2012, 37, 173–181. [Google Scholar] [CrossRef]
- Lovari, S.; Pellizzi, B.; Boesi, R.; Fusani, L. Mating dominance amongst male Himalayan tahr: Blonds do better. Behav. Proc. 2009, 81, 20–25. [Google Scholar] [CrossRef] [PubMed]
- West, J. Notes on the Sumatran serow Capricornis sumatraensis at Jakarta Zoo. Intern. Zoo Yearb. 1979, 19, 252–254. [Google Scholar] [CrossRef]
- Chokngamwong, R.; Chiu, L.S. Thailand daily rainfall and comparison with TRMM products. J. Hydrometeorol. 2008, 9, 256–266. [Google Scholar] [CrossRef]
- Meredith, M.; Ridout, M. Overview of the Overlap Package. 2014. Available online: http://cran.cs.wwu.edu/web/packages/overlap/vignettes/overlap.pdf (accessed on 24 April 2020).
- IBM Corp. IBM SPSS Statistics for Windows, Version 22.0; IBM Corp: Armonk, NY, USA, 2013. [Google Scholar]
- Lovari, S. Behavioural repertoire of the Abruzzo chamois, Rupicapra pyrenaica Neumann, 1899 (Artiodactyla: Bovidae). Saugetierkundl. Mitt. 1985, 32, 113–136. [Google Scholar]
- Lovari, S.; Fattorini, N.; Boesi, R.; Bocci, A. Male ruff colour as a rank signal in a monomorphic-horned mammal: Behavioural correlates. Sci. Nat. 2015, 102, 39. [Google Scholar] [CrossRef]
- McDonald, J.H. Handbook of Biological Statistics; Sparky House Publishing: Baltimore, MD, USA, 2014. [Google Scholar]
- Sokal, R.R.; Rohlf, F.J. Introduction to Biostatistics; Freeman and Co.: New York, NY, USA, 1987. [Google Scholar]
- Sakurai, M. Socio-ecological study of the Japanese serow, Capricornis crispus (Mammalia; Bovidae), with special reference to the flexibility of its social structure. Physiol. Ecol. Jpn. 1981, 18, 163–212. [Google Scholar]
- van Schaik, C.P.; Griffiths, M. Activity periods of Indonesian rain forest mammals. Biotropica 1996, 28, 105–112. [Google Scholar] [CrossRef]
- Duckworth, J.W.; Steinmetz, R.; Pattanavibool, A. Capricornis milneedwardsii. The IUCN Red List of Threatened Species 2008: E.T3814A10101852. Available online: https://dx.doi.org/10.2305/IUCN.UK.2008.RLTS.T3814A10101852.en (accessed on 22 May 2020).
- Pao-Chung, C. Breeding and behaviour of Formosan serow at Taipei Zoo. In The Biology and Management of Capricornis and Related Mountain Antelopes; Soma, H., Ed.; Croom-Helm: London, UK, 1987; pp. 154–164. [Google Scholar]
- Nash, L.T.; Chilton, S.M. Space or novelty? Effects of altered cage size on Galago behavior. Am. J. Primat. 1986, 10, 37–49. [Google Scholar]
- Cassinello, J.; Pieters, I. Multi-male captive groups of endangered dama gazelle: Social rank, aggression, and enclosure effects. Zoo Biol. 2000, 19, 121–129. [Google Scholar]
- Geist, V. On the relationship of social evolution and ecology in ungulates. Am. Zool. 1974, 14, 205–220. [Google Scholar]
- Tinbergen, N. The Study of Instinct; Oxford University Press: London, UK, 1951. [Google Scholar]
- Geist, V. Mountain Sheep. A Study in Behavior and Evolution; University of Chicago Press: Chicago, IL, USA, 1971. [Google Scholar]
- Wilson, E.O. Sociobiology; Belknap/Harvard University Press: Cambridge, MA, USA, 1975. [Google Scholar]
- Estes, R. The role of vomeronasal organ in mammalian reproduction. Mammalia 1972, 36, 315–341. [Google Scholar]
- Geist, V. On the rutting behavior of the mountain goat. J. Mammal. 1964, 45, 551. [Google Scholar]
- Ale, S.B. The Rutting Behaviour of Male Blue Sheep in the Manang Valley, Nepal. Master’s Thesis, University of Tromsø, Tromsø, Norway, 1999. [Google Scholar]
- Altmann, D. Verhaltensstudien am Mishmi-Takin (Budorcas taxicolor taxicolor) in Tierpark Berlin. Milu Berl. 1980, 5, 342–358. [Google Scholar]
- Cavallini, P. On the behaviour of male Sardinian mouflons (Ovis orientalis musimon) during the pre-rut. Mammalia 1987, 51, 195–200. [Google Scholar] [CrossRef]
- Masini, F.; Lovari, S. Systematics, phylogenetic relationships, and dispersal of the chamois (Rupicapra spp.). Quatern. Res. 1988, 30, 339–349. [Google Scholar]
- Locati, M.; Lovari, S. Sexual differences in aggressive behaviour of the Apennine chamois. Ethology 1990, 84, 295–306. [Google Scholar] [CrossRef]
- Lovari, S.; Locati, M. Temporal relationships, transitions and structure of the behavioural repertoire in male Apennine chamois during the rut. Behaviour 1991, 119, 77–103. [Google Scholar]
- Fournier, F.; Festa-Bianchet, M. Social dominance in adult female mountain goats. Anim. Behav. 1995, 49, 1449–1459. [Google Scholar]
- Festa-Bianchet, M.; Cote, S.D. Mountain Goats: Ecology, Behaviour, and Conservation of an Alpine Ungulate; Island Press: New York, NY, USA, 2008. [Google Scholar]
- Corlatti, L.; Caroli, M.; Pietrocini, V.; Lovari, S. Rutting behaviour of territorial and nonterritorial male chamois: Is there a home advantage? Behav. Proc. 2013, 92, 118–124. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marking Type | M1 | F1 | M2 | F2 | ||||
|---|---|---|---|---|---|---|---|---|
| n | % | n | % | n | % | n | % | |
| Preorbital Gland | 1216 | 88.1 | 258 | 77.3 | 90 | 88.2 | 38 | 45.2 |
| Forehead | 125 | 9.1 | 34 | 10.2 | 5 | 4.9 | 12 | 14.3 |
| Horn/Nape | 13 | 0.9 | 17 | 5.1 | 6 | 5.9 | 19 | 22.6 |
| Flank | 26 | 1.9 | 25 | 7.5 | 1 | 1 | 15 | 17.9 |
| Total | 1380 | 334 | 102 | 84 | ||||
| Behaviour Pattern | Description |
|---|---|
| Approaching (M, F) | Threat. The sender walks straight to the receiver. The mane may be erected (D). |
| Body/Head Shaking (F) | Dominance display. The sender vigorously shakes the head several times (I). |
| Chasing (F) | Threat. The female chases the male for a few metres. Performed only by F1 (D). |
| Clashing (M, F) | Threat. Two individuals front each other, with lowered head, and clash after a short rush. Sometimes alternated with Front Pushing (D). |
| Conflict Posture (M) | Dominance display. It shows elements of submissive behaviour mixed to dominant ones; mouth may be slightly open; ears may be turned outwardly, slightly drooping (I). |
| Frontal Pushing (M, F) | Threat. Two individuals move their foreheads into contact and start pushing (D). Sometimes preceded by Head Down (D). |
| Gamboling (M, F) | Dominance display. A downhill run that includes vigorous head nodding, alternately lifting up forequarters and hindquarters (I). |
| Head Butting (M, F) | Threat. An individual butts the opponent’s body with its forehead (D). |
| Head Down (M, F) | Threat (Figure 3C). An individual holds its head lower than main axis of body, pointing horns to another one. The mane is erected. It may be performed laterally or frontally (D). |
| Hooking (M, F) | Threat. One attempts to gore another with a horn sweep. It may be frontal (to shoulder/neck of opponent) or side-on (to rump/abdomen of opponent) (D). |
| Horning Vegetation (M, F) | Dominance display. An individual horns a bush vigorously and/or lay scent on it (I). |
| Licking Body (M, F) | Socially explorative behaviour [9]. An individual licks the other’s body. Licked areas are generally muzzle, flank, and genitals. |
| Naso-Nasal Contact (M, F) | Socially explorative behaviour. An individual touches the nasal region of the other with its nose. |
| Rushing (F) | Threat. An individual rushes quickly to the other one. Head position may vary. Performed only by F1 (D). |
| Side Display (M) | Dominance display (I). An individual stands on stiffly stretched legs, broadside on; back is hunched, head is lowered as in Head Down (D). |
| Sniffing Forehead/Hoof (M, F) | Socially explorative behaviour. An individual sniffs glands of the other. Sniffing may help partner recognition. |
| Snorting (F) | Threat. A sound uttered by a violent puff through nostrils, mainly during Chasing and Rushing (D). |
| Staring (M, F) | Threat (Figure 3G). An individual stares the other one, with mane erected (D). |
| Behaviour Pattern | Description |
|---|---|
| Courtship Foot Stamping (M) | Male stamps the ground with one of his fore-hooves, sometimes quickly alternating with both of them, behind the female. Mane can be erected (Figure 3D). |
| Croup Touch (M) | Male rests his chin, throat, and lower part of the neck on the female’s croup. Often preceded or followed by Kicking. |
| Flank Stroke (M) | The male softly strokes the female’s hindquarters with the inner surface of one of his forelegs in a downward sweep. |
| Following (M) | Male follows female closely. |
| Head Up (M) | Male approaches the female with stiff legs, neck raised, and uplifted muzzle. Tongue flicks may be performed by the male. |
| Kicking (M, F) | An individual, the male generally kicks the other’s flank, abdomen, or hind-legs with a stiff foreleg. It may be performed frontally, laterally, or from behind. |
| Licking Genitals/Urine (M, F) | An individual licks the other one’s genitals or urine. Individuals can lick each other simultaneously. It may help male to verify female oestrus stage through her pheromone odour. |
| Lip-Curling (M) | Male sniffs female’s vulva or urine, slowly lifting his muzzle and curling the upper lip. It helps scent perception by activation of the Jacobson’s organ. |
| Low Stretching (M) | Submissive display. Male approaches female with quick steps—or it may stand still—on flexed carpal joints, with a lowered and outstretched neck, with chin slightly raised. Horns partly concealed in mane (Figure 3E). |
| Mounting (M) | Copulative behaviour. Female stands with rear legs slightly spread and tail upraised, while male rises up on hindlegs and partly rests on top of female. Not all mounts end up in copulations, as female may withdraw (cf. Intentional Mounting). |
| Intentional Mounting (M) | Copulative behaviour. Male attempts to mount female, unsuccessfully. |
| Naso-Genital Contact (M) | Male sniffs the genital region of the female. |
| Sniffing Urine (M) | Male sniffs the area where female has urinated. Usually, Lip-Curling follows. |
| Tongue Flicking (M) | Male, behind female, from the Low Stretching posture, flicks out his tongue repeatedly, with head and neck in Low Stretching. |
| Twisting (M) | The male lowers his neck into a position that looks like the Low Stretching and then rotates his head up to 90° so that horns face away from the female (Figure 3F). |
| Behaviour Pattern | Sender | |||
|---|---|---|---|---|
| % M1 | % M2 | % F1 | % F2 | |
| Approaching | 2.8 | 1.3 | 3.2 | 1.2 |
| Body/Head Shaking | 0.0 | 0.0 | 7.9 | 0.0 |
| Chasing | 0.0 | 0.0 | 1.6 | 0.0 |
| Clashing | 1.4 | 0.5 | 1.6 | 3.7 |
| Conflict Posture | 1.4 | 0.0 | 0.0 | 0.0 |
| Croup Touching | 0.0 | 5.0 | 0.0 | 0.0 |
| Flank Stroking | 0.0 | 1.6 | 0.0 | 1.2 |
| Following | 0.7 | 0.5 | 0.0 | 0.0 |
| Foot Stamping | 3.5 | 1.6 | 0.0 | 0.0 |
| Frontal Pushing | 2.8 | 0.5 | 3.2 | 0.0 |
| Gamboling | 0.0 | 0.3 | 11.1 | 3.7 |
| Head Butting | 0.0 | 0.3 | 0.0 | 4.9 |
| Head Down | 7.8 | 2.6 | 20.6 | 21.0 |
| Head Up | 12.1 | 2.1 | 0.0 | 0.0 |
| Hooking | 0.0 | 0.5 | 7.9 | 7.4 |
| Horning Vegetation | 0.7 | 0.0 | 4.8 | 1.2 |
| Intentional Mounting | 0.7 | 6.3 | 0.0 | 0.0 |
| Kicking | 36.9 | 28.2 | 0.0 | 1.2 |
| Licking Body | 0.7 | 0.0 | 0.0 | 1.2 |
| Licking Genitals/Urine | 3.5 | 7.4 | 0.0 | 25.9 |
| Lip-Curling | 5.0 | 0.8 | 0.0 | 0.0 |
| Low Stretching | 6.4 | 1.6 | 0.0 | 0.0 |
| Mounting | 0.0 | 29.7 | 0.0 | 0.0 |
| Naso-Genital Contact | 5.0 | 2.9 | 0.0 | 0.0 |
| Naso-Nasal Contact | 2.1 | 1.8 | 4.8 | 9.9 |
| Rushing | 0.0 | 0.0 | 11.1 | 0.0 |
| Side Display | 0.0 | 0.8 | 0.0 | 0.0 |
| Sniffing Forehead/Hoof | 2.8 | 0.3 | 0.0 | 13.6 |
| Sniffing Urine | 2.1 | 1.1 | 0.0 | 0.0 |
| Snorting | 0.0 | 0.0 | 4.8 | 1.2 |
| Staring | 1.4 | 0.0 | 17.5 | 2.5 |
| Tongue Flicking | 0.0 | 1.3 | 0.0 | 0.0 |
| Twisting | 0.0 | 1.1 | 0.0 | 0.0 |
| Behaviour Pattern | Mainland Serow | Japanese Serow | Himalayan Goral | Northern Chamois | Southern Chamois | Takin | Himalayan Tahr |
|---|---|---|---|---|---|---|---|
| Approaching | + | - | + | + | + | + | + |
| Blocking | - | - | - | - | - | - | + |
| Body/Head Shaking | (+) | - | (+) | + | + | (+) | + |
| Bunting (Mock-Suck) | - | - | - | - | + | - | - |
| Chasing | (+) | + | + | + | + | + | + |
| Clashing | + | + | - | (+) | (+) | - | + |
| Conflict Posture | (+) | ? | - | - | + | - | - |
| Croup Touching | + | + | + | ? | - | + | + |
| Flank Stroking | (+) | - | + | - | + | (+) | - |
| Following | (+) | + | + | ? | ? | + | - |
| Foot Stamping/Herding | + | - | - | + | - | - | + |
| Frontal Pushing | + | + | - | - | - | + | + |
| Gamboling | + | - | + | + | + | - | - |
| Head Butting | (+) | + | + | + | + | - | + |
| Head Down (Static) | + | + | + | + | + | - | + |
| Head Down (Dynamic) | - | - | + | + | + | - | + |
| Head Up | + | + | + | + | + | - | - |
| Hopping | - | (+) | - | + | + | - | - |
| Hooking | + | + | + | + | + | - | + |
| Horning/marking Vegetation | (+) | + | + | + | + | + | + |
| Humping | - | - | - | - | - | - | + |
| Intentional Mounting | + | + | + | + | + | + | + |
| Kicking | + | + | + | (+) | - | + | - |
| Licking body | + | + | + | + | + | - | - |
| Licking genitals/urine | + | + | + | + | + | + | + |
| Lip-Curling | + | + | + | + | + | + | + |
| Low Stretching | + | + | + | + | + | - | + |
| Mounting | + | + | + | + | + | + | + |
| Naso-Genital Contact | (+) | + | + | (+) | (+) | + | + |
| Naso-Nasal Contact | + | + | + | (+) | + | + | + |
| Neck-Fighting | - | - | - | + | (+) | + | - |
| Neck-Up | - | - | + | + | + | + | - |
| Penile Display | - | - | - | + | + | + | + |
| Poking | - | - | - | + | - | - | + |
| Rushing | + | (+) | + | + | + | + | + |
| Side Display | (+) | - | - | + | + | + | - |
| Snorting | + | + | + | + | + | + | - |
| Sniffing Forehead/Hoof | + | + | - | - | - | - | - |
| Sniffing Urine | + | - | - | - | - | - | + |
| Staring | (+) | - | + | + | + | + | + |
| Male Urinating in Female Posture | - | - | - | + | + | - | - |
| Tongue Flicking | (+) | - | - | - | - | - | + |
| Twisting | (+) | - | - | - | - | - | - |
| TOTAL | 33 | 23 | 26 | 31 | 31 | 22 | 27 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lovari, S.; Mori, E.; Procaccio, E.L. On the Behavioural Biology of the Mainland Serow: A Comparative Study. Animals 2020, 10, 1669. https://doi.org/10.3390/ani10091669
Lovari S, Mori E, Procaccio EL. On the Behavioural Biology of the Mainland Serow: A Comparative Study. Animals. 2020; 10(9):1669. https://doi.org/10.3390/ani10091669
Chicago/Turabian StyleLovari, Sandro, Emiliano Mori, and Eva Luna Procaccio. 2020. "On the Behavioural Biology of the Mainland Serow: A Comparative Study" Animals 10, no. 9: 1669. https://doi.org/10.3390/ani10091669
APA StyleLovari, S., Mori, E., & Procaccio, E. L. (2020). On the Behavioural Biology of the Mainland Serow: A Comparative Study. Animals, 10(9), 1669. https://doi.org/10.3390/ani10091669