Antioxidant Status and Liver Function of Young Turkeys Receiving a Diet with Full-Fat Insect Meal from Hermetia illucens

 , ,
, ,
Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Birds and Housing
2.2. Experimental Design and Diets
2.3. Sample Collection
2.4. Chemical Analyses
2.5. Histological Examination of Tissue Samples
2.6. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Schiavone, A.; Dabbou, S.; de Marco, M.; Cullere, M.; Biasato, I.; Biasibetti, E.; Capucchio, M.T.; Bergagna, S.; Dezzutto, D.; Meneguz, M.; et al. Black soldier fly larva fat inclusion in finisher broiler chicken diet as analternative fat source. Animal 2018, 12, 2032–2039. [Google Scholar] [CrossRef]
- Gariglio, M.; Dabbou, S.; Biasato, I.; Capucchio, M.T.; Colombino, E.; Hernandez, F.; Madrid, S.J.; Martinez, S.; Gai, F.; Caimi, C.; et al. Nutritional effects of the dietary inclusion of partially defatted Hermetia illucens larva meal in Muscovy duck. J. Anim. Sci. Biotechnol. 2019, 10, 37. [Google Scholar] [CrossRef] [PubMed]
- Kierończyk, B.; Sypniewski, J.; Rawski, M.; Czekała, W.; Świątkiewicz, S.; Józefiak, D. From waste to sustainable feed material: The effect of Hermetia illucens oil on the growth performance, nutrient digestibility, and gastrointestinal tract morphometry of broiler chickens. Ann. Anim. Sci. 2020, 20, 157–177. [Google Scholar] [CrossRef]
- Dossey, A.T.; Morales-Ramos, J.A.; Rojas, M.G. Insects as Sustainable Food Ingredients: Production, Processing and Food Applications; Academic Press: London, UK, 2016. [Google Scholar]
- Bulak, P.; Proc, K.; Pawłowska, M.; Kasprzycka, A.; Berus, W.; Bieganowski, A. Biogas generation from insects breeding post production wastes. J. Clean Prod. 2020, 244, 118777. [Google Scholar] [CrossRef]
- Biasato, I.; de Marco, M.; Rotolo, L.; Renna, M.; Dabbou, S.; Capucchio, M.T.; Biasibetti, E.; Tarantola, M.; Costa, P.; Gai, F.; et al. Effects of dietary Tenebrio molitor meal inclusion in free-Range chickens. J. Anim. Physiol. Anim. Nutr. 2016, 100, 1104–1112. [Google Scholar] [CrossRef]
- Loponte, R.; Nizza, S.; Bovera, F.; de Riu, N.; Fliegerova, K.; Lombardi, P.; Vassalotti, G.; Mastellone, V.; Nizza, A.; Moniello, G. Growth performance, blood profiles and carcass traits of barbary partridge (Alectoris barbara) fed two different insect larvae meals (Tenebrio molitor and Hermetia illucens). Res. Vet. Sci. 2017, 115, 183–188. [Google Scholar] [CrossRef]
- Cullere, M.; Tasoniero, G.; Giaccone, V.; Miotti, S.R.; Claeys, R.; de Smet, S.; Dalle, Z.A. Black soldier fly as dietary protein source for broiler quails: Apparent digestibility, excreta microbial load, feed choice, performance, carcass and meat traits. Animal 2016, 10, 1923–1930. [Google Scholar] [CrossRef]
- Dabbou, S.; Gai, F.; Biasato, I.; Capucchio, M.T.; Biasibetti, E.; Dezzutto, D.; Meneguz, M.; Plachà, I.; Gasco, L.; Schiavone, A. Black soldier fly defatted meal as a dietary protein source for broiler chickens: Effects on growth performance, blood traits gut morphology and histological features. J. Anim. Sci. Biotechnol. 2018, 9, 49. [Google Scholar] [CrossRef]
- Barragan-Fonseca, K.B.; Dicke, M.; van Loon, J.J.A. Nutritional value of the black soldier fly (Hermetia illucens L.) and its suitability as animal feed—A review. J. Insects Food Feed 2017, 3, 105–120. [Google Scholar] [CrossRef]
- Dutta, P.K.; Dutta, J.; Tripathi, V.S. Chitin and chitosan: Chemistry, properties and applications. J. Sci. Ind. Res. 2004, 63, 20–31. [Google Scholar]
- Józefiak, A.; Engberg, R.M. Insect proteins as a potential source of antimicrobial peptides in livestock production. A review. J. Anim. Feed Sci. 2017, 26, 87–99. [Google Scholar] [CrossRef]
- Boman, H.G. Peptide antibiotics and their role in innate immunity. Ann. Rev. Immunol. 1995, 13, 61–92. [Google Scholar] [CrossRef] [PubMed]
- Imamura, M.; Wada, S.; Koizumi, N.; Kadotani, T.; Yaoi, K.; Sato, R.; Iwahana, H. Acaloleptins A: Inducible antibacterial peptides from larvae of the beetle, Acalolepta luxuriosa. Arch. Insect Biochem. Physiol. 1999, 40, 88–98. [Google Scholar] [CrossRef]
- Wu, Q.; Patocka, J.; Kuca, K. Insect antimicrobial peptides, a mini review. Toxins 2018, 10, 461. [Google Scholar] [CrossRef]
- Secci, G.; Bovera, F.; Nizza, S.; Baronti, N.; Gasco, L.; Conte, G.; Serra, A.; Bonelli, A.; Parisi, G. Quality of eggs from Lohmann Brown Classic laying hens fed black soldier fly meal as substitute for soya bean. Animal 2018, 12, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Ngo, D.H.; Kim, S.K. Two Antioxidant effects of chitin, chitosan, and their derivatives. In Advances in Food and Nutrition Research; Kim, S.K., Ed.; Academic Press: Waltham, MA, USA, 2014; Volume 73, pp. 15–31. [Google Scholar]
- Marono, S.; Loponte, R.; Lombardi, P.; Vassalotti, G.; Pero, M.E.; Russo, F.; Gasco, L.; Parisi, G.; Piccolo, G.; Nizza, S.; et al. Productive performance and blood profiles of laying hens fed Hermetia illucens larvae meal as total replacement of soybean meal from 24 to 45 weeks of age. Poult. Sci. 2017, 96, 1783–1790. [Google Scholar] [CrossRef] [PubMed]
- Gasco, L.; Finke, M.; van Huis, A. Can diets containing insects promote animal health? J. Insects Food Feed 2018, 4, 1–4. [Google Scholar] [CrossRef]
- Biasato, I.; Gasco, L.; de Marco, M.; Renna, M.; Rotolo, L.; Dabbou, S.; Capucchio, M.T.; Biasibetti, E.; Tarantola, M.; Sterpone, L.; et al. Yellow mealworm larva (Tenebrio molitor) inclusion in diets for male broiler chickens: Effects on growth performance, gut morphology, and histological findings. Poult. Sci. 2018, 97, 540–548. [Google Scholar] [CrossRef]
- Józefiak, A.; Kierończyk, B.; Rawski, M.; Mazurkiewicz, J.; Benzertiha, A.; Gobbi, P.; Nogales-Mérida, S.; Świątkiewicz, S.; Józefiak, D. Full-Fat insect meals as feed additive—The effect on broiler chicken growth performance and gastrointestinal tract microbiota. J. Anim. Feed Sci. 2018, 27, 131–139. [Google Scholar] [CrossRef]
- British United Turkeys (BUT): Aviagen Turkeys 2013. Management Guidelines for Raising Commercial Turkeys. Available online: https://www.aviagenturkeys.com/media/183481/aviagencommercial guide.pdf (accessed on 20 October 2013).
- Smulikowska, S.; Rutkowski, A. (Eds.) Recommended Allowances and Nutritive Value of Feedstuffs. Poultry Feeding Standards (in Polish), 4th ed.; The Kielanowski Institute of Animal Physiology and Nutrition, PAS: Jabłonna, Poland, 2005. [Google Scholar]
- Oluokun, J.A. Upgrading the nutritive value of full-Fat soya beans meal for broiler production with either fishmeal or black soldier fly larvae meal (Hermetia illucens). Niger. J. Anim. Sci. 2000, 3. [Google Scholar] [CrossRef]
- Khan, S.; Khan, R.U.; Sultan, A.; Khan, M.; Hayat, S.U.; Shahid, M.S. Evaluating the suitability of maggot meal as a partial substitute of soya bean on the productive traits, digestibility indices and organoleptic properties of broiler meat. J. Anim. Physiol. Anim. Nutr. 2016, 100, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Attivi, K.; Agboka, K.; Mlaga, G.K.; Oke, O.E.; Teteh, A.; Onagbesan, O.; Tona, K. Effect of black soldier fly (Hermetia Illucens) maggots meal as a substitute for fish meal on growth performance, biochemical parameters and digestibility of broiler chickens. Int. J. Poult. Sci. 2020, 19, 75–80. [Google Scholar] [CrossRef]
- Li, X.; Zhang, D.; Yang, T.Y.; Bryden, W.L. Phosphorus bioavailability: A key aspect for conserving this critical animal feed resource with reference to broiler nutrition. Agriculture 2016, 6, 25. [Google Scholar] [CrossRef]
- Newton, L.; Sheppard, C.; Watson, W.; Burtle, G.; Dove, R. Using the Black Soldier Fly, Hermetia illucens, as a Value-Added Tool for the Management of Swine Manure; Animal and Poultry Waste Management Center, North Carolina State University: Raleigh, NC, USA, 2005; pp. 1–17. [Google Scholar]
- Oonincx, D.G.A.B.; van Broekhoven, S.; van Huis, A.; van Loon, J.J.A. Feed conversion, survival and development, and composition of four insect species on diets composed of food by-products. PLoS ONE 2015, 10, e0144601. [Google Scholar] [CrossRef]
- Dierenfeld, E.S.; King, J. Digestibility and mineral availability of phoenix worms, hermetia illucens, ingested by mountain chicken frogs, Leptodactylus fallax. J. Herpetol. Med. Surg. 2008, 18, 100–105. [Google Scholar] [CrossRef]
- Bovera, F.; Piccolo, G.; Gasco, L.; Marono, S.; Loponte, R.; Vassalotti, G.; Mastellone, V.; Lombardi, P.; Attia, Y.A.; Nizza, A. Yellow mealworm larvae (Tenebrio molitor, L.) as a possible alternative to soybean meal in broiler diets. Br. Poult. Sci. 2015, 56, 569–575. [Google Scholar] [CrossRef]
- Hossain, S.M.; Blair, R. Chitin utilisation by broilers and its effect on body composition and blood metabolites. Br. Poult. Sci. 2007, 48, 33–38. [Google Scholar] [CrossRef]
- Prajapati, B.G.; Patel, R.P. Nutrition, dietary supplements and herbal medicines: A safest approach for obesity. Res. J. Pharmac. Biol. Chem. Sci. 2010, 1, 39–45. [Google Scholar]
- Schiavone, A.; Cullere, M.; de Marco, M.; Meneguz, M.; Biasato, I.; Bergagna, S.; Dezzutto, D.; Gai, F.; Dabbou, S.; Gasco, L.; et al. Partial or total replacement of soybean oil by black soldier fly larvae (Hermetia illucens L.) fat in broiler diets: Effect on growth performances, feed choice, blood traits, carcass characteristics and meat quality. Italian J. Anim. Sci. 2017, 16, 93–100. [Google Scholar] [CrossRef]
- Sypniewski, J.; Kierończyk, B.; Benzertiha, A.; Mikołajczak, Z.; Pruszyńska-Oszmałek, E.; Kołodziejski, P.; Sassek, M.; Rawski, M.; Czekała, W.; Józefiak, D. Replacement of soybean oil by Hermetia illucens fat in turkey nutrition: Effect on performance, digestibility, microbial community, immune and physiological status and final product quality. Br. Poult. Sci. 2020, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Tran, G.; Heuzé, V.; Makkar, H. Insects in fish diets. Anim. Front. 2015, 5, 37–44. [Google Scholar]
- Ushakova, N.A.; Brodskii, E.S.; Kovalenko, A.A.; Bastrakov, A.I.; Kozlova, A.A.; Pavlov, A.D.S. Characteristics of lipid fractions of larvae of the black soldier fly Hermetia illucens. Dokl. Biochem. Biophys. 2016, 468, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Klasing, K.C. Comparative Avian Nutrition; CAB International: Wallingford, UK, 2000. [Google Scholar]
- Biasato, I.; Gasco, L.; de Marco, M.; Renna, M.; Rotolo, L.; Dabbou, S.; Capucchio, M.T.; Biasibetti, E.; Tarantola, M.; Bianchi, C.; et al. Effects of yellow mealworm larva (Tenebrio molitor) inclusion in diets for female broiler chickens: Implications for animal health and gut histology. Anim. Feed Sci. Technol. 2017, 234, 253–263. [Google Scholar] [CrossRef]
- Vargas-Abúndez, J.; Basilio, R.A.; Foddai, M.; Sanchini, L.; Truzzi, C.; Giorgini, E.; Gasco, L.; Olivotto, I. Insect meal based diets for clownfish: Biometric, histological, spectroscopic, biochemical and molecular implications. Aqua 2019, 498, 1–11. [Google Scholar] [CrossRef]
- Chartrin, P.; Bernadet, M.D.; Sannier, M.; Baéza, E. Is ability to hepatic steatosis influenced by age at the beginning of the overfeeding period in Muscovy and Pekin ducks? Animal 2013, 7, 682–687. [Google Scholar] [CrossRef]
- Hérault, F.; Duby, C.; Baéza, E.; Diot, C. Adipogenic genes expression in relation to hepatic steatosis in the liver of two ducks species. Animal 2018, 3, 1–7. [Google Scholar] [CrossRef]
- Hyder, M.A.; Hasan, M.; Mohieldein, A.H. Comparative levels of ALT, AST, ALP and GGT in liver associated diseases. Europ. J. Exp. Biol. 2013, 3, 280–284. [Google Scholar]
- Chaklader, M.R.; Muhammad, A.; Siddik, B.; Fotedar, R.; Howieson, J. Insect larvae, Hermetia illucens in poultry by-product meal for barramundi, Lates calcarifer modulates histomorphology, immunity and resistance to Vibrio harveyi. Sci. Rep. 2019, 9, 16703. [Google Scholar] [CrossRef]
- Elia, A.C.; Capucchio, M.T.; Caldaronia, B.; Magara, G.; Josef, A.; Dörra, M.; Biasato, I.; Biasibetti, E.; Righetti, M.; Pastorino, P.; et al. Influence of Hermetia illucens meal dietary inclusion on the histological traits, gut mucin composition and the oxidative stress biomarkers in rainbow trout (Oncorhynchus mykiss). Aquaculture 2018, 496, 50–57. [Google Scholar] [CrossRef]
 foci of fatty degeneration;
foci of fatty degeneration;  blood stasis in the central vein;
blood stasis in the central vein;  local infiltration of mononuclear cells
foci of fatty degeneration; blood stasis in the central vein; local infiltration of mononuclear cells
local infiltration of mononuclear cells
foci of fatty degeneration; blood stasis in the central vein; local infiltration of mononuclear cells

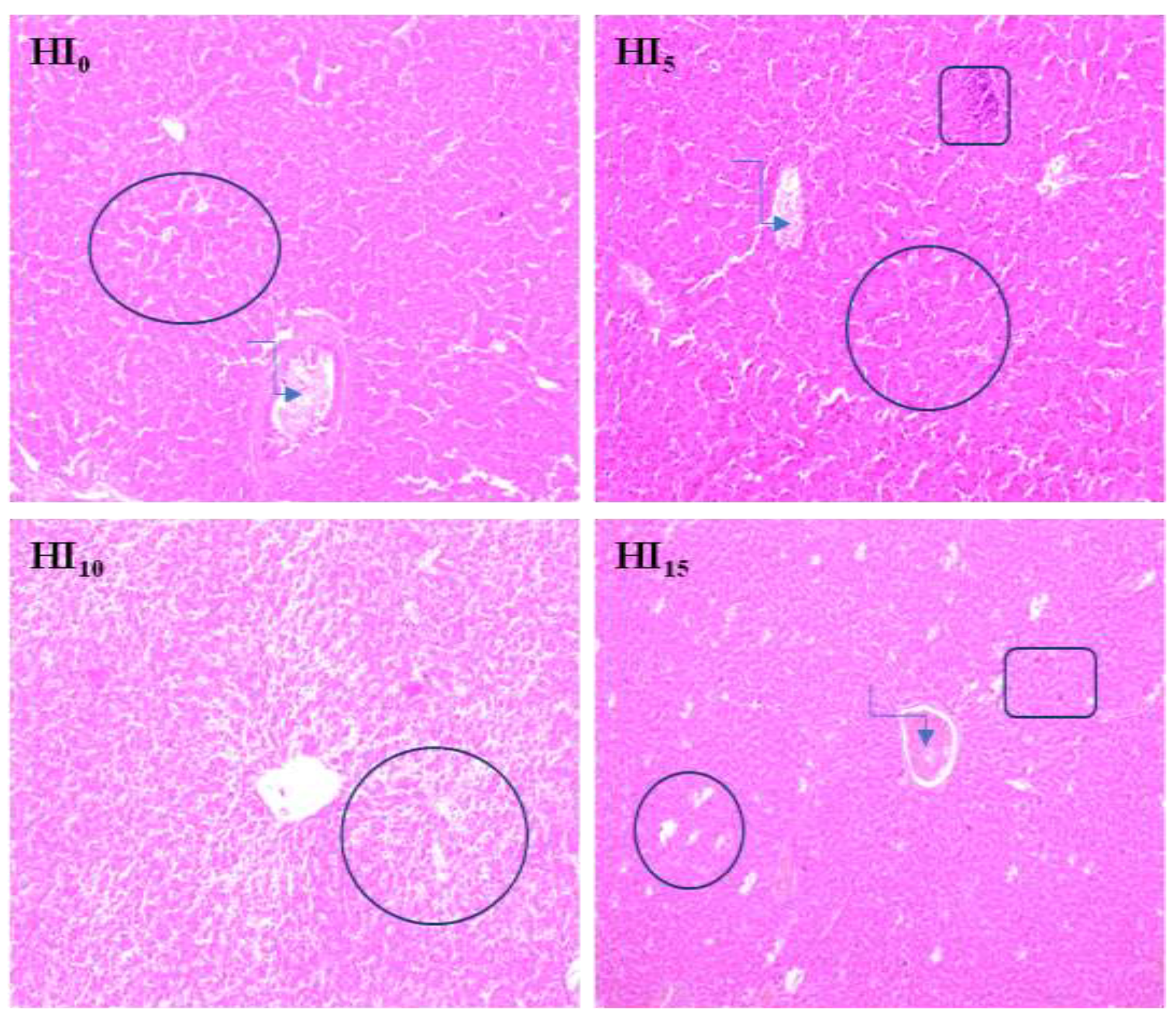{kind=link}
| Item | HI |
|---|---|
| Dry matter, g/100 g | 97.50 |
| Crude protein, g/100 g | 40.4 |
| Ether extract, g/100 g | 33.5 |
| Calcium, g/100 g | 1.36 |
| Phosphorus, g/100 g | 0.79 |
| Zinc, mg/kg | 146 |
| Copper, mg/kg | 11 |
| Chitin, g/100 g | 8.0 |
| Fatty acid profile, g/100 g | |
| C8:0 Caprylic acid | 0.15 |
| C12:0 Lauric acid | 21.0 |
| C14:0 Myristic acid | 2.65 |
| C14:1 Miristoleic acid | 0.08 |
| C16:0 Palmitic acid | 3.54 |
| C16:1n7 Palmitoleic acid | 1.05 |
| C18:0 Stearic acid | 0.45 |
| C18:1n0 Oleic acid | 2.71 |
| C18:2n6 Linoleic acid | 1.73 |
| C18:3n3 α-Linoleic acid | 0.15 |
| C20:0 Arachidic acid | 0.08 |
| C20:4n-6 Arachidonic acid | 0.03 |
| C20:5n-3 Eicosapentaenoic acid | 0.03 |
| C22:6n-3 Docosapentaenoic acid | 0.03 |
| SAFA | 18.73 |
| MUFA | 3.84 |
| PUFA | 1.97 |
| Total Omega-3 | 0.15 |
| Diet 1 | ||||
|---|---|---|---|---|
| HI0 | HI5 | HI10 | HI15 | |
| Component | ||||
| Wheat | 43.51 | 44.80 | 46.09 | 47.41 |
| Soybean meal | 42.94 | 38.47 | 33.99 | 29.50 |
| Black soldier fly meal | - | 5.00 | 10.00 | 15.00 |
| Rapeseed meal | 3.00 | 3.00 | 3.00 | 3.00 |
| Soybean oil | 4.88 | 3.27 | 1.66 | 0.05 |
| Salt | 0.30 | 0.30 | 0.29 | 0.28 |
| Limestone | 1.79 | 1.71 | 1.64 | 1.56 |
| MCP | 2.29 | 2.14 | 2.00 | 1.86 |
| Choline chloride | 0.10 | 0.10 | 0.10 | 0.10 |
| DL-Methionine | 0.33 | 0.34 | 0.35 | 0.36 |
| L-Lysine | 0.49 | 0.50 | 0.51 | 0.51 |
| L-Threonine | 0.12 | 0.12 | 0.12 | 0.12 |
| Vitamins + trace minerals 2 | 0.25 | 0.25 | 0.25 | 0.25 |
| Calculated nutrient density 3 | ||||
| ME (kcal/kg) | 2850 | 2850 | 2850 | 2850 |
| Crude protein | 27.50 | 27.50 | 27.50 | 27.50 |
| Crude fibre | 3.11 | 3.16 | 3.21 | 3.26 |
| Lysine | 1.80 | 1.80 | 1.80 | 1.80 |
| Methionine | 0.71 | 0.72 | 0.73 | 0.75 |
| Met. + Cys. | 1.17 | 1.17 | 1.17 | 1.17 |
| Threonine | 1.10 | 1.10 | 1.10 | 1.10 |
| Calcium | 1.30 | 1.30 | 1.30 | 1.30 |
| Available phosphorus | 0.70 | 0.70 | 0.70 | 0.70 |
| Analysed nutrients | ||||
| Crude protein | 28.26 | 27.92 | 27.14 | 28.47 |
| Crude fat | 6.84 | 6.17 | 5.63 | 5.29 |
| Dry matter | 90.54 | 90.33 | 90.08 | 90.51 |
| Crude ash | 7.35 | 7.24 | 7.22 | 7.39 |
| MJ ME/kg | 17.68 | 17.70 | 17.54 | 17.66 |
| Initial BW kg | Final BW kg | FCR kg/kg | Liveability % | |
|---|---|---|---|---|
| Group 1 | ||||
| HI0 | 0.059 | 1.104 | 1.642 b | 99.07 |
| HI5 | 0.059 | 1.121 | 1.604 ab | 99.07 |
| HI10 | 0.059 | 1.129 | 1.609 ab | 99.07 |
| HI15 | 0.059 | 1.149 | 1.577 a | 99.07 |
| SEM | 0.0001 | 0.008 | 0.008 | - |
| p-value | ||||
| Group | - | 0.074 | 0.042 | - |
| Linear | - | 0.056 | 0.011 | - |
| Quadratic | - | 0.936 | 0.877 | - |
| Cubic | - | 0.775 | 0.272 | - |
| Hb g/L | GLU mmol/L | TP g/L | TC mmol/L | MDA µmol/mL | AST U/L | SOD U/mL | |
|---|---|---|---|---|---|---|---|
| Group 1 | |||||||
| HI0 | 24.22 b | 17.63 c | 22.31 ab | 2.137 b | 1.033 | 159.8 c | 36.75 a |
| HI5 | 26.21 ab | 20.31 bc | 23.65 a | 2.145 b | 1.034 | 158.4 c | 36.65 ab |
| HI10 | 27.96 a | 22.11 ab | 21.88 b | 3.443 a | 0.999 | 185.3 b | 34.92 ab |
| HI15 | 27.47 a | 23.74 a | 21.91 b | 3.038 a | 0.986 | 223.7 a | 33.71 b |
| SEM total | 0.366 | 0.482 | 0.231 | 0.102 | 0.016 | 4.478 | 0.426 |
| p-Value: | |||||||
| Group | 0.001 | <0.001 | 0.018 | <0.001 | 0.633 | <0.001 | 0.027 |
| Linear | 0.001 | <0.001 | 0.135 | <0.001 | 0.223 | <0.001 | 0.004 |
| Quadratic | 0.061 | 0.517 | 0.138 | 0.169 | 0.829 | 0.002 | 0.497 |
| Cubic | 0.496 | 0.848 | 0.014 | <0.001 | 0.691 | 0.539 | 0.533 |
| Ca mmol/L | P mmol/L | Mg mmol/L | Fe µmol/L | Cu µmol/L | Zn µmol/L | |
|---|---|---|---|---|---|---|
| Group 1 | ||||||
| HI0 | 2.957 | 1.400 b | 3.196 | 11.53 b | 50.26 | 58.08 b |
| HI5 | 3.082 | 1.466 ab | 3.119 | 11.12 b | 50.70 | 64.05 b |
| HI10 | 2.999 | 1.464 ab | 3.350 | 14.93 a | 51.41 | 75.56 a |
| HI15 | 3.007 | 1.535 a | 2.676 | 15.67 a | 53.05 | 75.27 a |
| SEM total | 0.028 | 0.014 | 0.101 | 0.469 | 0.902 | 1.722 |
| p-Value: | ||||||
| Group | 0.475 | 0.006 | 0.102 | <0.001 | 0.720 | <0.001 |
| Linear | 0.792 | 0.001 | 0.136 | <0.001 | 0.273 | <0.001 |
| Quadratic | 0.303 | 0.919 | 0.134 | 0.480 | 0.744 | 0.294 |
| Cubic | 0.244 | 0.233 | 0.173 | 0.048 | 0.934 | 0.194 |
| MDA µmol/kg | GSH + GSSG µmol/kg | CAT U/g Protein | SOD U/g Protein | |
|---|---|---|---|---|
| Group 1 | ||||
| HI0 | 1.107 y | 53.56 a | 11.92 ab | 6.198 x |
| HI5 | 1.339 xy | 50.71 a | 10.60 b | 5.732 xy |
| HI10 | 1.631 x | 49.96 a | 13.95 a | 4.785 y |
| HI15 | 1.367 xy | 41.73 b | 13.23 ab | 5.642 xy |
| SEM total | 0.071 | 1.200 | 0.461 | 0.186 |
| p-Value: | ||||
| Group | 0.072 | 0.002 | 0.049 | 0.054 |
| Linear | 0.085 | <0.001 | 0.072 | 0.108 |
| Quadratic | 0.075 | 0.222 | 0.739 | 0.070 |
| Cubic | 0.317 | 0.331 | 0.031 | 0.159 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ognik, K.; Kozłowski, K.; Stępniowska, A.; Listos, P.; Józefiak, D.; Zduńczyk, Z.; Jankowski, J. Antioxidant Status and Liver Function of Young Turkeys Receiving a Diet with Full-Fat Insect Meal from Hermetia illucens. Animals 2020, 10, 1339. https://doi.org/10.3390/ani10081339
Ognik K, Kozłowski K, Stępniowska A, Listos P, Józefiak D, Zduńczyk Z, Jankowski J. Antioxidant Status and Liver Function of Young Turkeys Receiving a Diet with Full-Fat Insect Meal from Hermetia illucens. Animals. 2020; 10(8):1339. https://doi.org/10.3390/ani10081339
Chicago/Turabian StyleOgnik, Katarzyna, Krzysztof Kozłowski, Anna Stępniowska, Piotr Listos, Damian Józefiak, Zenon Zduńczyk, and Jan Jankowski. 2020. "Antioxidant Status and Liver Function of Young Turkeys Receiving a Diet with Full-Fat Insect Meal from Hermetia illucens" Animals 10, no. 8: 1339. https://doi.org/10.3390/ani10081339
APA StyleOgnik, K., Kozłowski, K., Stępniowska, A., Listos, P., Józefiak, D., Zduńczyk, Z., & Jankowski, J. (2020). Antioxidant Status and Liver Function of Young Turkeys Receiving a Diet with Full-Fat Insect Meal from Hermetia illucens. Animals, 10(8), 1339. https://doi.org/10.3390/ani10081339