Impact of the Mid-Pleistocene Revolution and Anthropogenic Factors on the Dispersion of Asian Black-Spined Toads (Duttaphrynus melanostictus)

 ,
,  , ,
, , 
Abstract
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Taxa Sampling and DNA Extraction
2.2. PCR Amplification and DNA Sequencing
2.3. Haplotype Network and Phylogenetic Analyses
2.4. Molecular Dating
2.5. Estimating Past Population Dynamics
2.6. Genetic Admixtures Analyses
2.7. Isolation by Distance and Population Structure Analyses
3. Results
3.1. Haplotype Network
3.2. Phylogenetic Reconstruction Based on mtDNA Sequence
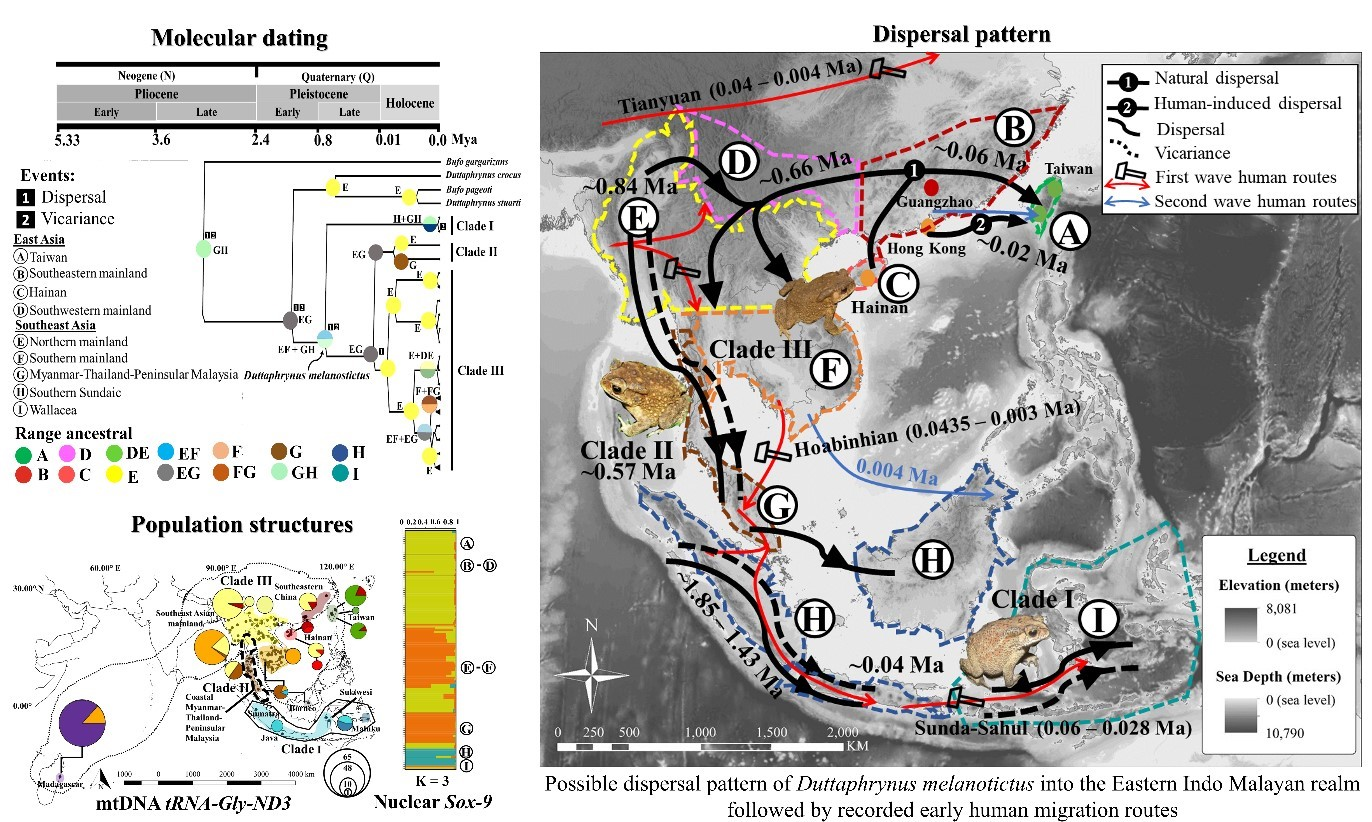
3.3. Molecular Dating and Ancestral Range Reconstruction
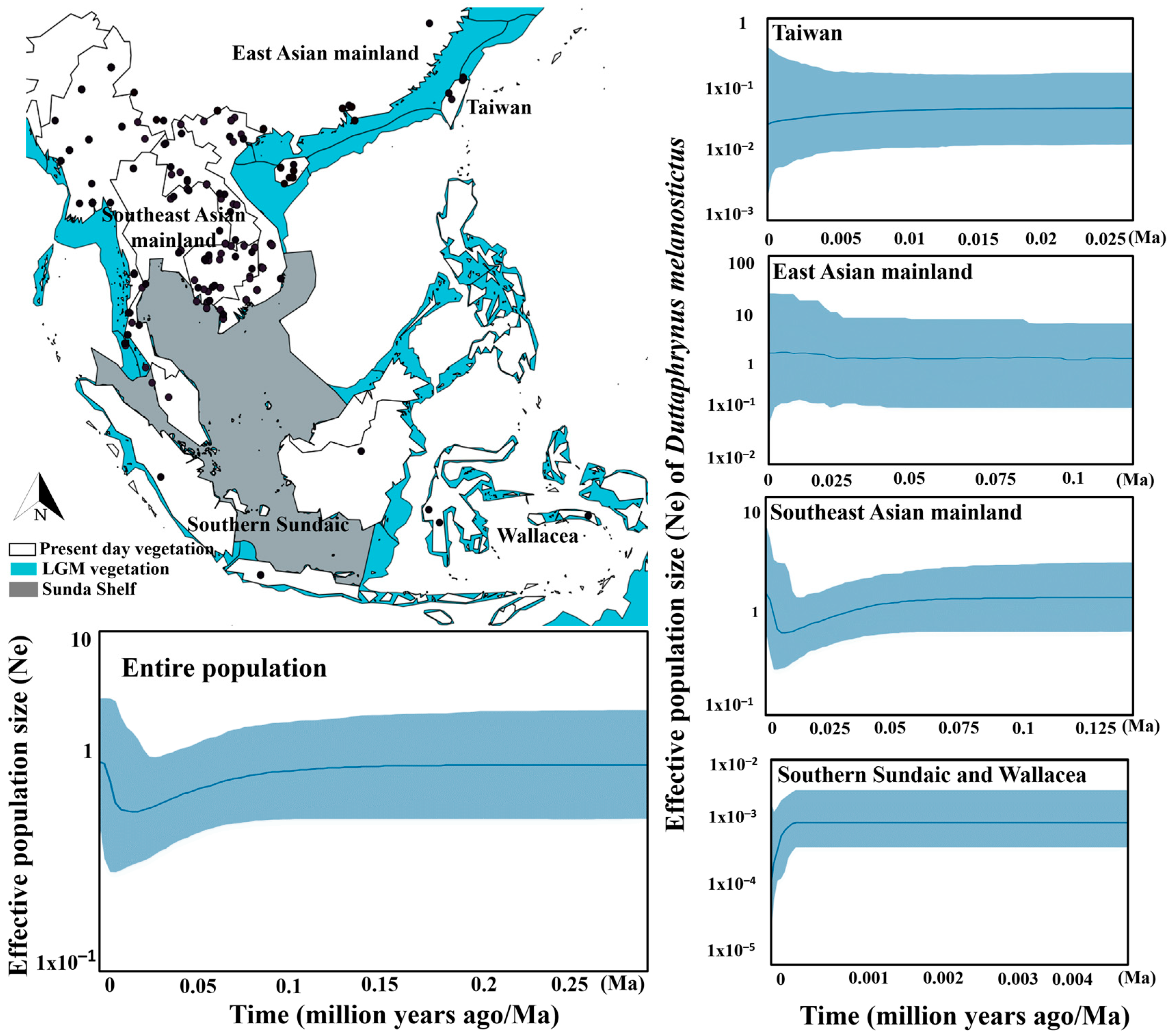
3.4. Population Dynamic over the Last Glacial Maximum
3.5. Population Cluster and Genetic Admixtures
3.6. Population Genetics and Isolation by Distance
4. Discussion
4.1. Vicariance during the Lower Pleistocene
4.2. Coastal-Mainland Lineages Split during MPR
4.3. Population Expansion since the LGM
4.4. Land Bridges and Anthropogenic Dispersion
4.5. IBD Outlier Population Resulting from Anthropologic Activities
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hewitt, G.M. Genetic consequences of climatic oscillations in the Quaternary. Philos. Trans. R. Soc. B Biol. Sci. 2004, 359, 183–195. [Google Scholar] [CrossRef]
- Hofreiter, M.; Stewart, J. Ecological Change, Range Fluctuations and Population Dynamics during the Pleistocene. Curr. Biol. 2009, 19, R584–R594. [Google Scholar] [CrossRef] [PubMed]
- Jaeger, J.R.; Riddle, B.R.; Bradford, D.F. Cryptic Neogene vicariance and Quaternary dispersal of the red-spotted toad (Bufo punctatus): Insights on the evolution of North American warm desert biotas. Mol. Ecol. 2005, 14, 3033–3048. [Google Scholar] [CrossRef] [PubMed]
- Arntzen, J.W.; Recuero, E.; Canestrelli, D.; Martínez-Solano, I. How complex is the Bufo bufo species group? Mol. Phylogenet. Evol. 2013, 69, 1203–1208. [Google Scholar] [CrossRef]
- Fan, Q.; Ma, H.; Hou, G. Late Pleistocene lake and glaciation evolution on the northeastern Qinghai-Tibetan Plateau: A review. Environ. Earth Sci. 2012, 66, 625–634. [Google Scholar] [CrossRef]
- Song, W.; Cao, L.-J.; Li, B.-Y.; Gong, Y.-J.; Hoffmann, A.A.; Wei, S.-J. Multiple refugia from penultimate glaciations in East Asia demonstrated by phylogeography and ecological modelling of an insect pest. BMC Evol. Biol. 2018, 18, 1–16. [Google Scholar] [CrossRef]
- Zhang, D.; Fengquan, L.; Jianmin, B. Eco-environmental effects of the Qinghai-Tibet Plateau uplift during the Quaternary in China. Environ. Geol. 2000, 39, 1352–1358. [Google Scholar] [CrossRef]
- Liu, J.; Wang, C.; Fu, D.; Hu, X.; Xie, X.; Liu, P.; Zhang, Q.; Li, M. Phylogeography of Nanorana parkeri (Anura: Ranidae) and multiple refugia on mitochondrial and nuclear DNA. Sci. Rep. 2015, 5, 9857. [Google Scholar] [CrossRef][Green Version]
- Wang, B.; Xie, F.; Li, J.; Wang, G.; Li, C.; Jiang, J. Phylogeographic investigation and ecological niche modelling of the endemic frog species Nanorana pleskei revealed multiple refugia in the eastern Tibetan Plateau. PeerJ 2017, 5, e3770. [Google Scholar] [CrossRef]
- Dong, B.; Che, J.; Ding, L.; Huang, S.; Murphy, R.W.; Zhao, E.; Zhang, Y. Testing hypotheses of Pleistocene population history using coalescent simulations: Refugial isolation and secondary contact in Pseudepidalea raddei (amphibia: Bufonidae). Asian Herpetol. Res. 2012, 3, 103–113. [Google Scholar] [CrossRef]
- Borzée, A.; Santos, J.L.; Sánchez-RamÍrez, S.; Bae, Y.; Heo, K.; Jang, Y.; Jowers, M.J. Phylogeographic and population insights of the Asian common toad (Bufo gargarizans) in Korea and China: Population isolation and expansions as response to the ice ages. PeerJ 2017, 5, e4044. [Google Scholar] [CrossRef] [PubMed]
- Mead, J.I.; Wilkins, J.; Collins, P.W. Late Quaternary Chorus Frog (Pseudacris) from the Channel Islands, California. Bull. South. Calif. Acad. Sci. 2018, 117, 52–63. [Google Scholar] [CrossRef]
- Schmitt, T. Molecular biogeography of Europe: Pleistocene cycles and postglacial trends. Front. Zool. 2007, 4, 1–13. [Google Scholar] [CrossRef]
- Roberts, P.; Delson, E.; Miracle, P.; Ditchfield, P.; Roberts, R.G.; Jacobs, Z.; Blinkhorn, J.; Ciochon, R.L.; Fleagle, J.G.; Frost, S.R.; et al. Continuity of mammalian fauna over the last 200,000 y in the Indian subcontinent. Proc. Natl. Acad. Sci. USA 2014, 111, 5848–5853. [Google Scholar] [CrossRef] [PubMed]
- Yodthong, S.; Siler, C.D.; Prasankok, P.; Aowphol, A. Phylogenetic patterns of the southeast Asian tree frog Chiromantis hansenae in Thailand. Asian Herpetol. Res. 2014, 5, 179–196. [Google Scholar] [CrossRef]
- Hertwig, S.T.; Das, I.; Schweizer, M.; Brown, R.; Haas, A. Phylogenetic relationships of the Rhacophorus everetti-group and implications for the evolution of reproductive modes in Philautus (Amphibia: Anura: Rhacophoridae). Zool. Scr. 2012, 41, 29–46. [Google Scholar] [CrossRef]
- Che, J.; Zhou, W.W.; Hu, J.S.; Yan, F.; Papenfuss, T.J.; Wake, D.B.; Zhang, Y.P. Spiny frogs (Paini) illuminate the history of the Himalayan region and Southeast Asia. Proc. Natl. Acad. Sci. USA 2010, 107, 13765–13770. [Google Scholar] [CrossRef]
- Matsui, M.; Tominaga, A.; Liu, W.; Khonsue, W.; Grismer, L.L.; Diesmos, A.C.; Das, I.; Sudin, A.; Yambun, P.; Yong, H.; et al. Phylogenetic relationships of Ansonia from Southeast Asia inferred from mitochondrial DNA sequences: Systematic and biogeographic implications (Anura: Bufonidae). Mol. Phylogenet. Evol. 2010, 54, 561–570. [Google Scholar] [CrossRef]
- Inger, R.F.; Stuart, B.L.; Iskandar, D.T. Systematics of a widespread Southeast Asian frog, Rana chalconota (Amphibia: Anura: Ranidae). Zool. J. Linn. Soc. 2009, 155, 123–147. [Google Scholar] [CrossRef]
- Gower, D.J.; Kupfer, A.; Oommen, O.V.; Himstedt, W.; Nussbaum, R.A.; Loader, S.P.; Presswell, B.; Müller, H.; Krishna, S.B.; Boistel, R.; et al. A molecular phylogeny of Ichthyophiid caecilians (Amphibia: Gymnophiona: Ichthyophiidae): Out of India or out of South East Asia? Proc. R. Soc. B Biol. Sci. 2002, 269, 1563–1569. [Google Scholar] [CrossRef]
- Brown, R.M.; Linkem, C.W.; Siler, C.D.; Sukumaran, J.; Esselstyn, J.A.; Diesmos, A.C.; Iskandar, D.T.; Bickford, D.; Evans, B.J.; McGuire, J.A.; et al. Phylogeography and historical demography of Polypedates leucomystax in the islands of Indonesia and the Philippines: Evidence for recent human-mediated range expansion? Mol. Phylogenet. Evol. 2010, 57, 598–619. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.O.; Grismer, L.L. To split or not to split? Multilocus phylogeny and molecular species delimitation of southeast Asian toads (family: Bufonidae). BMC Evol. Biol. 2019, 19, 95. [Google Scholar] [CrossRef]
- Rodríguez, J.; Mateos, A.; Hertler, C.; Palombo, M.R. Modelling human presence and environmental dynamics during the Mid-Pleistocene Revolution: New approaches and tools. Quat. Int. 2016, 393, 19–23. [Google Scholar] [CrossRef]
- Maslin, M.A.; Brierley, C.M. The role of orbital forcing in the Early Middle Pleistocene Transition. Quat. Int. 2015, 389, 47–55. [Google Scholar] [CrossRef]
- Verstappen, H.T. Quaternary Climatic Changes and Natural Environment in SE Asia. GeoJournal 1980, 4, 45–54. [Google Scholar] [CrossRef]
- Hanebuth, T.J.J.; Voris, H.K.; Yokoyama, Y.; Saito, Y.; Okuno, J. Formation and fate of sedimentary depocentres on Southeast Asia’s Sunda Shelf over the past sea-level cycle and biogeographic implications. Earth Sci. Rev. 2011, 104, 92–110. [Google Scholar] [CrossRef]
- Heaney, L.R. Mammalian species richness on islands on the Sunda Shelf, Southeast Asia. Int. Assoc. Ecol. 1984, 61, 11–17. [Google Scholar] [CrossRef]
- Morley, R.J. Palynological evidence for Tertiary plant dispersals in the SE Asian region in relation to plate tectonics and climate. In Biogeography and Geological Evolution of SE Asia; Backhuys Publishers: Leiden, The Netherlands, 1998; pp. 211–234. [Google Scholar]
- Hall, R. The palaeogeography of Sundaland and Wallacea since the Late Jurassic. J. Limnol. 2013, 72, 1–17. [Google Scholar] [CrossRef]
- Sartono, S. On Pleistocene Migration Routes of Vertebrate Fauna. Bull. Geol. Soc. Malays. 1973, 6, 273–286. [Google Scholar] [CrossRef]
- Stelbrink, B. A Biogeographic View on Southeast Asia’s History. Ph.D. Thesis, Faculty of Life Sciences, Humboldt University of Berlin, Berlin, Germany, 2015. [Google Scholar]
- Metcalfe, I. Tectonic evolution of Sundaland Tectonic evolution of Sundaland. Bull. Geol. Soc. Malays. 2017, 63, 27–60. [Google Scholar] [CrossRef]
- Hall, R. Sundaland and Wallacea: Geology, plate tectonics and palaeogeography. In Biotic Evolution and Environmental Change in Southeast Asia; Gower, D., Ohnson, K., Richardson, J., Rosen, B., Rüber, L., Williams, S., Eds.; Cambridge University Press: New York, NY, USA, 2012; pp. 32–78. [Google Scholar]
- Raes, N.; Cannon, C.H.; Hijmans, R.J.; Slik, J.W.F.; van Welzen, P.C.; Saw, L.G.; Piessens, T.; Raes, N. Historical distribution of Sundaland’s Dipterocarp rainforests at Quaternary glacial maxima. Proc. Natl. Acad. Sci. USA 2014, 111, 16790–16795. [Google Scholar] [CrossRef]
- Harrison, T.; Krigbaum, J.; Manser, J. Primate Biogeography and Ecology on the Sunda Shelf Islands: A Paleontological and Zooarchaeological Perspective. Primate Biogeogr. 2006, 331–372. [Google Scholar] [CrossRef]
- Sathiamurthy, E.; Voris, K.H. Maps of Holocene Sea Level Transgression and Submerged Lakes on the Sunda Shelf. Nat. Hist. J. Chulalongkorn Univ. 2006, 2, 1–44. [Google Scholar]
- Hewitt, G.M. The genetic legacy of the Quaternary ice ages. Nature 2000, 405, 907–913. [Google Scholar] [CrossRef]
- Reeves, J.M.; Bostock, H.C.; Ayliffe, L.K.; Barrows, T.T.; De Deckker, P.; Devriendt, L.S.; Dunbar, G.B.; Drysdale, R.N.; Fitzsimmons, K.E.; Gagan, M.K.; et al. Palaeoenvironmental change in tropical Australasia over the last 30,000 years—A synthesis by the OZ-INTIMATE group. Quat. Sci. Rev. 2013, 74, 97–114. [Google Scholar] [CrossRef]
- Hope, G.; Kershaw, A.P.; van der Kaars, S.; Xiangjun, S.; Liew, P.M.; Heusser, L.E.; Takahara, H.; McGlone, M.; Miyoshi, N.; Moss, P.T. History of vegetation and habitat change in the Austral-Asian region. Quat. Int. 2004, 118, 103–126. [Google Scholar] [CrossRef]
- Woodruff, D.S. Biogeography and conservation in Southeast Asia: How 2.7 million years of repeated environmental fluctuations affect today’s patterns and the future of the remaining refugial-phase biodiversity. Biodivers. Conserv. 2010, 19, 919–941. [Google Scholar] [CrossRef]
- Mulcahy, D.G.; Lee, J.L.; Miller, A.H.; Chand, M.; Thura, M.K.; Zug, G.R. Filling the bins of life: Report of an amphibian and reptile survey of the Tanintharyi (Tenasserim) Region of Myanmar, with DNA barcode data. ZooKeys 2018, 85–152. [Google Scholar] [CrossRef]
- Garg, S.; Biju, S.D. New microhylid frog genus from Peninsular India with Southeast Asian affinity suggests multiple Cenozoic biotic exchanges between India and Eurasia. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Inger, R.F.; Voris, H.K. The biogeographical relations of the frogs and snakes of Sundaland. J. Biogeogr. 2001, 28, 863–891. [Google Scholar] [CrossRef]
- Prasad, K.N. Pleistocene cave fauna from peninsular India. J. Cave Karst Stud. 1996, 58, 30–34. [Google Scholar]
- Lydekker, R. The fauna of the Kurnool Caves. Palaeontol. Indica Ser. X 1886, 4, 23–58. [Google Scholar]
- Patnaik, R.; Badam, G.L.; Murty, M.L.K. Additional vertebrate remains from one of the Late Pleistocene-Holocene Kurnool Caves (Muchchatla Chintamanu Gavi) of South India. Quat. Int. 2008, 192, 43–51. [Google Scholar] [CrossRef]
- Tingley, R.; García-Díaz, P.; Arantes, C.R.R.; Cassey, P. Integrating transport pressure data and species distribution models to estimate invasion risk for alien stowaways. Ecography 2018, 41, 635–646. [Google Scholar] [CrossRef]
- Church, G. The Invasion of Bali by Bufo melanostictus. Herpetologica 1960, 16, 15–21. [Google Scholar]
- Moore, M.; Niaina Fidy, J.F.S.; Edmonds, D. The new toad in town: Distribution of the Asian toad, Duttaphrynus melanostictus, in the Toamasina area of eastern Madagascar. Trop. Conserv. Sci. 2015, 8, 440–455. [Google Scholar] [CrossRef]
- Trainor, R. The Black-spined Toad Bufo melanostictus invading Timor-Leste: Identification, distribution, public health and biodiversity impacts. Presentation of AusAID funded collaborative survey of Black-spined Toad in Timor-Leste, Dili, Timor-Leste, September 2009. [Google Scholar]
- Mo, M. Asian Black-spined Toads (Duttaphrynus melanostictus) in Australia: An invasion worth avoiding. IRCF Reptiles Amphib. 2017, 24, 155–161. [Google Scholar]
- Jang-Liaw, N.-H.; Chou, W.-H. Anuran Fauna of Taiwan and Adjacent Islands Based on Valid Specimen Records. Coll. Res. 2015, 28, 5–53. [Google Scholar]
- Van Dijk, P.P.; Iskandar, D.; Lau, M.W.N.; Huiqing, G.; Baorong, G.; Kuangyang, L.; Wenhao, C.; Zhigang, Y.; Chan, B.; Dutta, S.; et al. Duttaphrynus melanostictus (errata version published in 2016). IUCN Red List Threat. Species 2004, 2004. [Google Scholar] [CrossRef]
- Wogan, G.O.U.; Stuart, B.L.; Iskandar, D.T.; McGuire, J.A. Deep genetic structure and ecological divergence in a widespread human commensal toad. Biol. Lett. 2016, 12. [Google Scholar] [CrossRef]
- Vences, M.; Brown, J.L.; Lathrop, A.; Rosa, G.M.; Cameron, A.; Crottini, A.; Dolch, R.; Edmonds, D.; Freeman, K.L.M.; Glaw, F.; et al. Tracing a toad invasion: Lack of mitochondrial DNA variation, haplotype origins, and potential distribution of introduced Duttaphrynus melanostictus in Madagascar. Amphib. Reptilia 2017, 38, 197–207. [Google Scholar] [CrossRef]
- Stuart, B.L.; Inger, R.F.; Voris, H.K. High level of cryptic species diversity revealed by sympatric lineages of Southeast Asian forest frogs. Biol. Lett. 2006, 2, 470–474. [Google Scholar] [CrossRef] [PubMed]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed]
- Múrias Dos Santos, A.; Cabezas, M.P.; Tavares, A.I.; Xavier, R.; Branco, M. TcsBU: A tool to extend TCS network layout and visualization. Bioinformatics 2015, 32, 627–628. [Google Scholar] [CrossRef] [PubMed]
- Leigh, J.W.; Bryant, D. POPART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Mardulyn, P. Trees and⁄or networks to display intraspecific DNA sequence variation? Mol. Ecol. 2012, 3385–3390. [Google Scholar] [CrossRef]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. Partitionfinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. Mrbayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Lakner, C.; Van Der Mark, P.; Huelsenbeck, J.P.; Larget, B.; Ronquist, F. Efficiency of markov chain monte carlo tree proposals in bayesian phylogenetics. Syst. Biol. 2008, 57, 86–103. [Google Scholar] [CrossRef]
- Drummond, A.J.; Rambaut, A.; Shapiro, B.; Pybus, O.G. Bayesian coalescent inference of past population dynamics from molecular sequences. Mol. Biol. Evol. 2005, 22, 1185–1192. [Google Scholar] [CrossRef]
- Kozlov, A.M.; Darriba, D.; Morel, B.; Stamatakis, A. RAxML-NG: A fast, scalable, and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics 2018, 35, 4453–4455. [Google Scholar] [CrossRef] [PubMed]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [PubMed]
- Kolby, J.E.; Kraus, F.; Rabemananjara, F.C.E.; Rabesihanaka, S.; Rabibisoa, N.H.C.; Randrianantoandro, C.; Randrianiana, R.D.; Raxworthy, C.J.; Razafimahatratra, B.; Robsomanitrandrasana, E. Stop Madagascar’s toad invasion now. Nat. Corresp. 2014, 509, 563. [Google Scholar] [CrossRef] [PubMed]
- Swofford, D.L. Phylogenetic Analysis Using Parsimony PAUP* 4.0 Beta Version Disclaimer and User Agreement; Sinauer Associates: Sunderland, MA, USA, 2002; pp. 1–144. [Google Scholar]
- Pond, S.L.K.; Frost, S.D.W.; Muse, S.V. HyPhy: Hypothesis testing using phylogenies. Bioinformatics 2005, 21, 676–679. [Google Scholar] [CrossRef] [PubMed]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef]
- Tong, H.; Wo, Y.; Liao, P.; Jin, Y. Phylogenetic, demographic and dating analyses of Bufo gargarizans populations from the Zhoushan Archipelago and Mainland China. Asian Herpetol. Res. 2017, 8, 165–173. [Google Scholar] [CrossRef]
- Fu, J.; Weadick, C.J.; Zeng, X.; Wang, Y.; Liu, Z.; Zheng, Y.; Li, C.; Hu, Y. Phylogeographic analysis of the Bufo gargarizans species complex: A revisit. Mol. Phylogenet. Evol. 2005, 37, 202–213. [Google Scholar] [CrossRef]
- Deng, W.; Maust, B.S.; Nickle, D.C.; Learn, G.H.; Liu, Y.; Heath, L.; Pond, S.L.K.; Mullins, J.I. DIVEIN: A web server to analyze phylogenies, sequence divergence, diversity, and informative sites. BioTechniques 2010, 48, 405–408. [Google Scholar] [CrossRef]
- Kjer, K.M.; Honeycutt, R.L. Site specific rates of mitochondrial genomes and the phylogeny of eutheria. BMC Evol. Biol. 2007, 7, 1–9. [Google Scholar] [CrossRef][Green Version]
- Suchard, M.A.; Lemey, P.; Baele, G.; Ayres, D.L.; Drummond, A.J.; Rambaut, A. Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10. Virus Evol. 2018, 4, 1–5. [Google Scholar] [CrossRef]
- Yu, Y.; Harris, A.J.; Blair, C.; He, X. RASP (Reconstruct Ancestral State in Phylogenies): A tool for historical biogeography. Mol. Phylogenet. Evol. 2015, 87, 46–49. [Google Scholar] [CrossRef]
- Fu, Y.X. Statistical tests of neutrality of mutations against population growth, hitchhiking and background selection. Genetics 1997, 147, 915–925. [Google Scholar]
- Tajima, F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 1989, 123, 585–595. [Google Scholar] [PubMed]
- Ramos-Onsins, S.E.; Rozas, J. Erratum: Statistical properties of new neutrality tests against population growth (Molecular Biology and Evolution (2002) 19 (2092–2100)). Mol. Biol. Evol. 2006, 23, 1642. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Bouckaert, R.; Vaughan, T.G.; Barido-Sottani, J.; Duchêne, S.; Fourment, M.; Gavryushkina, A.; Heled, J.; Jones, G.; Kühnert, D.; De Maio, N.; et al. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef] [PubMed]
- Flot, J.F. CHAMPURU 1.0: A computer software for unraveling mixtures of two DNA sequences of unequal lengths. Mol. Ecol. Notes 2007, 7, 974–977. [Google Scholar] [CrossRef]
- Garrick, R.C.; Sunnucks, P.; Dyer, R.J. Nuclear gene phylogeography using PHASE: Dealing with unresolved genotypes, lost alleles, and systematic bias in parameter estimation. BMC Evol. Biol. 2010, 10. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Wen, X.; Falush, D. Documentation for Structure Software: Version 2.3; University of Chicago: Chicago, IL, USA, 2009. [Google Scholar]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Early, R.; Bradley, B.A.; Dukes, J.S.; Lawler, J.J.; Olden, J.D.; Blumenthal, D.M.; Gonzalez, P.; Grosholz, E.D.; Ibañez, I.; Miller, L.P.; et al. Global threats from invasive alien species in the twenty-first century and national response capacities. Nat. Commun. 2016, 7. [Google Scholar] [CrossRef]
- Jakobsson, M.; Rosenberg, N.A. CLUMPP: A cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics 2007, 23, 1801–1806. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, R.K.; Ramasamy, S.; Bindroo, B.B.; Naik, V.G. STRUCTURE PLOT: A program for drawing elegant STRUCTURE bar plots in user friendly interface. SpringerPlus 2014, 3, 1–3. [Google Scholar] [CrossRef]
- Mantel, N. The Detection of Disease Clustering and a Generalized Regression Approach. Nature 1967, 70, 279–280. [Google Scholar] [CrossRef]
- Miller, M.P.; Davis, R.J.; Forsman, E.D.; Mullins, T.D.; Haig, S.M. Isolation by distance versus landscape resistance: Understanding dominant patterns of genetic structure in Northern Spotted Owls (Strix occidentalis caurina). PLoS ONE 2018, 13, e0201720. [Google Scholar] [CrossRef]
- Ji, X.; Kuman, K.; Clarke, R.J.; Forestier, H.; Li, Y.; Ma, J.; Qiu, K.; Li, H.; Wu, Y. The oldest Hoabinhian technocomplex in Asia (43.5 ka) at Xiaodong rockshelter, Yunnan Province, southwest China. Quat. Int. 2016, 400, 166–174. [Google Scholar] [CrossRef]
- Yang, M.A.; Gao, X.; Theunert, C.; Tong, H.; Aximu-Petri, A.; Nickel, B.; Slatkin, M.; Meyer, M.; Pääbo, S.; Kelso, J.; et al. 40,000-Year-Old Individual from Asia Provides Insight into Early Population Structure in Eurasia. Curr. Biol. 2017, 27, 3202–3208.e9. [Google Scholar] [CrossRef] [PubMed]
- Mccoll, H.; Racimo, F.; Vinner, L.; Demeter, F.; Gakuhari, T.; Moreno-mayar, J.V.; Van Driem, G.; Wilken, U.G.; Seguin-orlando, A.; De la Fuente Castro, C.; et al. The prehistoric peopling of Southeast Asia. Hum. Genom. 2018, 92, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Gomes, S.M.; Bodner, M.; Souto, L.; Zimmermann, B.; Huber, G.; Strobl, C.; Röck, A.W.; Achilli, A.; Olivieri, A.; Torroni, A.; et al. Human settlement history between Sunda and Sahul: A focus on East Timor (Timor-Leste) and the Pleistocenic mtDNA diversity. BMC Genom. 2015, 16, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Brandão, A.; Eng, K.K.; Rito, T.; Cavadas, B.; Bulbeck, D.; Gandini, F.; Pala, M.; Mormina, M.; Hudson, B.; White, J.; et al. Quantifying the legacy of the Chinese Neolithic on the maternal genetic heritage of Taiwan and Island Southeast Asia. Hum. Genet. 2016, 135, 363–376. [Google Scholar] [CrossRef] [PubMed]
- QGIS Development Team QGIS Geographic Information System. Open Source Geospatial Foundation Project. Available online: http://qgis.osgeo.org (accessed on 2 January 2019).
- Ray, N.; Adams, J.M.; Ray, N. A GIS-based Vegetation Map of the World at the Last Glacial Maximum (25,000-15,000 BP). Internet Archaeology. Volume 11. 2001. Available online: http://intarch.ac.uk/journal/issue11/rayadams_toc.html (accessed on 21 August 2018).
- Flanders Marine Institute. IHO Sea Areas. Version 3. 2018. Available online: https://www.marineregions.org/ (accessed on 20 January 2019).
- Macey, J.R.; Larson, A.; Fang, Z.; Tuniyev, B.S. Phylogenetic Relationships of Toads in the. Mol. Phylogenet. Evol. 1998, 9, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Yu, G.; Rao, D.; Yang, J. Phylogeography and demographic history of babina pleuraden (anura, ranidae) in southwestern china. PLoS ONE 2012, 7, e34013. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Zhou, W.; Zhao, H.; Yuan, Z.; Wang, Y.; Jiang, K.; Jin, J.; Murphy, R.W.; Che, J.; Zhang, Y. Geological events play a larger role than Pleistocene climatic fluctuations in driving the genetic structure of Quasipaa boulengeri (Anura: Dicroglossidae). Mol. Ecol. 2013, 22, 1120–1133. [Google Scholar] [CrossRef] [PubMed]
- Van Bocxlaer, I.; Biju, S.D.; Loader, S.P.; Bossuyt, F. Toad radiation reveals into-India dispersal as a source of endemism in the Western Ghats-Sri Lanka biodiversity hotspot. BMC Evol. Biol. 2009, 9, 131. [Google Scholar] [CrossRef] [PubMed]
- Biju, S.D.; Van Bocxlaer, I.; Giri, V.; Loader, S.; Bossuyt, F. Two new endemic genera and a new species of toad (Anura: Bufonidae) from the Western Ghats of India. BMC Res. Notes 2009, 2, 1–6. [Google Scholar] [CrossRef]
- Meijer, H.J.M.; Due, R.A.; Sutikna, T.; Saptomo, W.; Jatmiko; Wasisto, S.; Tocheri, M.W.; Mayr, G. Late Pleistocene songbirds of Liang Bua (Flores, Indonesia); the first fossil passerine fauna described from Wallacea. PeerJ 2017, 2017, 1–25. [Google Scholar] [CrossRef]
- Evans, B.J.; McGuire, J.A.; Brown, R.M.; Andayani, N.; Supriatna, J. A coalescent framework for comparing alternative models of population structure with genetic data: Evolution of Celebes toads. Biol. Lett. 2008, 4, 430–433. [Google Scholar] [CrossRef][Green Version]
- Arruda, M.P.; Costa, W.P.; Recco-Pimentel, S.M. Genetic diversity of morato’s digger toad, Proceratophrys moratoi: Spatial structure, gene flow, effective size and the need for differential management strategies of populations. Genet. Mol. Biol. 2017, 40, 502–514. [Google Scholar] [CrossRef]
- Li, Q.; Zheng, F.; Chen, M.; Xiang, R.; Qiao, P.; Shao, L.; Cheng, X. Glacial paleoceanography off the mouth of the Mekong River, southern South China Sea, during the last 500ka. Quat. Res. 2010, 73, 563–572. [Google Scholar] [CrossRef]
- Pan, T.; Zhang, Y.; Wang, H.; Wu, J.; Kang, X.; Qian, L.; Chen, J.; Rao, D.; Jiang, J.; Zhang, B. The reanalysis of biogeography of the Asian tree frog, Rhacophorus (Anura: Rhacophoridae): Geographic shifts and climatic change influenced the dispersal process and diversification. PeerJ 2017, 5, 1–25. [Google Scholar] [CrossRef]
- Song, G.; Qu, Y.; Yin, Z.; Li, S.; Liu, N.; Lei, F. Phylogeography of the Alcippe morrisonia (Aves: Timaliidae): Long population history beyond late Pleistocene glaciations. BMC Evol. Biol. 2009, 9, 1–11. [Google Scholar] [CrossRef]
- Zhisheng, A.; Kutzbach, J.E.; Prell, W.L.; Porter, S.C. Evolution of Asian monsoons and phased uplift of the Himalaya-Tibetan plateau since Late Miocene times. Nature 2001, 411, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Chalk, T.B.; Hain, M.P.; Foster, G.L.; Rohling, E.J.; Sexton, P.F.; Badger, M.P.S.; Cherry, S.G.; Hasenfratz, A.P.; Haug, G.H.; Jaccard, S.L.; et al. Causes of ice age intensification across the Mid-Pleistocene transition. Proc. Natl. Acad. Sci. USA 2017, 114, 13114–13119. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Sigman, D.M.; Martínez-García, A.; Anderson, R.F.; Chen, M.T.; Ravelo, A.C.; Straub, M.; Wong, G.T.F.; Haug, G.H. Impact of glacial/interglacial sea level change on the ocean nitrogen cycle. Proc. Natl. Acad. Sci. USA 2017, 114, E6759–E6766. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Ree, R.H. Uplift-driven diversification in the Hengduan Mountains, a temperate biodiversity hotspot. Proc. Natl. Acad. Sci. USA 2017, 114, E3444–E3451. [Google Scholar] [CrossRef] [PubMed]
- Dever, J.A.; Fuiten, A.M.; Konu, Ö.; Wilkinson, J.A. Cryptic Torrent Frogs of Myanmar: An examination of the Amolops marmoratus species complex with the resurrection of Amolops afghanus and the Identification of a new species. Copeia 2012, 2012, 57–76. [Google Scholar] [CrossRef]
- Yang, J.-H.; Zeng, Z.-C.; Wang, Y.-Y. Description of two new sympatric species of the genus Leptolalax (Anura: Megophryidae) from western Yunnan of China. PeerJ 2018, 6, e4586. [Google Scholar] [CrossRef]
- Jacobs, J.B. A History of Pre-Invasion Taiwan. Taiwan Hist. Res. 2016, 23, 1–38. [Google Scholar]
- Oshida, T.; Lee, J.; Lin, L.; Chen, Y. Phylogeography of Pallas’s Squirrel in Taiwan: Geographical isolation in an arboreal small mammal. Am. Soc. Mammal. 2006, 87, 247–254. [Google Scholar] [CrossRef][Green Version]
- Wu, C.R.; Chao, S.Y.; Hsu, C. Transient, seasonal and interannual variability of the Taiwan Strait current. J. Oceanogr. 2007, 63, 821–833. [Google Scholar] [CrossRef]
- Lu, J.; Li, S.P.; Wu, Y.; Jiang, L. Are Hong Kong and Taiwan stepping-stones for invasive species to the mainland of China? Ecol. Evol. 2018, 8, 1966–1973. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, G.R.; Brodie, E.D.J. Occurrence of amphibians in saline habitats: A review and evolutionary perspective. Herpetol. Monogr. 2015, 29, 1–27. [Google Scholar] [CrossRef]
- Reilly, S.B.; Wogan, G.O.U.; Stubbs, A.L.; Arida, E.; Iskandar, D.T.; McGuire, J.A. Toxic toad invasion of Wallacea: A biodiversity hotspot characterized by extraordinary endemism. Glob. Change Biol. 2017, 23, 5029–5031. [Google Scholar] [CrossRef] [PubMed]
- Marshall, B.M.; Casewell, N.R.; Vences, M.; Glaw, F.; Andreone, F.; Rakotoarison, A.; Zancolli, G.; Woog, F.; Wüster, W. Widespread vulnerability of Malagasy predators to the toxins of an introduced toad. Curr. Biol. 2018, 28, R654–R655. [Google Scholar] [CrossRef] [PubMed]
- Simanjuntak, T.; Asikin, I.N. Early Holocene Human Settlement in Eastern Java. Indo Pac. Prehist. Assoc. Bull. 2004, 24, 13–19. [Google Scholar]
- Gathorne-Hardy, F.J.; Davies, R.G.; Eggleton, P.; Jones, D.T. Quaternary rainforest refugia in south-east Asia: Using termites (Isoptera) as indicators. Biol. J. Linn. Soc. 2002, 75, 453–466. [Google Scholar] [CrossRef]
- Solihuddin, T. A Drowning Sunda Shelf Model during Last Glacial Maximum (LGM) and Holocene: A Review. Indones. J. Geosci. 2014, 1, 99–107. [Google Scholar] [CrossRef][Green Version]
- Wurster, C.M.; Bird, M.I.; Bull, I.D.; Creed, F.; Bryant, C.; Dungait, J.A.J.; Paz, V. Forest contraction in north equatorial Southeast Asia during the Last Glacial Period. Proc. Natl. Acad. Sci. USA 2010, 107, 15508–15511. [Google Scholar] [CrossRef]
- Licata, F.; Ficetola, G.F.; Freeman, K.; Mahasoa, R.H.; Ravololonarivo, V.; Solofo Niaina Fidy, J.F.; Koto-Jean, A.B.; Nahavitatsara, E.R.; Andreone, F.; Crottini, A. Abundance, distribution and spread of the invasive Asian toad Duttaphrynus melanostictus in eastern Madagascar. Biol. Invasions 2019, 21, 1615–1626. [Google Scholar] [CrossRef]
- McKee, A.M.; Maerz, J.C.; Smith, L.L.; Glenn, T.C. Habitat predictors of genetic diversity for two sympatric wetland-breeding amphibian species. Ecol. Evol. 2017, 7, 6271–6283. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Relevant Nodes | Dating Estimates (Mean/Ma ± SD) | Reconstruction of Ancestral State in Phylogenies Route | Events * (Matrix) | ||
|---|---|---|---|---|---|
| Vicariance | Dispersal | Extinction | |||
| Outgroup Bufo gargarizans | 2.94 (±0.10) | Western China to Myanmar | 1 | 2 | 0 |
| Stem origin of D. melanostictus | 1.85 (±0.77) | Myanmar to southern Sundaic (Sumatra) to Wallacea | 3 | 4 | 0 |
| Stem origin of D. melanostictus clade I | 1.43 (±0.10) | Myanmar to southern Sundaic and Wallacea | 2 | 3 | 0 |
| Stem origin of D. melanostictus clade II and III | 0.84 (±0.32) | Myanmar to eastern Mainland Southeast Asia | 0 | 0 | 0 |
| Crown origin of D. melanostictus complex I | 0.04 (±13.94) | Wallacea to Southern Sundaic | 1 | 2 | 0 |
| Crown origin of D. melanostictus complex II | 0.57 (±1.49) | Myanmar to Peninsular Malaysia | 1 | 2 | 0 |
| Crown origin of D. melanostictus complex III | 0.66 (±0.67) | Myanmar to eastern Mainland Southeast Asia | 0 | 0 | 0 |
| Crown origin of Taiwanese population | 0.06 (±5.96) | Southwestern China to Taiwan | 0 | 0 | 0 |
| Population | Sample Size (No. of Haplotype) | Nucleotide Diversity (π) | Neutrality Tests | Ramos-Onsins and Rozas’s (R2) | Goodness-of Fit Tests | Mismatch Distribution | |||
|---|---|---|---|---|---|---|---|---|---|
| Tajima’s D | p Value | Fu’s Fs | p Value | r (p Value) | |||||
| Taiwan | 23 (3) | 1.882 | 1.855 | 0.972 | 4.891 | 0.974 | 0.2353 | 0.7232 | multimodal |
| Southeastern China | 8 (4) | 2.273 | −0.033 | 0.536 | −0.998 | 0.240 | 0.1572 | 0.0569 | unimodal |
| Hainan | 8 (5) | 1.857 | −0.168 | 0.458 | −1.315 | 0.099 | 0.1779 | 0.0536 | unimodal |
| Southwestern China | 11 (5) | 5.389 | −1.825 | 0.013 | 4.742 | 0.974 | 0.2970 | 0.1667 | multimodal |
| Northern Southeast Asian mainland | 46 (13) | 14.915 | 0.298 | 0.703 | 1.289 | 0.721 | 0.1007 | 0.0117 | multimodal |
| Southern Southeast Asian mainland | 36 (14) | 1.431 | −2.057 | 0.003 | −17.476 | 0.000 | 0.0308 | 0.0474 | unimodal |
| Coastal Myanmar, Thailand and Peninsular Malaysia | 13 (6) | 22.048 | 2.229 | 0.995 | 10.859 | 0.996 | 0.1991 | 0.0817 | multimodal |
| Southern Sundaic | 33 (2) | 9.733 | −2.021 | 0.001 | 7.916 | 1.000 | 0.2262 | 0.2937 | multimodal |
| Wallacea | 5 (1) | 0.000 | 0.000 | 1.000 | 0.000 | - | - | - | - |
| Madagascar | 65 (1) | 0.000 | 0.000 | 1.000 | 0.000 | - | - | - | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Othman, S.N.; Chen, Y.-H.; Chuang, M.-F.; Andersen, D.; Jang, Y.; Borzée, A. Impact of the Mid-Pleistocene Revolution and Anthropogenic Factors on the Dispersion of Asian Black-Spined Toads (Duttaphrynus melanostictus). Animals 2020, 10, 1157. https://doi.org/10.3390/ani10071157
Othman SN, Chen Y-H, Chuang M-F, Andersen D, Jang Y, Borzée A. Impact of the Mid-Pleistocene Revolution and Anthropogenic Factors on the Dispersion of Asian Black-Spined Toads (Duttaphrynus melanostictus). Animals. 2020; 10(7):1157. https://doi.org/10.3390/ani10071157
Chicago/Turabian StyleOthman, Siti N., Yi-Huey Chen, Ming-Feng Chuang, Desiree Andersen, Yikweon Jang, and Amaël Borzée. 2020. "Impact of the Mid-Pleistocene Revolution and Anthropogenic Factors on the Dispersion of Asian Black-Spined Toads (Duttaphrynus melanostictus)" Animals 10, no. 7: 1157. https://doi.org/10.3390/ani10071157
APA StyleOthman, S. N., Chen, Y.-H., Chuang, M.-F., Andersen, D., Jang, Y., & Borzée, A. (2020). Impact of the Mid-Pleistocene Revolution and Anthropogenic Factors on the Dispersion of Asian Black-Spined Toads (Duttaphrynus melanostictus). Animals, 10(7), 1157. https://doi.org/10.3390/ani10071157