Overfeeding-Induced Obesity Could Cause Potential Immuno-Physiological Disorders in Rainbow Trout (Oncorhynchus mykiss)

 ,
, 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish
2.2. Overfeeding Stress
2.3. Hematology and Serology
2.4. Histological Analysis
2.5. RNA Extraction, cDNA Synthesis, and Real-Time PCR (Quantitative PCR)
2.6. Leukocyte Isolation and OxLDL Treatment
2.7. Annexin-V/PI Double Staining Assay
2.8. Statistical Analysis
3. Results
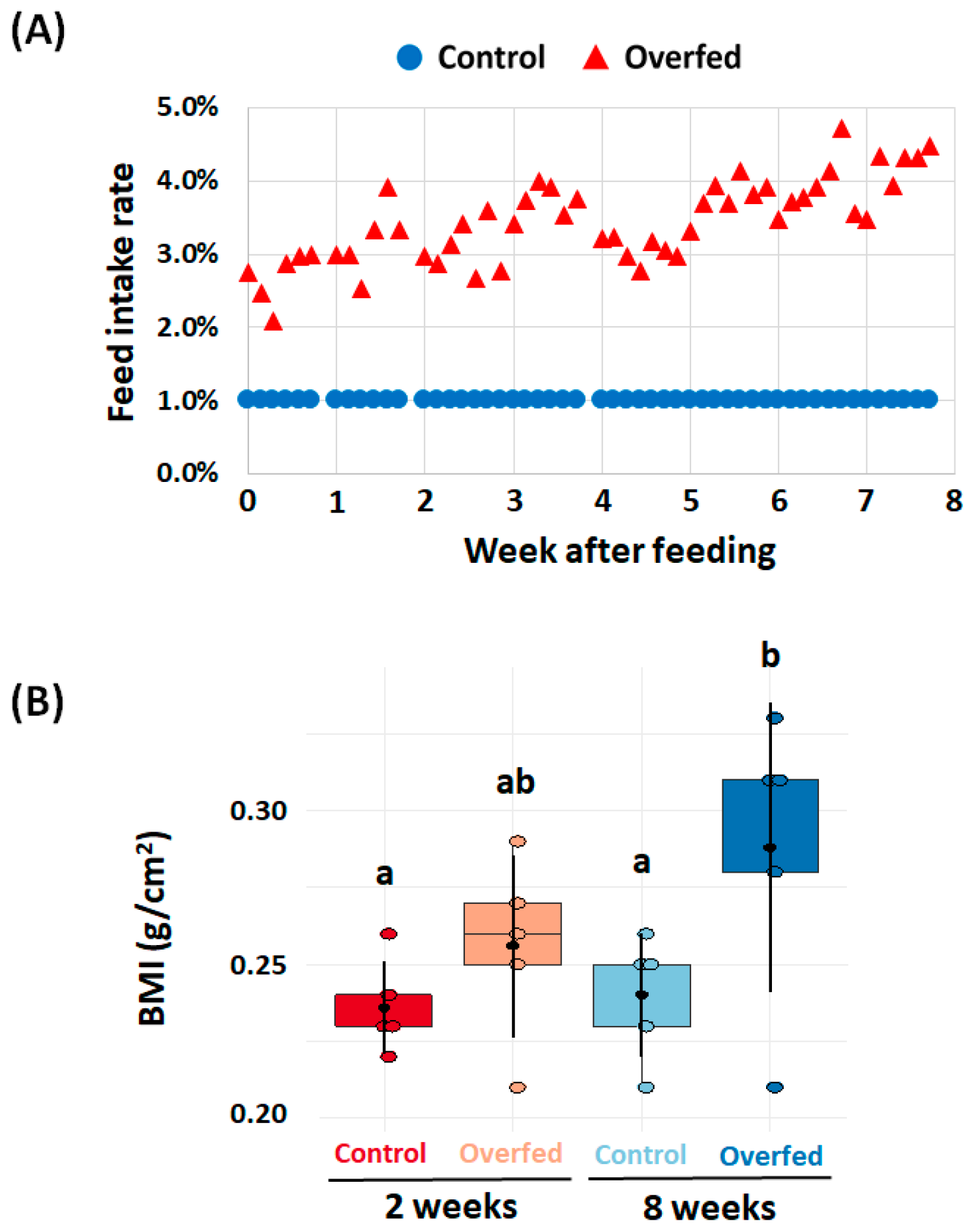
3.1. Fish Feeding Rate and Growth
3.2. Hematology and Hepatosomatic Index (HSI)
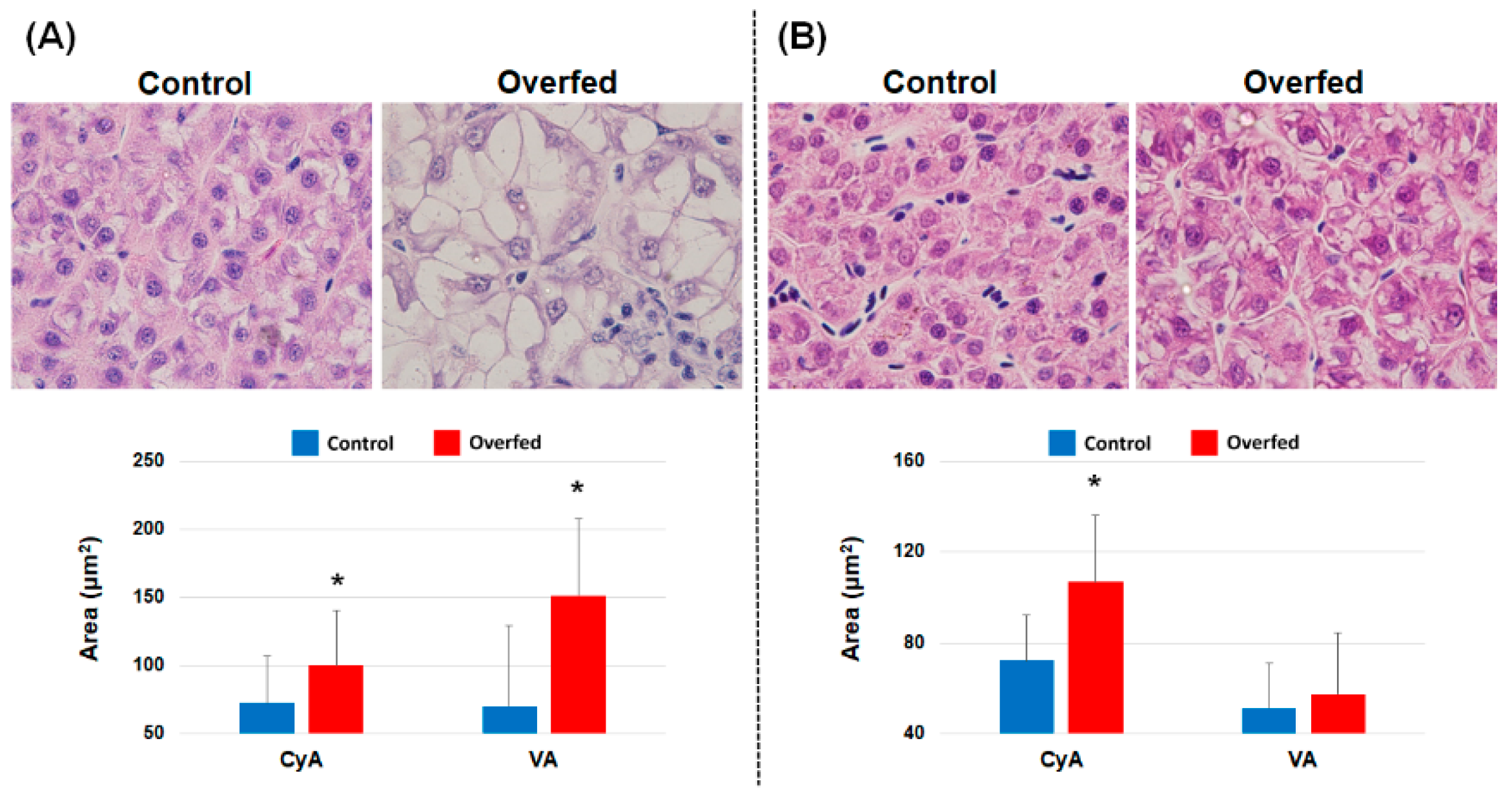
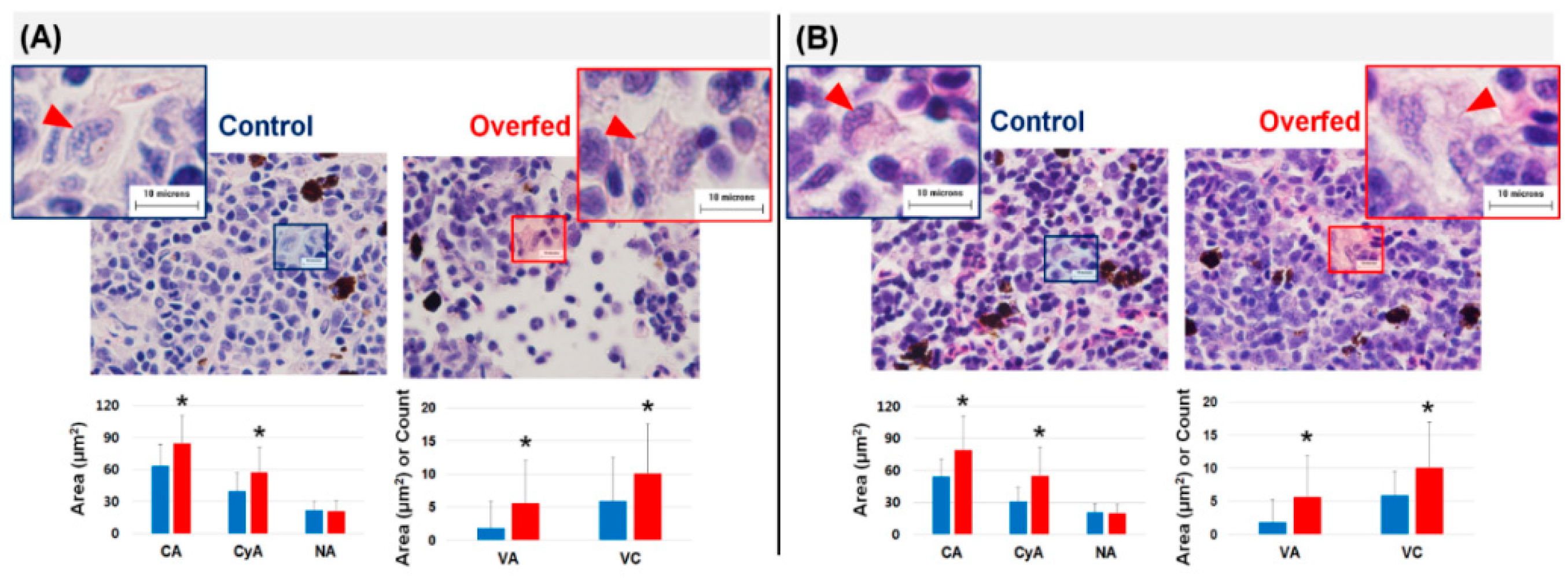
3.3. Histopathology of the Liver and Head Kidney
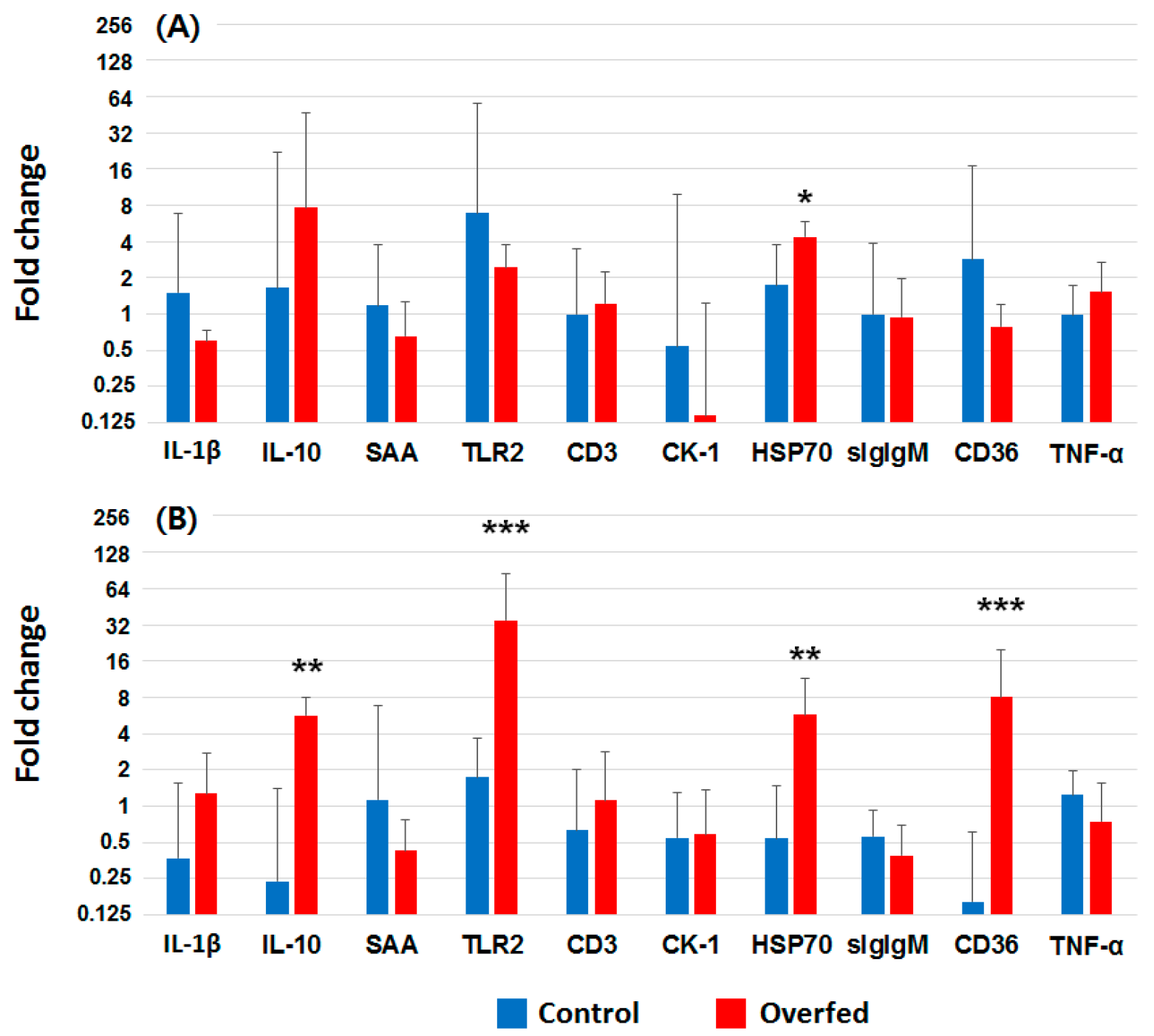
3.4. Gene Expression
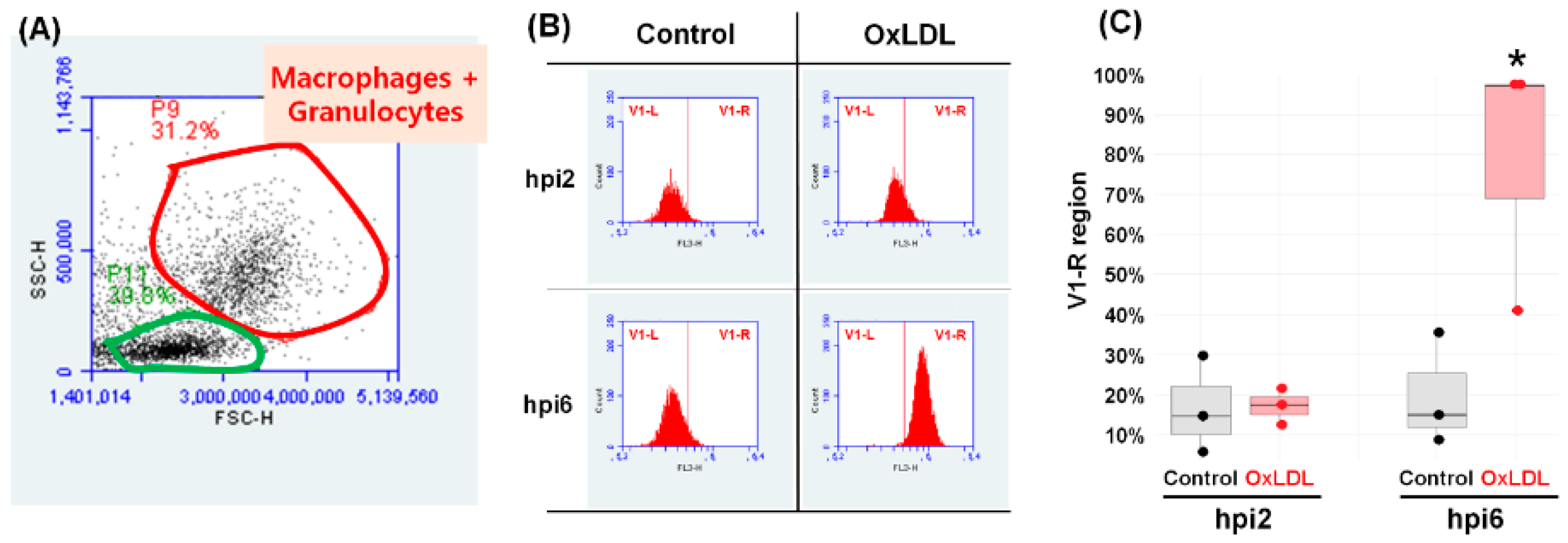
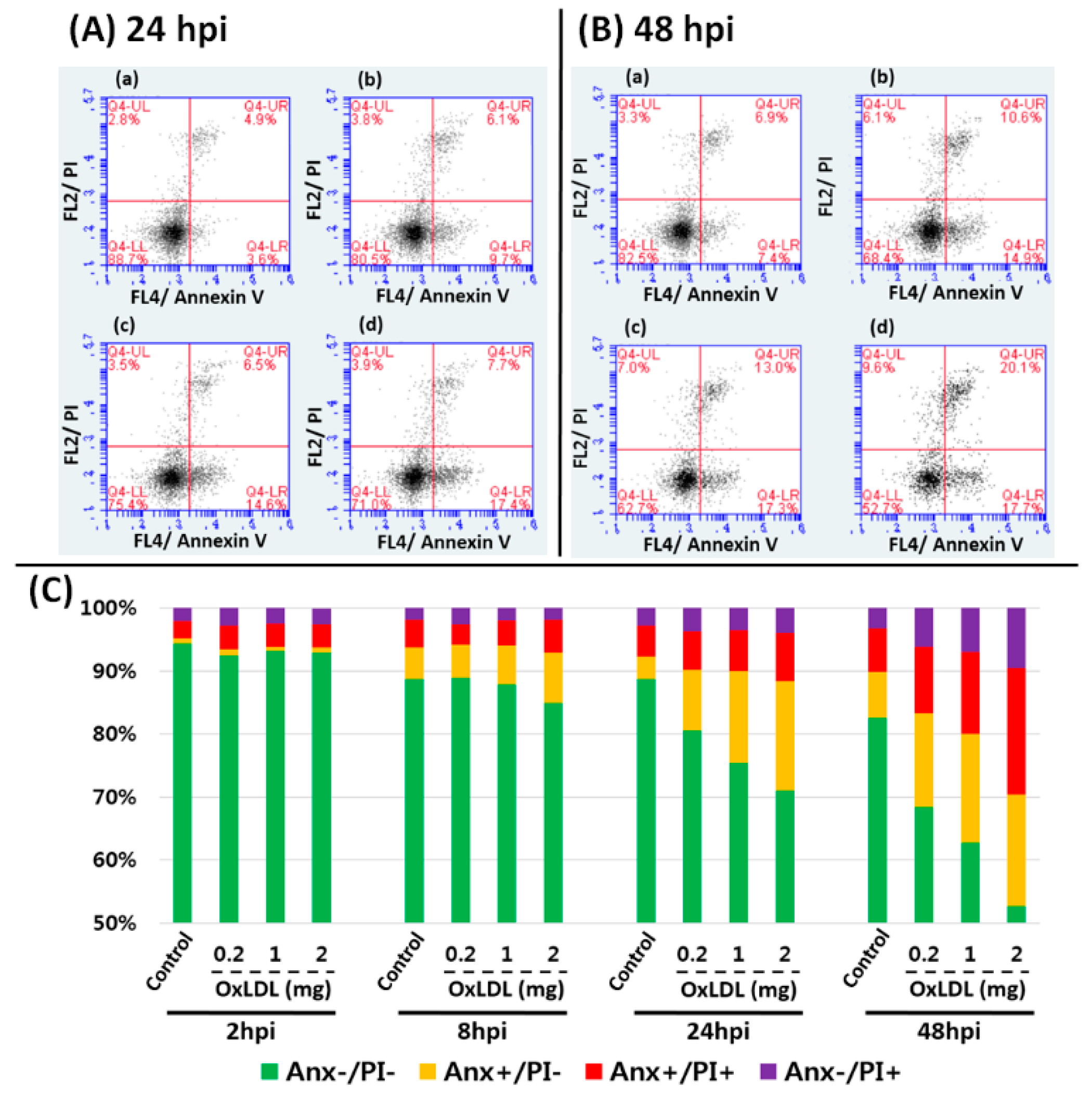
3.5. Flow Cytometry Analysis
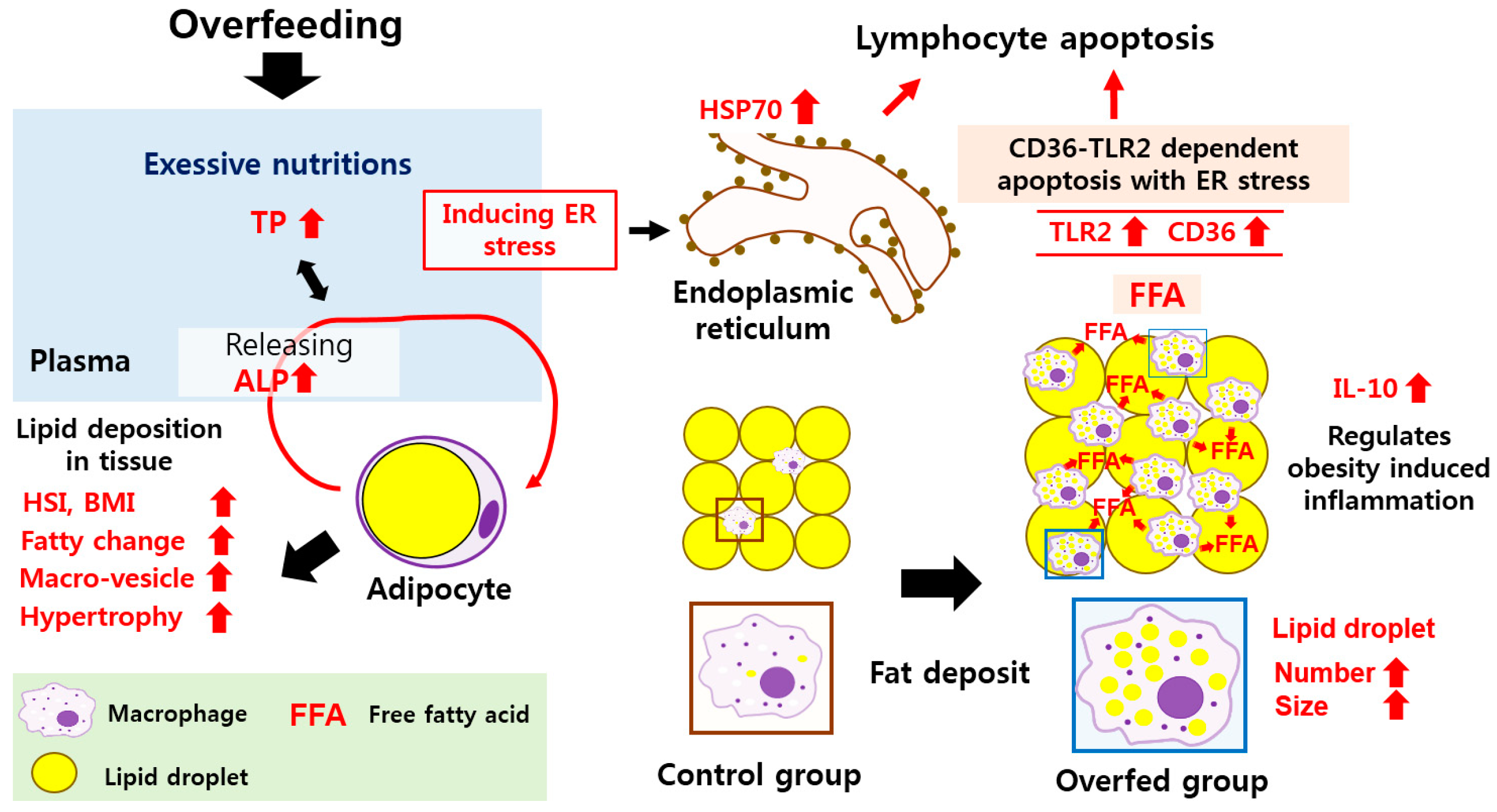
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bureau, D.P.; Hua, K.; Cho, C.Y. Effect of feeding level on growth and nutrient deposition in rainbow trout (Oncorhynchus mykiss Walbaum) growing from 150 to 600 g. Aquac. Res. 2006, 37, 1090–1098. [Google Scholar] [CrossRef]
- Taddese, F.; Huh, M.; Bai, S.C.; Vijverberg, J. Histological changes of liver in overfed young nile tilapia. J. Fish. Aquat. Sci. 2014, 9, 63–74. [Google Scholar] [CrossRef]
- Landgraf, K.; Schuster, S.; Meusel, A.; Garten, A.; Riemer, T.; Schleinitz, D.; Kiess, W.; Körner, A. Short-term overfeeding of zebrafish with normal or high-fat diet as a model for the development of metabolically healthy versus unhealthy obesity. BMC Physiol. 2017, 17, 4. [Google Scholar] [CrossRef] [PubMed]
- Craig, S.; Helfrich, L.A.; Kuhn, D.; Schwarz, M.H. Understanding Fish Nutrition, Feeds, and Feeding; Communications and Marketing, College of Agriculture and Life Sciences, Virginia Tech: Blacksburg, VA, USA, 2017. [Google Scholar]
- White, P. Environmental Consequences of Poor Feed Quality and Feed Management. In On-Farm Feeding and Feed Management in Aquaculture; FAO Fisheries and Aquaculture Technical Paper; FAO: Rome, Italy, 2013. [Google Scholar]
- Talbot, C.; Corneillie, S.; Korsøen, Ø. Pattern of feed intake in four species of fish under commercial farming conditions: Implications for feeding management. Aquac. Res. 1999, 30, 509–518. [Google Scholar] [CrossRef]
- Thorpe, J.; Cho, C.Y. Minimising waste through bioenergetically and behaviourally based feeding strategies. Water Sci. Technol. 1995, 31, 29–40. [Google Scholar] [CrossRef]
- Cho, C.Y.; Bureau, D.P. Development of bioenergetic models and the Fish-PrFEQ software to estimate production, feeding ration and waste output in aquaculture. Aquat. Living Resour. 1998, 11, 199–210. [Google Scholar] [CrossRef]
- Sorof, J.; Daniels, S. Obesity hypertension in children: A problem of epidemic proportions. Hypertension 2002, 40, 441–447. [Google Scholar] [CrossRef]
- Raghavan, S.; Singh, N.K.; Gali, S.; Mani, A.M.; Rao, G.N. Protein kinase Cθ via activating transcription factor 2-mediated CD36 expression and foam cell formation of Ly6Chi cells contributes to atherosclerosis. Circulation 2018, 138, 2395–2412. [Google Scholar] [CrossRef]
- West-Eberhard, M.J. Nutrition, the visceral immune system, and the evolutionary origins of pathogenic obesity. Proc. Natl. Acad. Sci. USA 2019, 116, 723–731. [Google Scholar] [CrossRef]
- Apostolopoulos, V.; de Courten, M.P.; Stojanovska, L.; Blatch, G.L.; Tangalakis, K.; de Courten, B. The complex immunological and inflammatory network of adipose tissue in obesity. Mol. Nutr. Food Res. 2016, 60, 43–57. [Google Scholar] [CrossRef]
- Interator, H.; Lebenthal, Y.; Hoshen, M.; Safra, I.; Balicer, R.; Leshno, M.; Shamir, R. Distinct Lipoprotein Curves in Normal Weight, Overweight, and Obese Children and Adolescents. J. Pediatr. Gastroenterol. Nutr. 2017, 65, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Kassi, E.; Dalamaga, M.; Faviou, E.; Hroussalas, G.; Kazanis, K.; Nounopoulos, C.; Dionyssiou-Asteriou, A. Circulating oxidized LDL levels, current smoking and obesity in postmenopausal women. Atherosclerosis 2009, 205, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.; Ni, L.; Zhuge, F.; Xu, L.; Fu, Z.; Ota, T. Adipose Tissue Macrophage Phenotypes and Characteristics: The Key to Insulin Resistance in Obesity and Metabolic Disorders. Obesity 2020, 28, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Dai, Y.; Liu, M.; Yuan, X.; Wang, C.; Huang, Y.; Liu, W.; Jiang, G. High-fat diet induces aberrant hepatic lipid secretion in blunt snout bream by activating endoplasmic reticulum stress-associated IRE1/XBP1 pathway. Biochim. Biophys. Acta -Mol. Cell Biol. Lipids 2019, 1864, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Pignatelli, J.; Castro, R.; Granja, A.G.; Abós, B.; González, L.; Jensen, L.B.; Tafalla, C. Immunological characterization of the teleost adipose tissue and its modulation in response to viral infection and fat-content in the diet. PLoS ONE 2014, 9, e110920. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, Y.; Hou, C.; Gao, Y.; Wang, Y. Physiological and molecular changes in large yellow croaker (P seudosciaena crocea R.) with high-fat diet-induced fatty liver disease. Aquac. Res. 2015, 46, 272–282. [Google Scholar] [CrossRef]
- Wang, X.; Jiang, W.; Feng, L.; Wu, P.; Liu, Y.; Zeng, Y.; Jiang, J.; Kuang, S.; Tang, L.; Tang, W. Low or excess levels of dietary cholesterol impaired immunity and aggravated inflammation response in young grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2018, 78, 202–221. [Google Scholar] [CrossRef]
- Lauriano, E.; Pergolizzi, S.; Capillo, G.; Kuciel, M.; Alesci, A.; Faggio, C. Immunohistochemical characterization of Toll-like receptor 2 in gut epithelial cells and macrophages of goldfish Carassius auratus fed with a high-cholesterol diet. Fish Shellfish Immunol. 2016, 59, 250–255. [Google Scholar] [CrossRef]
- Roh, H.J.; Kim, A.; Kang, G.S.; Kim, B.S.; Kim, D. Blue light-emitting diode light at 405 and 465 nm can inhibit a Miamiensis avidus infection in olive flounder, Paralichthys olivaceus. Aquaculture 2018, 493, 176–185. [Google Scholar] [CrossRef]
- Castro, R.; Abos, B.; Pignatelli, J.; von Gersdorff Jorgensen, L.; Gonzalez Granja, A.; Buchmann, K.; Tafalla, C. Early immune responses in rainbow trout liver upon viral hemorrhagic septicemia virus (VHSV) infection. PLoS ONE 2014, 9, e111084. [Google Scholar] [CrossRef]
- Kim, D.; Austin, B. Innate immune responses in rainbow trout (Oncorhynchus mykiss, Walbaum) induced by probiotics. Fish Shellfish Immunol. 2006, 21, 513–524. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Konishi, K.; Shima, T.; Furuita, H.; Suzuki, N.; Tabata, M. Influence of dietary fat and carbohydrate levels on growth and body composition of rainbow trout Oncorhynchus mykiss under selffeeding conditions. Fish. Sci. 2001, 67, 221–227. [Google Scholar] [CrossRef]
- FAO. Aquaculture Feed and Fertilizer Resources Information System: Rainbow Trout–Feed Formulation (Website). 2020. Available online: http://www.fao.org/fishery/affris/species-profiles/rainbow-trout/feed-formulation/en/ (accessed on 12 July 2020).
- Qiu, H.; Jin, M.; Li, Y.; Lu, Y.; Hou, Y.; Zhou, Q. Dietary lipid sources influence fatty acid composition in tissue of large yellow croaker (Larmichthys crocea) by regulating triacylglycerol synthesis and catabolism at the transcriptional level. PLoS ONE 2017, 12, e0169985. [Google Scholar] [CrossRef]
- Leitritz, E.; Lewis, R.C. Trout and salmon culture: Hatchery Methods; UCANR Publications: California, CA, USA, 1980. [Google Scholar]
- Forn-Cuni, G.; Varela, M.; Fernandez-Rodriguez, C.M.; Figueras Huerta, A.; Novoa, B. Liver immune responses to inflammatory stimuli in a dietinduced obesity model of zebrafish. J. Endocrinol. 2015, 224, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Montalbano, G.; Mania, M.; Guerrera, M.C.; Abbate, F.; Laurà, R.; Navarra, M.; Vega, J.A.; Ciriaco, E.; Germanà, A. Morphological differences in adipose tissue and changes in BDNF/Trkb expression in brain and gut of a diet induced obese zebrafish model. Ann. Anat.-Anat. Anz. 2016, 204, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Oka, T.; Nishimura, Y.; Zang, L.; Hirano, M.; Shimada, Y.; Wang, Z.; Umemoto, N.; Kuroyanagi, J.; Nishimura, N.; Tanaka, T. Diet-induced obesity in zebrafish shares common pathophysiological pathways with mammalian obesity. BMC Physiol. 2010, 10, 21. [Google Scholar] [CrossRef]
- Melillo, A. Rabbit clinical pathology. J. Exot. Pet Med. 2007, 16, 135–145. [Google Scholar] [CrossRef]
- Mohamed, W.A.; El-Houseiny, W.; Ibrahim, R.E.; Abd-Elhakim, Y.M. Palliative effects of zinc sulfate against the immunosuppressive, hepato-and nephrotoxic impacts of nonylphenol in Nile tilapia (Oreochromis niloticus). Aquaculture 2019, 504, 227–238. [Google Scholar] [CrossRef]
- de Medeiros, A.M.Z.; Côa, F.; Alves, O.L.; Martinez, D.S.T.; Barbieri, E. Metabolic effects in the freshwater fish Geophagus iporangensis in response to single and combined exposure to graphene oxide and trace elements. Chemosphere 2020, 243, 125316. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.T.; Paiker, J.E.; Crowther, N.J. The relationship between anthropometry and serum concentrations of alkaline phosphatase isoenzymes, liver enzymes, albumin, and bilirubin. Am. J. Clin. Pathol. 2006, 126, 437–442. [Google Scholar] [CrossRef]
- Khan, A.R.; Awan, F.R.; Najam, S.; Islam, M.; Siddique, T.; Zain, M. Elevated serum level of human alkaline phosphatase in obesity. Age 2015, 48, 42–45. [Google Scholar]
- Kim, J.; Lee, H.S.; Park, H.; Lee, Y. Serum alkaline phosphatase level is positively associated with metabolic syndrome: A nationwide population-based study. Clin. Chim. Acta 2020, 500, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Rohmah, Z.; Fitria, L.; Kang, S.; Kim, J.; Lee, J. Anti-Obesity effects of Lipid Extract from Sea-Reared of Rainbow Trout (Oncorhynchusmykiss) Fed with Sea Squirt (Halocynthiaroretzi) Tunic’s Carotenoids and CLA. J. Nutr. Food Sci. 2016, 6, 2. [Google Scholar]
- McGoogan, B.B.; Gatlin Iii, D.M. Dietary manipulations affecting growth and nitrogenous waste production of red drum, Sciaenops ocellatus I. Effects of dietary protein and energy levels. Aquaculture 1999, 178, 333–348. [Google Scholar] [CrossRef]
- Kikuchi, K.; Furuta, T.; Iwata, N.; Onuki, K.; Noguchi, T. Effect of dietary lipid levels on the growth, feed utilization, body composition and blood characteristics of tiger puffer Takifugu rubripes. Aquaculture 2009, 298, 111–117. [Google Scholar] [CrossRef]
- Reddy, J.K.; Sambasiva Rao, M. Lipid metabolism and liver inflammation. II. Fatty liver disease and fatty acid oxidation. Am. J. Physiol.-Gastrointest. Liver Physiol. 2006, 290, G852–G858. [Google Scholar] [CrossRef]
- Grundy, S.M. Obesity, metabolic syndrome, and cardiovascular disease. J. Clin. Endocrinol. Metabolism 2004, 89, 2595–2600. [Google Scholar] [CrossRef]
- Schienkiewitz, A.; Schulze, M.B.; Hoffmann, K.; Kroke, A.; Boeing, H. Body mass index history and risk of type 2 diabetes: Results from the European Prospective Investigation into Cancer and Nutrition (EPIC)–Potsdam Study. Am. J. Clin. Nutr. 2006, 84, 427–433. [Google Scholar] [CrossRef]
- Goodman, Z.D. The impact of obesity on liver histology. Clin. Liver Dis. 2014, 18, 33–40. [Google Scholar] [CrossRef]
- Tripathy, D.; Mohanty, P.; Dhindsa, S.; Syed, T.; Ghanim, H.; Aljada, A.; Dandona, P. Elevation of free fatty acids induces inflammation and impairs vascular reactivity in healthy subjects. Diabetes 2003, 52, 2882–2887. [Google Scholar] [CrossRef]
- Ahmad, R.; Al-Mass, A.; Atizado, V.; Al-Hubail, A.; Al-Ghimlas, F.; Al-Arouj, M.; Bennakhi, A.; Dermime, S.; Behbehani, K. Elevated expression of the toll like receptors 2 and 4 in obese individuals: Its significance for obesity-induced inflammation. J. Inflamm. 2012, 9, 48. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, M.T.; Favelyukis, S.; Nguyen, A.K.; Reichart, D.; Scott, P.A.; Jenn, A.; Liu-Bryan, R.; Glass, C.K.; Neels, J.G.; Olefsky, J.M. A subpopulation of macrophages infiltrates hypertrophic adipose tissue and is activated by free fatty acids via Toll-like receptors 2 and 4 and JNK-dependent pathways. J. Biol. Chem. 2007, 282, 35279–35292. [Google Scholar] [CrossRef] [PubMed]
- Senn, J.J. Toll-like receptor-2 is essential for the development of palmitate-induced insulin resistance in myotubes. J. Biol. Chem. 2006, 281, 26865–26875. [Google Scholar] [CrossRef] [PubMed]
- Ehses, J.; Meier, D.; Wueest, S.; Rytka, J.; Boller, S.; Wielinga, P.; Schraenen, A.; Lemaire, K.; Debray, S.; Van Lommel, L. Toll-like receptor 2-deficient mice are protected from insulin resistance and beta cell dysfunction induced by a high-fat diet. Diabetologia 2010, 53, 1795–1806. [Google Scholar] [CrossRef] [PubMed]
- Couper, K.N.; Blount, D.G.; Riley, E.M. IL-10: The master regulator of immunity to infection. J. Immunol. 2008, 180, 5771–5777. [Google Scholar] [CrossRef] [PubMed]
- Glatz, J.F.C.; Luiken, J.J.F.P. Dynamic role of the transmembrane glycoprotein CD36 (SR-B2) in cellular fatty acid uptake and utilization. J. Lipid Res. 2018, 59, 1084–1093. [Google Scholar] [CrossRef]
- Silverstein, R.L.; Febbraio, M. CD36, a scavenger receptor involved in immunity, metabolism, angiogenesis, and behavior. Sci. Signal. 2009, 2, re3. [Google Scholar] [CrossRef]
- Sánchez-Gurmaches, J.; Cruz-Garcia, L.; Gutiérrez, J.; Navarro, I. mRNA expression of fatty acid transporters in rainbow trout: In vivo and in vitro regulation by insulin, fasting and inflammation and infection mediators. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2012, 163, 177–188. [Google Scholar] [CrossRef]
- Torstensen, B.; Nanton, D.; Olsvik, P.; Sundvold, H.; Stubhaug, I. Gene expression of fatty acid-binding proteins, fatty acid transport proteins (cd36 and FATP) and β-oxidation-related genes in Atlantic salmon (Salmo salar L.) fed fish oil or vegetable oil. Aquac. Nutr. 2009, 15, 440–451. [Google Scholar] [CrossRef]
- Fink, I.R.; Benard, E.L.; Hermsen, T.; Meijer, A.H.; Forlenza, M.; Wiegertjes, G.F. Molecular and functional characterization of the scavenger receptor CD36 in zebrafish and common carp. Mol. Immunol. 2015, 63, 381–393. [Google Scholar] [CrossRef]
- Seimon, T.A.; Nadolski, M.J.; Liao, X.; Magallon, J.; Nguyen, M.; Feric, N.T.; Koschinsky, M.L.; Harkewicz, R.; Witztum, J.L.; Tsimikas, S. Atherogenic lipids and lipoproteins trigger CD36-TLR2-dependent apoptosis in macrophages undergoing endoplasmic reticulum stress. Cell Metab. 2010, 12, 467–482. [Google Scholar] [CrossRef]
- Park, S.; Bolender, N.; Eisele, F.; Kostova, Z.; Takeuchi, J.; Coffino, P.; Wolf, D.H. The cytoplasmic Hsp70 chaperone machinery subjects misfolded and endoplasmic reticulum import-incompetent proteins to degradation via the ubiquitin–proteasome system. Mol. Biol. Cell 2007, 18, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Schröder, M. Endoplasmic reticulum stress responses. Cell. Mol. Life Sci. 2008, 65, 862–894. [Google Scholar] [CrossRef] [PubMed]
- Haka, A.S.; Barbosa-Lorenzi, V.C.; Lee, H.J.; Falcone, D.J.; Hudis, C.A.; Dannenberg, A.J.; Maxfield, F.R. Exocytosis of macrophage lysosomes leads to digestion of apoptotic adipocytes and foam cell formation. J. Lipid Res. 2016, 57, 980–992. [Google Scholar] [CrossRef]
- Catrysse, L.; van Loo, G. Adipose tissue macrophages and their polarization in health and obesity. Cell Immunol. 2018, 330, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Fjeldborg, K.; Pedersen, S.B.; Moller, H.J.; Christiansen, T.; Bennetzen, M.; Richelsen, B. Human adipose tissue macrophages are enhanced but changed to an anti-inflammatory profile in obesity. J. Immunol. Res. 2014, 2014, 309548. [Google Scholar] [CrossRef]
- Shapiro, H.; Pecht, T.; Shaco-Levy, R.; Harman-Boehm, I.; Kirshtein, B.; Kuperman, Y.; Chen, A.; Blüher, M.; Shai, I.; Rudich, A. Adipose tissue foam cells are present in human obesity. J. Clin. Endocrinol. Metab. 2013, 98, 1173–1181. [Google Scholar] [CrossRef]
- Tabas, I. Macrophage death and defective inflammation resolution in atherosclerosis. Nat. Rev. Immunol. 2010, 10, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Di Pietro, N.; Formoso, G.; Pandolfi, A. Physiology and pathophysiology of oxLDL uptake by vascular wall cells in atherosclerosis. Vasc. Pharmacol. 2016, 84, 1–7. [Google Scholar] [CrossRef]
- Siklova, M.; Koc, M.; Rossmeislová, L.; Kraml, P. Serum oxLDL–β2GPI complex reflects metabolic syndrome and inflammation in adipose tissue in obese. Int. J. Obes. 2018, 42, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Crowley, L.C.; Marfell, B.J.; Scott, A.P.; Waterhouse, N.J. Quantitation of Apoptosis and Necrosis by Annexin V Binding, Propidium Iodide Uptake, and Flow Cytometry. Cold Spring Harb. Protoc. 2016, 2016. [Google Scholar] [CrossRef] [PubMed]
- Bustamante, M.; Diaz, F.; Munoz, M.; Gross, H.; Rivas, C.; Llancaqueo, A.; Nunez, L.; Campos, L.; Kirsten, L.; Grandón, J. Oxidized low density lipoproteins induce apoptosis in human lymphocytes: Involvement of mitogen-activated protein kinases. Cell Mol. Biol. 2007, 53, OL954–OL964. [Google Scholar] [PubMed]
- Stemme, S.; Faber, B.; Holm, J.; Wiklund, O.; Witztum, J.L.; Hansson, G.K. T lymphocytes from human atherosclerotic plaques recognize oxidized low density lipoprotein. Proc. Natl. Acad. Sci. USA 1995, 92, 3893–3897. [Google Scholar] [CrossRef] [PubMed]
- Ghio, M.; Fabbi, P.; Contini, P.; Fedele, M.; Brunelli, C.; Indiveri, F.; Barsotti, A. OxLDL-and HSP-60 antigen-specific CD8 T lymphocytes are detectable in the peripheral blood of patients suffering from coronary artery disease. Clin. Exp. Med. 2013, 13, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Alcouffe, J.; Therville, N.; Ségui, B.; Nazzal, D.; Blaes, N.; Salvayre, R.; Thomsen, M.; Benoist, H. Expression of membrane-bound and soluble FasL in Fas-and FADD-dependent T lymphocyte apoptosis induced by mildly oxidized LDL. FASEB J. 2004, 18, 122–124. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters + | At Week 2 | At Week 8 | ||
|---|---|---|---|---|
| Control | Overfed | Control | Overfed | |
| HSI (%) | 0.93 ±0.15 | 1.69 ± 0.4 ** | 0.83 ± 0.11 | 1.43 ± 0.57 * |
| RBC (108 cells/mL) | 10.8 ± 3.86 | 12 ± 4.51 | 10.7 ± 1.56 | 10.5 ± 1.94 |
| Hematocrit (%) | 36.6 ± 2.1 | 41.4 ± 4.04 | 33.4 ± 1.8 | 42.6 ± 2.88 *** |
| Hemoglobin (g/dL) | 5.82 ± 1.24 | 7.55 ± 2.37 | 5.71 ± 0.84 | 6.60 ± 1.64 |
| MCV (μm2) | 328 ± 71 | 376 ± 103 | 319 ± 55 | 415 ± 71 |
| MHC (pg) | 55 ± 14 | 64 ± 10 | 54 ± 9 | 65 ± 23 |
| MCHC (%) | 17 ± 3 | 18 ± 5 | 17 ± 2 | 15 ± 3 |
| TP (g/dL) | 4.5 ± 0.4 | 5.3 ± 0.6 * | 4.9 ± 0.4 | 5.9 ± 0.9 * |
| ALP (U/L) | 603 ± 238 | 767 ± 257 | 665 ± 251 | 991 ± 166 * |
| GOT (U/L) | 409.2 ± 152.5 | 364 ± 54 | 440.4 ± 128.3 | 406 ± 73 |
| GPT (U/L) | 14.6 ± 8.6 | 13.80 ± 2.2 | 20.8 ± 9.4 | 15.8 ± 2.1 |
| GLU (mg/dL) | 131.8 ± 15.7 | 118.8 ± 3.4 | 112.2 ± 12.5 | 113.6 ± 20.3 |
| TCHO (mg/dL) | 359.0 ± 37.3 | 394.2 ± 39.0 | 442.8 ± 9.7 | 465.4 ± 50.5 |
| LDH (U/L) | 2268 ± 1200 | 1397 ± 1127 | 820.8 ± 481.4 | 846 ± 139 |
| Ca (mg/dL) | 12.4 ± 0.5 | 11.9 ± 0.65 | 9.4 ± 0.4 | 9.44 ± 0.36 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roh, H.; Park, J.; Kim, A.; Kim, N.; Lee, Y.; Kim, B.S.; Vijayan, J.; Lee, M.K.; Park, C.-I.; Kim, D.-H. Overfeeding-Induced Obesity Could Cause Potential Immuno-Physiological Disorders in Rainbow Trout (Oncorhynchus mykiss). Animals 2020, 10, 1499. https://doi.org/10.3390/ani10091499
Roh H, Park J, Kim A, Kim N, Lee Y, Kim BS, Vijayan J, Lee MK, Park C-I, Kim D-H. Overfeeding-Induced Obesity Could Cause Potential Immuno-Physiological Disorders in Rainbow Trout (Oncorhynchus mykiss). Animals. 2020; 10(9):1499. https://doi.org/10.3390/ani10091499
Chicago/Turabian StyleRoh, HyeongJin, Jiyeon Park, Ahran Kim, Nameun Kim, Yoonhang Lee, Bo Seong Kim, Jasna Vijayan, Mu Kun Lee, Chan-Il Park, and Do-Hyung Kim. 2020. "Overfeeding-Induced Obesity Could Cause Potential Immuno-Physiological Disorders in Rainbow Trout (Oncorhynchus mykiss)" Animals 10, no. 9: 1499. https://doi.org/10.3390/ani10091499
APA StyleRoh, H., Park, J., Kim, A., Kim, N., Lee, Y., Kim, B. S., Vijayan, J., Lee, M. K., Park, C.-I., & Kim, D.-H. (2020). Overfeeding-Induced Obesity Could Cause Potential Immuno-Physiological Disorders in Rainbow Trout (Oncorhynchus mykiss). Animals, 10(9), 1499. https://doi.org/10.3390/ani10091499