The Contribution of Mutual Grooming to Affiliative Relationships in a Feral Misaki Horse Herd


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites, Target Herd, and Study Period
2.2. Behavioral Data Collection and Treatment
2.2.1. Kinship
2.2.2. Grooming
2.2.3. Aggression
2.2.4. Proximity Matrices, Social Rank, and Social Network Analysis
2.3. Statistical Analyses
2.4. Ethical Approval
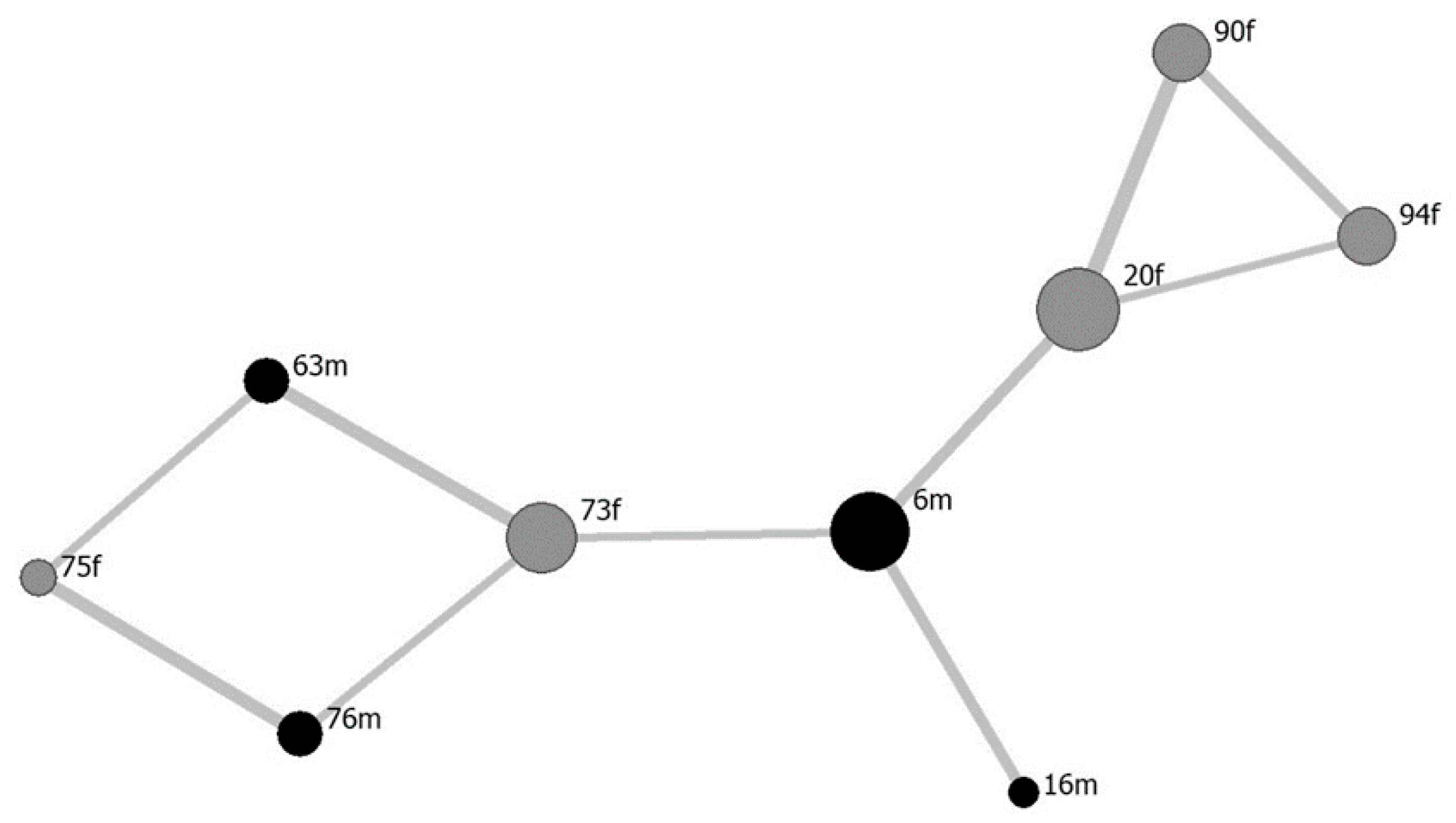
3. Results
3.1. Mutual Grooming Interaction Structure
3.2. Social Relationships and Interactions Among “6m” Herd Members
3.3. Tau Kr Test
4. Discussion
4.1. Social Structure and Social Ranking in the “6m” Herd
4.2. Mutual Grooming And Affiliative Relationships between Horses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Silk, J.B. The adaptive value of sociality in mammalian groups. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2007, 362, 539–559. [Google Scholar] [CrossRef] [PubMed]
- Cameron, E.Z.; Setsaas, T.H.; Linklater, W.L. Social bonds between unrelated females increase reproductive success in feral horses. Proc. Natl. Acad. Sci. USA 2009, 106, 13850–13853. [Google Scholar] [CrossRef] [PubMed]
- Costa, H.; Fragoso, S.; Heitor, F. The relevance of affiliative relationships in horses: Review and future directions. Pet Behav. Sci. 2019, 8, 11–26. [Google Scholar] [CrossRef]
- Seyfarth, R.M. A model of social grooming among adult female monkeys. J. Theor. Biol. 1977, 65, 671–698. [Google Scholar] [CrossRef]
- Schino, G.; Aureli, F. Primate reciprocity and its cognitive requirements. Evol. Anthr. Iss News Rev. 2010, 19, 130–135. [Google Scholar] [CrossRef]
- Silk, J.B.; Beehner, J.C.; Bergman, T.J.; Crockford, C.; Engh, A.L.; Moscovice, L.R.; Wittig, R.M.; Seyfarth, R.M.; Cheney, D.L. Female chacma baboons form strong, equitable, and enduring social bonds. Behav. Ecol. Sociobiol. 2010, 64, 1733–1747. [Google Scholar] [CrossRef] [PubMed]
- Furuya, Y. Grooming behavior in the wild Japanese monkeys. Primates 1957, 1, 47–72. [Google Scholar] [CrossRef]
- Inoue, N.; Shimada, M. Comparison of the activity budget, interactions, and social structure of captive and wild chimpanzees. Animals 2020, 10, 1063. [Google Scholar] [CrossRef]
- Furuichi, T. Symmetrical patterns in non-agonistic social interactions found in unprovisioned Japanese macaques. J. Ethol. 1984, 2, 109–119. [Google Scholar] [CrossRef]
- Muroyama, Y. Mutual reciprocity of grooming in female japanese macaques (Macaca Fuscata). Behaviour 1991, 119, 161–170. [Google Scholar] [CrossRef]
- Gomes, C.M.; Mundry, R.; Boesch, C. Long-term reciprocation of grooming in wild West African chimpanzees. Proc. Biol. Sci. 2009, 276, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Mills, D.S.; McDonnell, S.M. The Domestic Horse: The Origins, Development and Management of its Behaviour; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2005. [Google Scholar]
- Salter, R.E.; Hudson, R.J. Feeding Ecology of Feral Horses in Western Alberta. J. Range Manag. 1979, 32, 221. [Google Scholar] [CrossRef]
- Feh, C.; de Mazières, J. Grooming at a preferred site reduces heart rate in horses. Anim. Behav. 1993, 46, 1191–1194. [Google Scholar] [CrossRef]
- Ringhofer, M.; Inoue, S.; Mendonça, R.S.; Pereira, C.; Matsuzawa, T.; Hirata, S.; Yamamoto, S. Comparison of the social systems of primates and feral horses: Data from a newly established horse research site on Serra D’Arga, northern Portugal. Primates 2017, 58, 479–484. [Google Scholar] [CrossRef]
- Kimura, R. Mutual grooming and preferred associate relationships in a band of free-ranging horses. Appl. Anim. Behav. Sci. 1998, 59, 265–276. [Google Scholar] [CrossRef]
- Linklater, W.L.; Cameron, E.Z.; Stafford, K.J.; Veltman, C.J. Social and spatial structure and range use by Kaimanawa wild horses (Equus caballus: Equidae). New Zealand J. Ecol. 2000, 24, 139–152. [Google Scholar]
- Houpt, K.A.; Law, K.; Martinisi, V. Dominance hierarchies in domestic horses. Appl. Anim. Ethol. 1978, 4, 273–283. [Google Scholar] [CrossRef]
- Vervaecke, H.; Stevens, J.M.G.; Vandemoortele, H.; Sigurjónsdóttir, H.; De Vries, H. Aggression and dominance in matched groups of subadult Icelandic horses (Equus caballus). J. Ethol. 2006, 25, 239–248. [Google Scholar] [CrossRef]
- Hartmann, E.; Christensen, J.W.; McGreevy, P.D. Dominance and Leadership: Useful Concepts in Human–Horse Interactions? J. Equine Vet. Sci. 2017, 52, 1–9. [Google Scholar] [CrossRef]
- Kaseda, Y. The structure of the groups of misaki horses in toi cape. Nihon Chikusan Gakkaiho 1981, 52, 227–235. [Google Scholar] [CrossRef]
- Khalil, A.; Murakami, N. Factors Affecting the Harem Formation Process by Young Misaki Feral Stallions. J. Vet. Med. Sci. 1999, 61, 667–671. [Google Scholar] [CrossRef] [PubMed]
- McDonnell, S.M.; Haviland, J.C.S. Agonistic ethogram of the equid bachelor band. Appl. Anim. Behav. Sci. 1995, 43, 147–188. [Google Scholar] [CrossRef]
- Sigurjónsdóttir, H.; van Dierendonck, M.C.; Snorrason, S.; Thórhallsdóttir, A.G. Social relationships in a group of horses without a mature stallion. Behaviour 2003, 140, 783–804. [Google Scholar] [CrossRef]
- Feh, C. Alliances and reproductive success in Camargue stallions. Anim. Behav. 1999, 57, 705–713. [Google Scholar] [CrossRef]
- Linklater, W.L.; Cameron, E.Z. Tests for cooperative behaviour between stallions. Anim. Behav. 2000, 60, 731–743. [Google Scholar] [CrossRef]
- Feh, C. Relationships and communication in socially. In The Domestic Horse: The Origins, Development and Management of its Behaviour; Mills, D.S., McDonnell, S.M., McDonnell, S., Eds.; Cambridge University Press: Cambridge, UK, 2005; pp. 83–93. [Google Scholar]
- Imanishi, K. Social life of semi-wild horses in Toimisaki. Annu. Anim. Psychol. 1953, 3, 11–31. [Google Scholar] [CrossRef]
- Boyd, L.E.; Carbonaro, D.A.; Houpt, K.A. The 24-hour time budget of Przewalski horses. Appl. Anim. Behav. Sci. 1988, 21, 5–17. [Google Scholar] [CrossRef]
- McDonnell, S.M. A Practical Field Guide to Horse Behavior. The Equid Ethogram; Blood-Horse Publications: Lexington, KY, USA, 2003. [Google Scholar]
- Mooring, M.S.; Blumstein, D.T.; Stoner, C.J. The evolution of parasite-defence grooming in ungulates. Biol. J Linn. Soc. 2004, 81, 17–37. [Google Scholar] [CrossRef]
- Houpt, K.A. Maintenance behaviours. In The Domestic Horse: The Origins, Development and Management of its Behaviour; Mills, D.S., McDonnell, S.M., McDonnell, S., Eds.; Cambridge University Press: Cambridge, UK, 2005; pp. 94–109. [Google Scholar]
- Wells, S.M.; Goldschmidt-Rothschild, B. Social behaviour and relationships in a herd of camargue horses. Zeitschrift für Tierpsychologie 1979, 49, 363–380. [Google Scholar] [CrossRef]
- Briard, L.; Dorn, C.; Petit, O. Personality and Affinities Play a Key Role in the Organisation of Collective Movements in a Group of Domestic Horses. Ethology 2015, 121, 888–902. [Google Scholar] [CrossRef]
- Arnold, G.W.; Grassia, A. Ethogram of agonistic behaviour for thoroughbred horses. Appl. Anim. Ethol. 1982, 8, 5–25. [Google Scholar] [CrossRef]
- Cozzi, A.; Sighieri, C.; Gazzano, A.; Nicol, C.J.; Baragli, P. Post-conflict friendly reunion in a permanent group of horses (Equus caballus). Behav. Process. 2010, 85, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Linklater, W.L.; Cameron, E.Z.; Minot, E.O.; Stafford, K.J. Stallion harassment and the mating system of horses. Anim. Behav. 1999, 58, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Klingel, H. Social organization and reproduction in equids. J. Reprod. Fertil. 1975, 23, 7–11. [Google Scholar]
- Keiper, R.R.; Sambraus, H.H. The stability of equine dominance hierarchies and the effects of kinship, proximity and foaling status on hierarchy rank. Appl. Anim. Behav. Sci. 1986, 16, 121–130. [Google Scholar] [CrossRef]
- Croft, D.P.; James, R.; Krause, J. Exploring Animal Social Networks; Princeton University Press: Princeton, NJ, USA, 2008. [Google Scholar]
- Heitor, F.; Oom, M.d.M.; Vicente, L. Social relationships in a herd of Sorraia horses. Behav. Process. 2006, 73, 170–177. [Google Scholar] [CrossRef]
- Granquist, S.M.; Thorhallsdottir, A.G.; Sigurjonsdottir, H. The effect of stallions on social interactions in domestic and semi feral harems. Appl. Anim. Behav. Sci. 2012, 141, 49–56. [Google Scholar] [CrossRef]
- Kaseda, Y.; Khalil, A.M.; Ogawa, H. Harem stability and reproductive success of Misaki feral mares. Equine Vet. J. 1995, 27, 368–372. [Google Scholar] [CrossRef]
- Kaseda, Y. Seasonal changes in time spent grazing and resting of misaki horses. Nihon Chikusan Gakkaiho 1983, 54, 464–469. [Google Scholar] [CrossRef]
- Kaseda, Y.; Khalil, A.M. Harem size reproductive success of stallions in Misaki feral horses. Appl. Anim. Behav. Sci. 1996, 47, 163–173. [Google Scholar] [CrossRef]
- Kaseda, Y.; Ogawa, H.; Khalil, A.M. Causes of natal dispersal and emigration and their effects on harem formation in Misaki feral horses. Equine Vet. J. 1997, 29, 262–266. [Google Scholar] [CrossRef] [PubMed]
- Martin, P.; Bateson, P. Measuring Behaviour; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Fureix, C.; Bourjade, M.; Henry, S.; Sankey, C.; Hausberger, M. Exploring aggression regulation in managed groups of horses Equus caballus. Appl. Anim. Behav. Sci. 2012, 138, 216–228. [Google Scholar] [CrossRef]
- Jameson, K.A.; Appleby, M.C.; Freeman, L.C. Finding an appropriate order for a hierarchy based on probabilistic dominance. Anim. Behav. 1999, 57, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Hemelrijk, C.K. A matrix partial correlation test used in investigations of reciprocity and other social interaction patterns at group level. J. Theor. Biol. 1990, 143, 405–420. [Google Scholar] [CrossRef]
- Shimada, M.; Sueur, C. Social play among juvenile wild Japanese macaques (Macaca fuscata) strengthens their social bonds. Am. J. Primatol. 2017, 80, e22728. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, H. An introduction to the statistical free software HAD: Suggestions to improve teaching, learning and practice data analysis. J. Media. Inform. Commun. 2016, 1, 59–73. [Google Scholar]
- Borgatti, S.P.; Everett, M.G.; Freeman, L.C. UCINET. In Encyclopedia of Social Network Analysis and Mining; Springer: New York, NY, USA, 2018; pp. 3243–3249. [Google Scholar]
- Inoue, S.; Yamamoto, S.; Ringhofer, M.; Mendonça, R.S.; Pereira, C.; Hirata, S. Spatial positioning of individuals in a group of feral horses: A case study using drone technology. Mammal. Res. 2018, 64, 249–259. [Google Scholar] [CrossRef]
- Gardner, A.; West, S.A. Spite. Curr. Biol. 2006, 16, R662–R664. [Google Scholar] [CrossRef]
- Fedurek, P.; Dunbar, R.I.M. What does mutual grooming tell us about why chimpanzees groom? Ethology 2009, 115, 566–575. [Google Scholar] [CrossRef]
- Matsuzawa, T. Horse cognition and behavior from the perspective of primatology. Primates 2017, 58, 473–477. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| 6m | 16m | 63m | 20f | 73f | 90f | 94f | 75f | 76m | |
|---|---|---|---|---|---|---|---|---|---|
| 6m | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| 16m | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| 63m | 0 | 0.5 | 0 | 0 | 0.25 | 0.25 | |||
| 20f | 0 | 0 | 0 | 0 | 0 | ||||
| 73f | 0 | 0 | 0.5 | 0.5 | |||||
| 90f | 0 | 0 | 0 | ||||||
| 94f | 0 | 0 | |||||||
| 75f | 0.25 | ||||||||
| 76m |
| 6m | 16m | 63m | 20f | 73f | 90f | 94f | 75f | 76m | |
|---|---|---|---|---|---|---|---|---|---|
| 6m | 0 | 0 | 6 | 4 | 1 | 0 | 0 | 1 | |
| 16m | 0 | 0 | 3 | 3 | 0 | 0 | 3 | ||
| 63m | 0 | 0 | 0 | 0 | 0 | 0 | |||
| 20f | 0 | 6 | 0 | 0 | 1 | ||||
| 73f | 0 | 0 | 0 | 2 | |||||
| 90f | 0 | 0 | 4 | ||||||
| 94f | 0 | 0 | |||||||
| 75f | 0 | ||||||||
| 76m |
| 6m | 16m | 63m | 20f | 73f | 90f | 94f | 75f | 76m | |
|---|---|---|---|---|---|---|---|---|---|
| 6m | 0.2383 | 0.0622 | 0.2313 | 0.1775 | 0.1225 | 0.1328 | 0.0687 | 0.0523 | |
| 16m | 0.0359 | 0.0514 | 0.0982 | 0.0741 | 0.1166 | 0.0734 | 0.0619 | ||
| 63m | 0.1274 | 0.2988 | 0.0691 | 0.1294 | 0.1843 | 0.1357 | |||
| 20f | 0.0544 | 0.3393 | 0.1601 | 0.0839 | 0.0660 | ||||
| 73f | 0.0798 | 0.0779 | 0.1141 | 0.1769 | |||||
| 90f | 0.2323 | 0.0705 | 0.0566 | ||||||
| 94f | 0.0967 | 0.0599 | |||||||
| 75f | 0.2889 | ||||||||
| 76m |
| 6m | 16m | 63m | 20f | 73f | 90f | 94f | 75f | 76m | |
|---|---|---|---|---|---|---|---|---|---|
| 6m | 24 | 10 | 13 | 9 | 7 | 14 | 6 | 0 | |
| 16m | 4 | 3 | 1 | 3 | 3 | 2 | 4 | ||
| 63m | 0 | 0 | 5 | 10 | 1 | 2 | |||
| 20f | 2 | 6 | 0 | 0 | 0 | ||||
| 73f | 6 | 3 | 1 | 0 | |||||
| 90f | 12 | 4 | 2 | ||||||
| 94f | 1 | 1 | |||||||
| 75f | 1 | ||||||||
| 76m |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shimada, M.; Suzuki, N. The Contribution of Mutual Grooming to Affiliative Relationships in a Feral Misaki Horse Herd. Animals 2020, 10, 1564. https://doi.org/10.3390/ani10091564
Shimada M, Suzuki N. The Contribution of Mutual Grooming to Affiliative Relationships in a Feral Misaki Horse Herd. Animals. 2020; 10(9):1564. https://doi.org/10.3390/ani10091564
Chicago/Turabian StyleShimada, Masaki, and Nae Suzuki. 2020. "The Contribution of Mutual Grooming to Affiliative Relationships in a Feral Misaki Horse Herd" Animals 10, no. 9: 1564. https://doi.org/10.3390/ani10091564
APA StyleShimada, M., & Suzuki, N. (2020). The Contribution of Mutual Grooming to Affiliative Relationships in a Feral Misaki Horse Herd. Animals, 10(9), 1564. https://doi.org/10.3390/ani10091564