A Detailed Study of Rainbow Trout (Onchorhynchus mykiss) Intestine Revealed That Digestive and Absorptive Functions Are Not Linearly Distributed along Its Length

 ,
, 
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples Collection
2.2. Histology and Histochemistry
2.3. Morphometric Evaluation
2.4. Quantitative Stereological Analysis
2.5. Immunohistochemistry
2.6. TUNEL Test
2.7. Statistical Analysis
3. Results
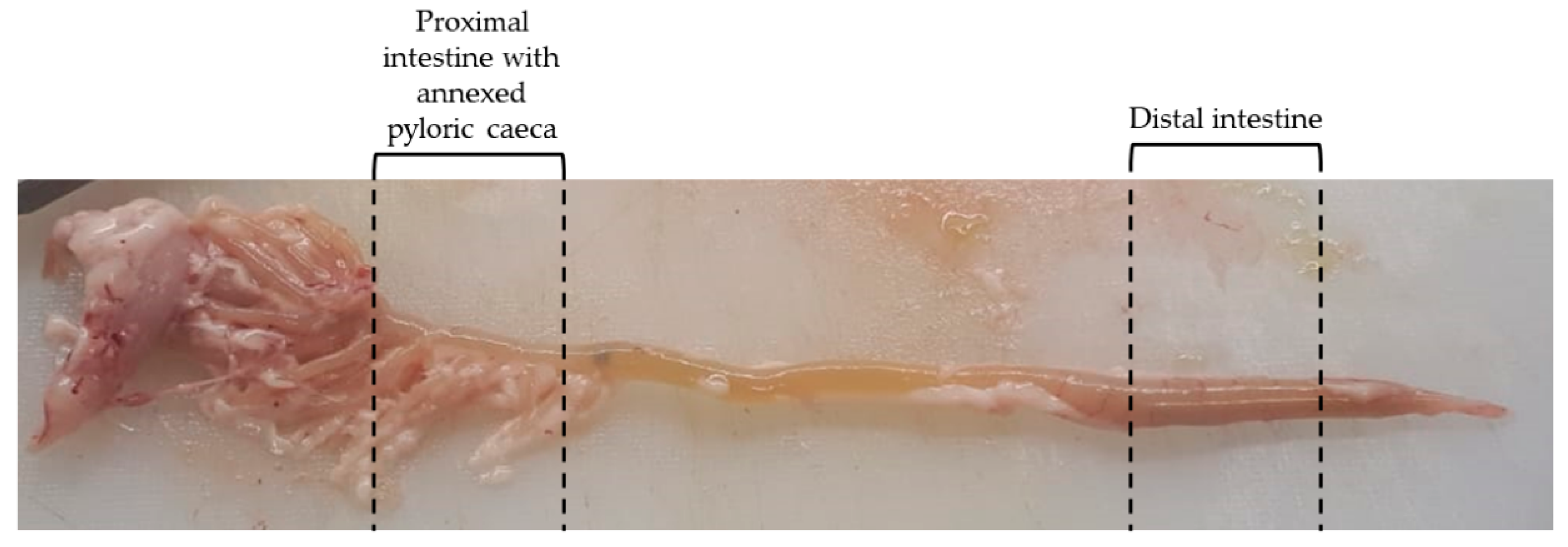
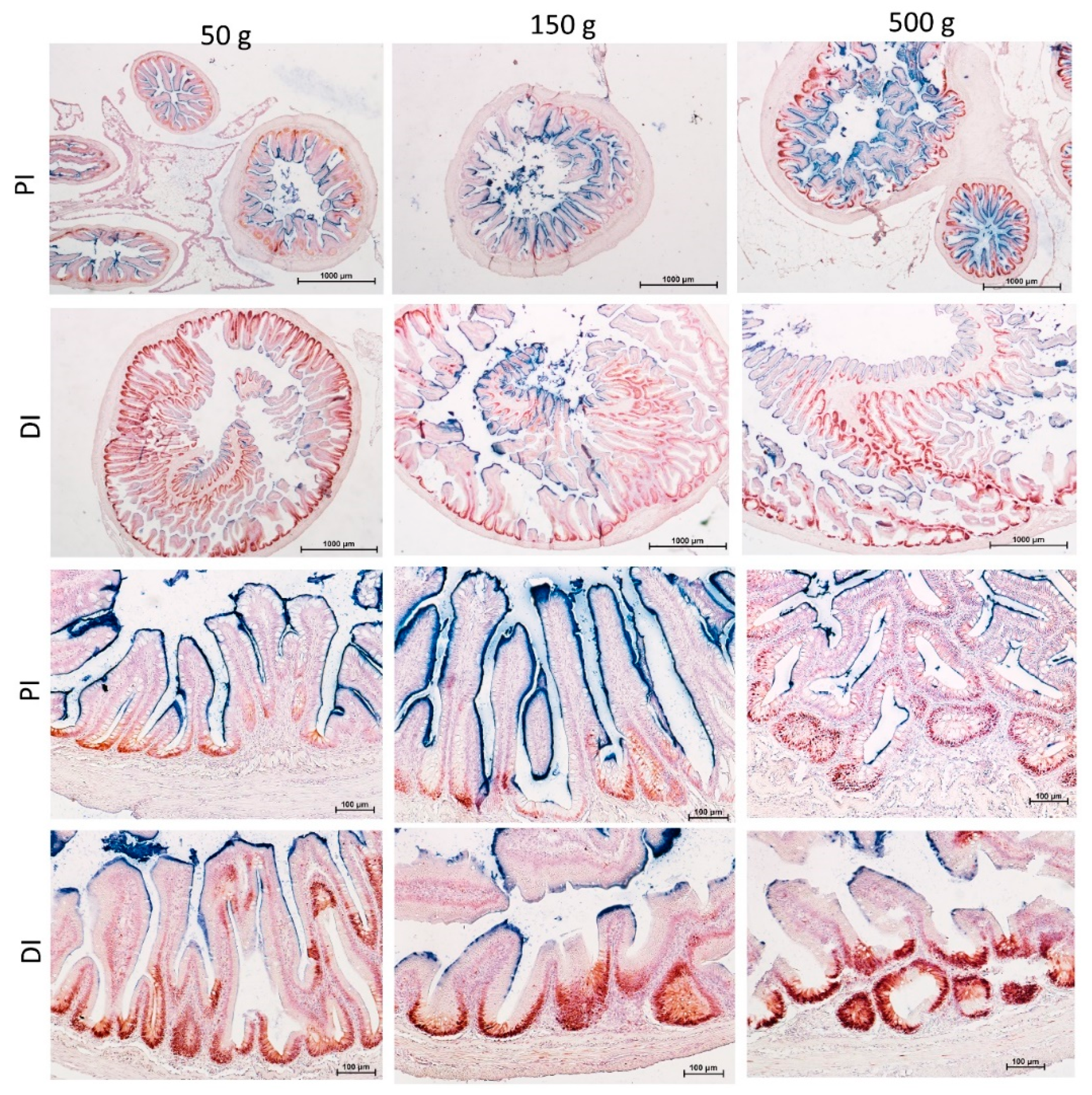
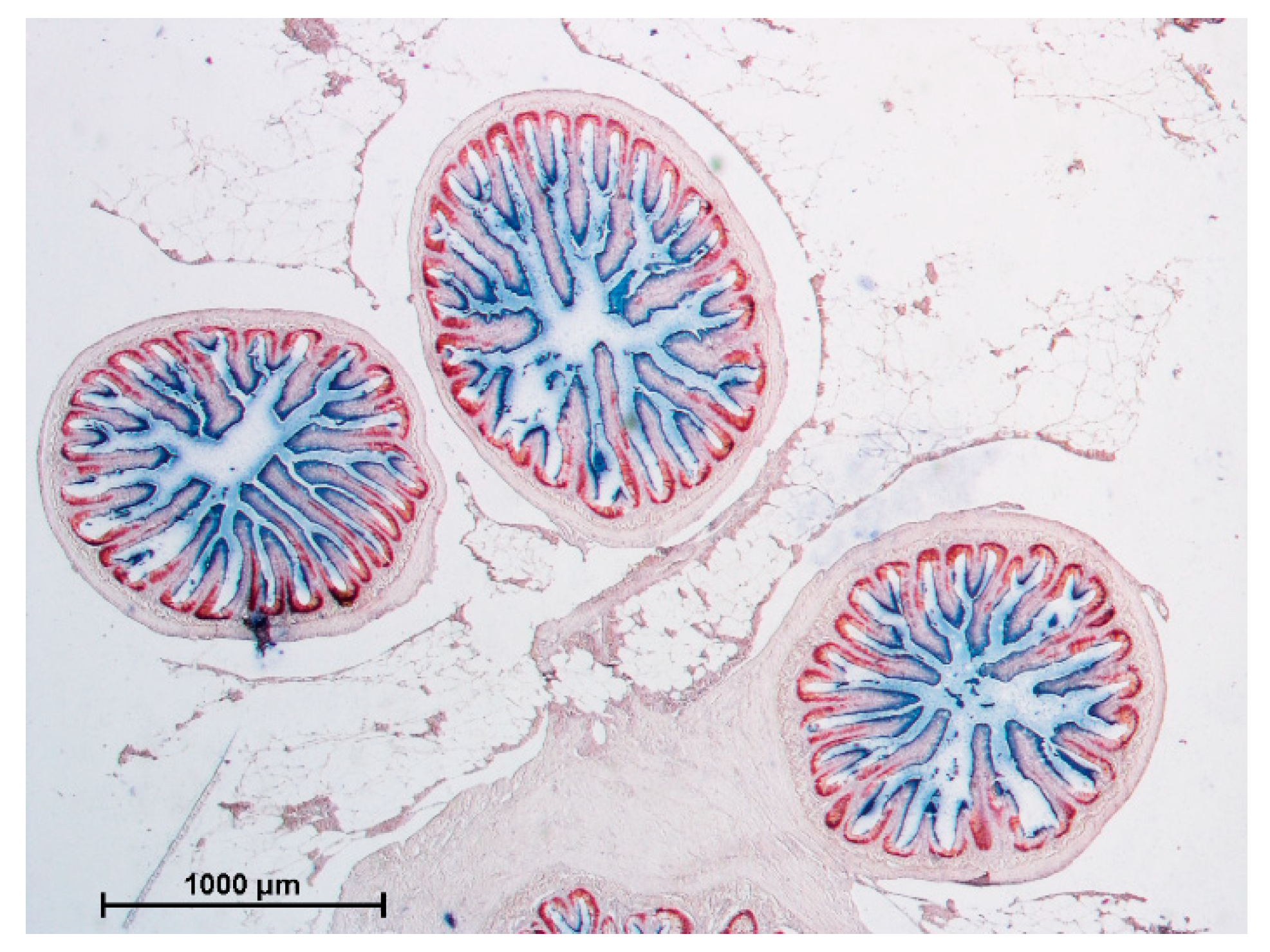
3.1. Gross Anatomy
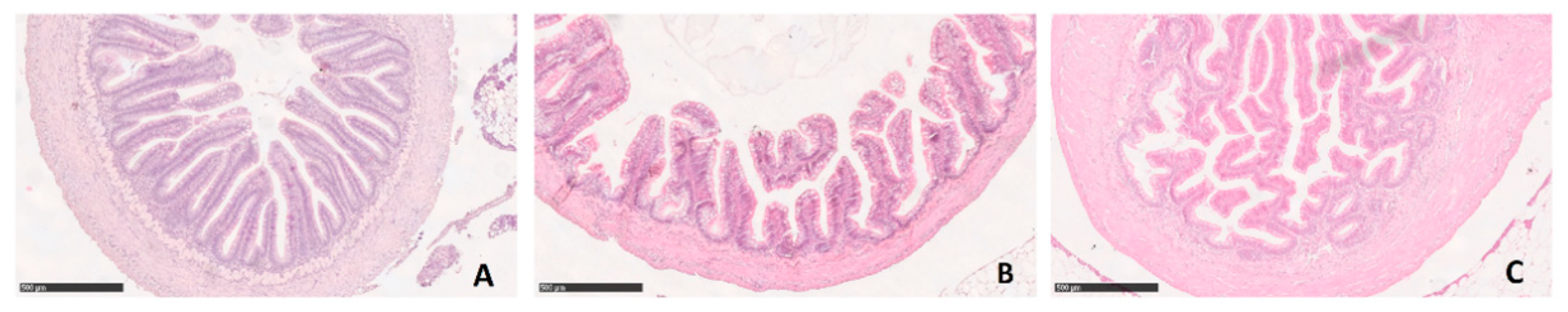
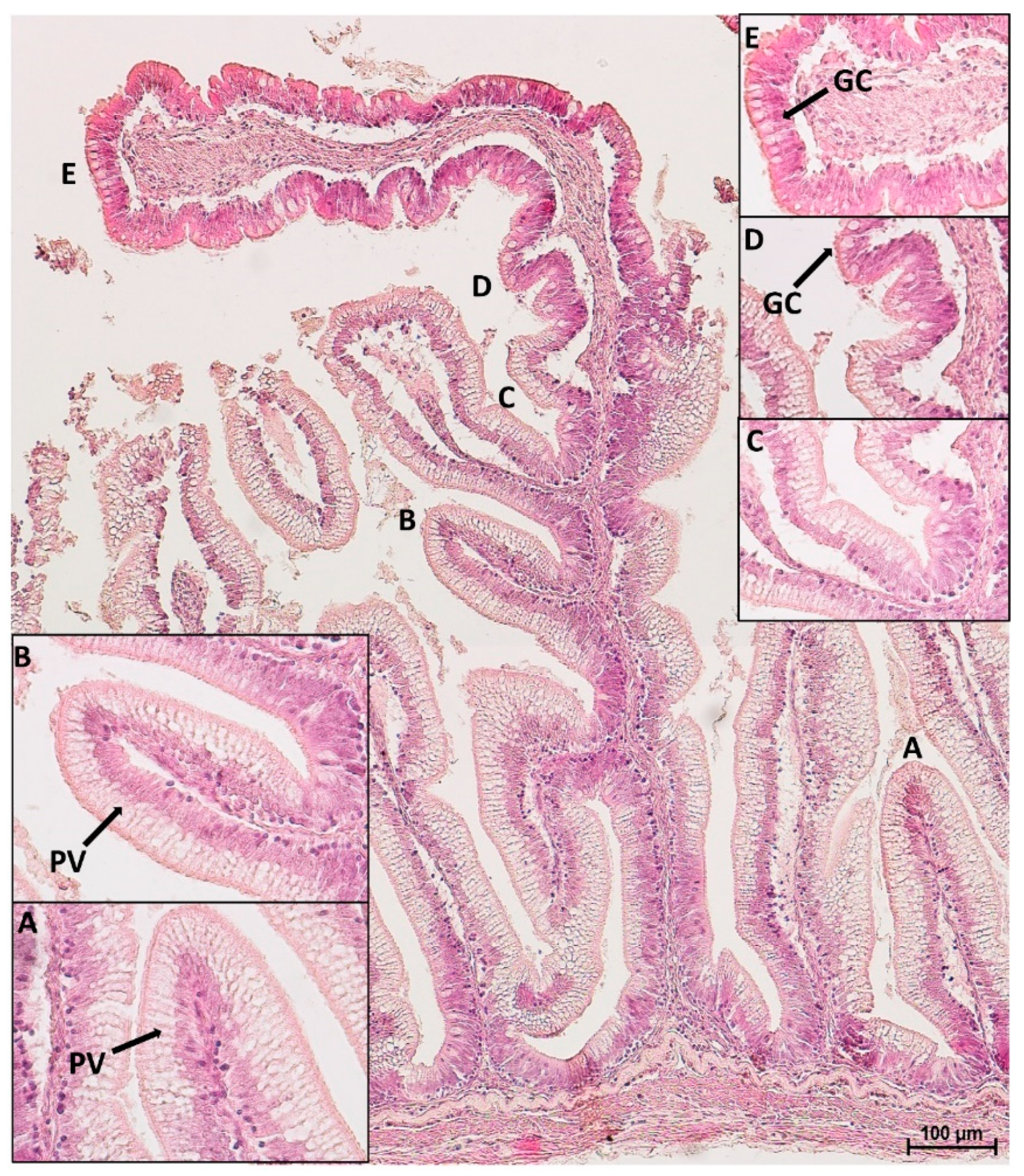
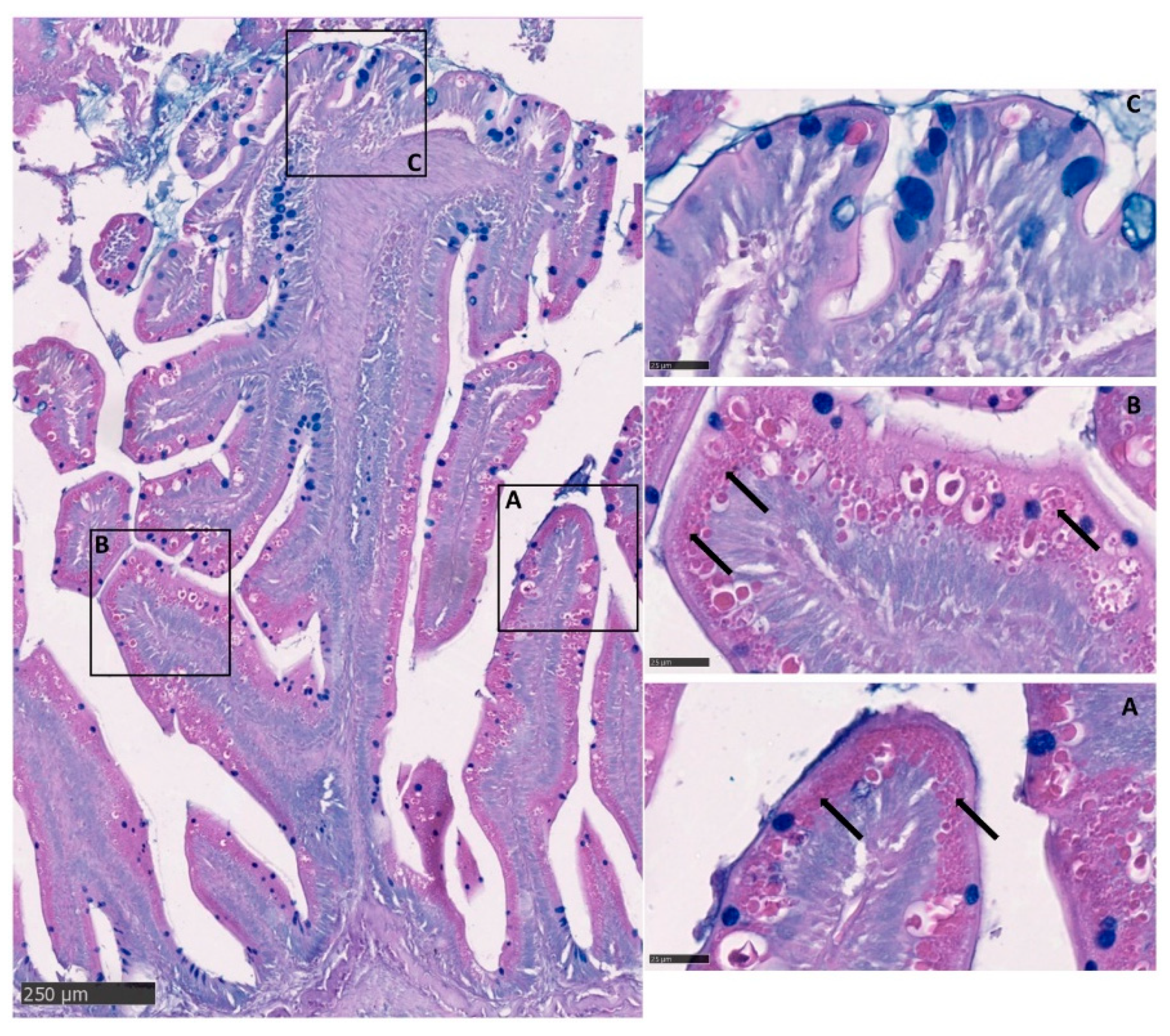
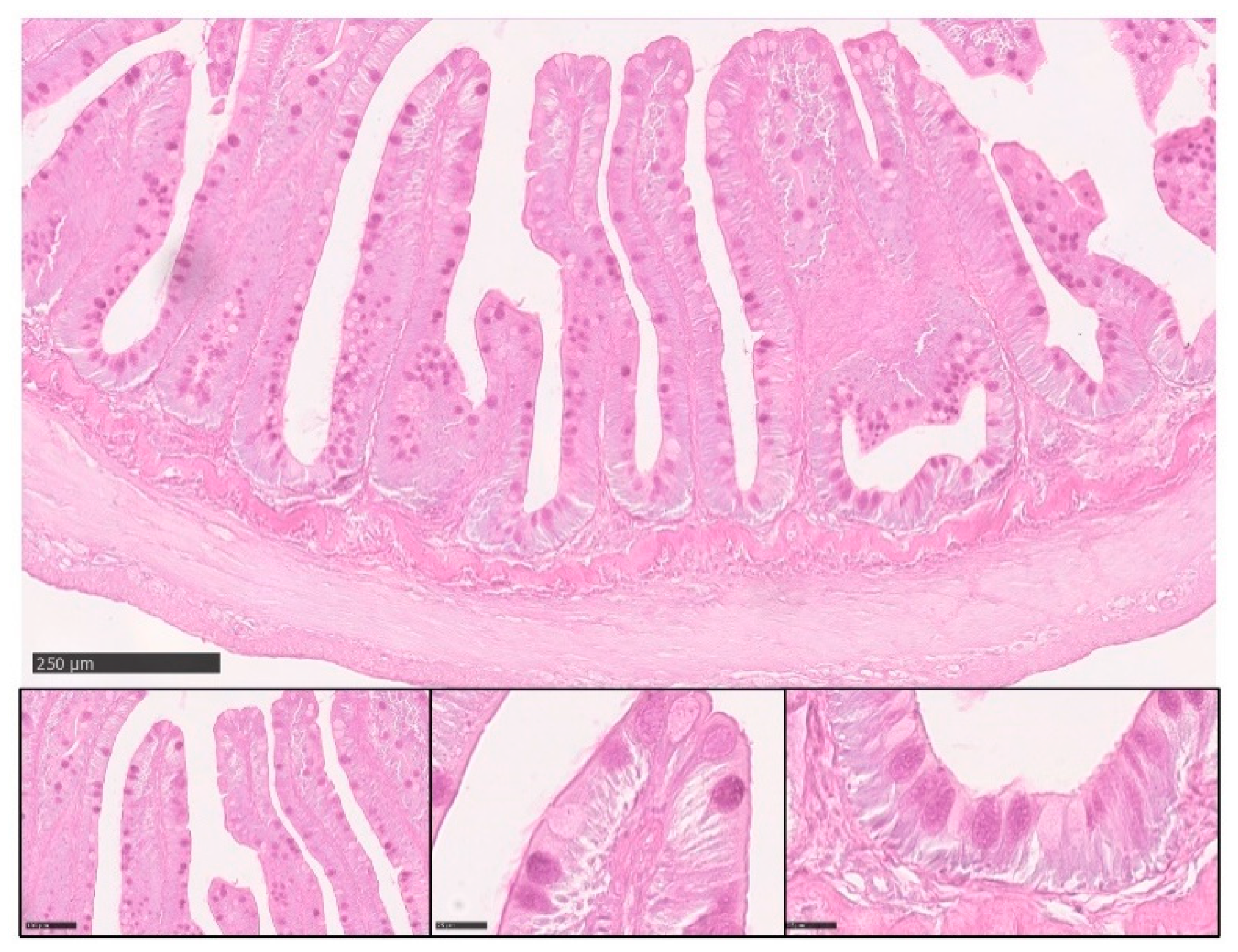
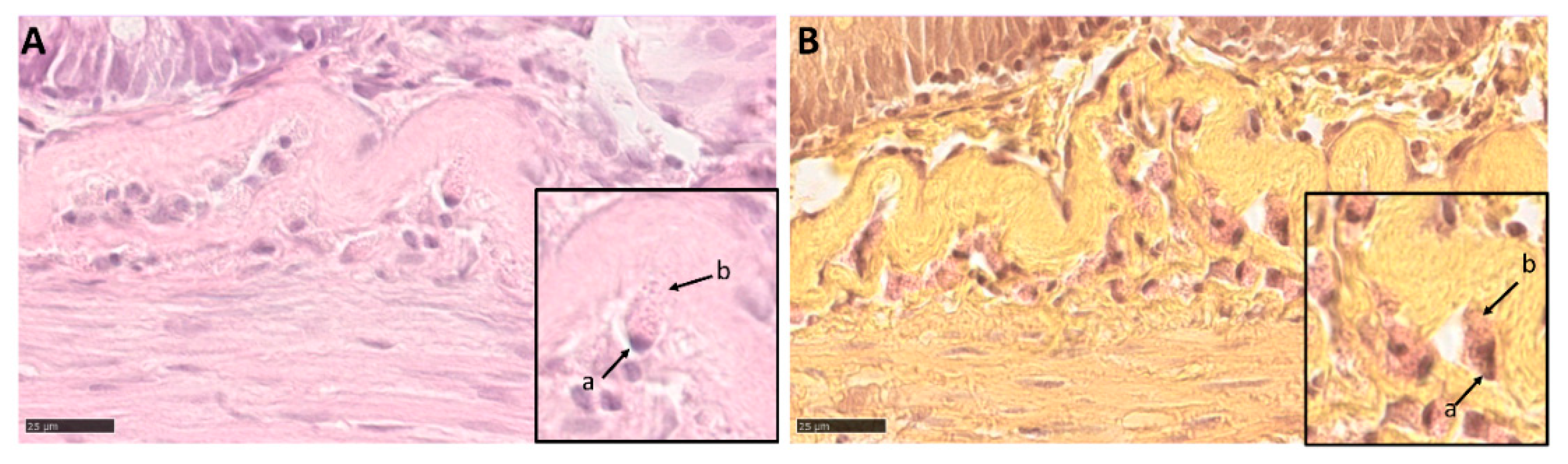
3.2. Microscopical Anatomy
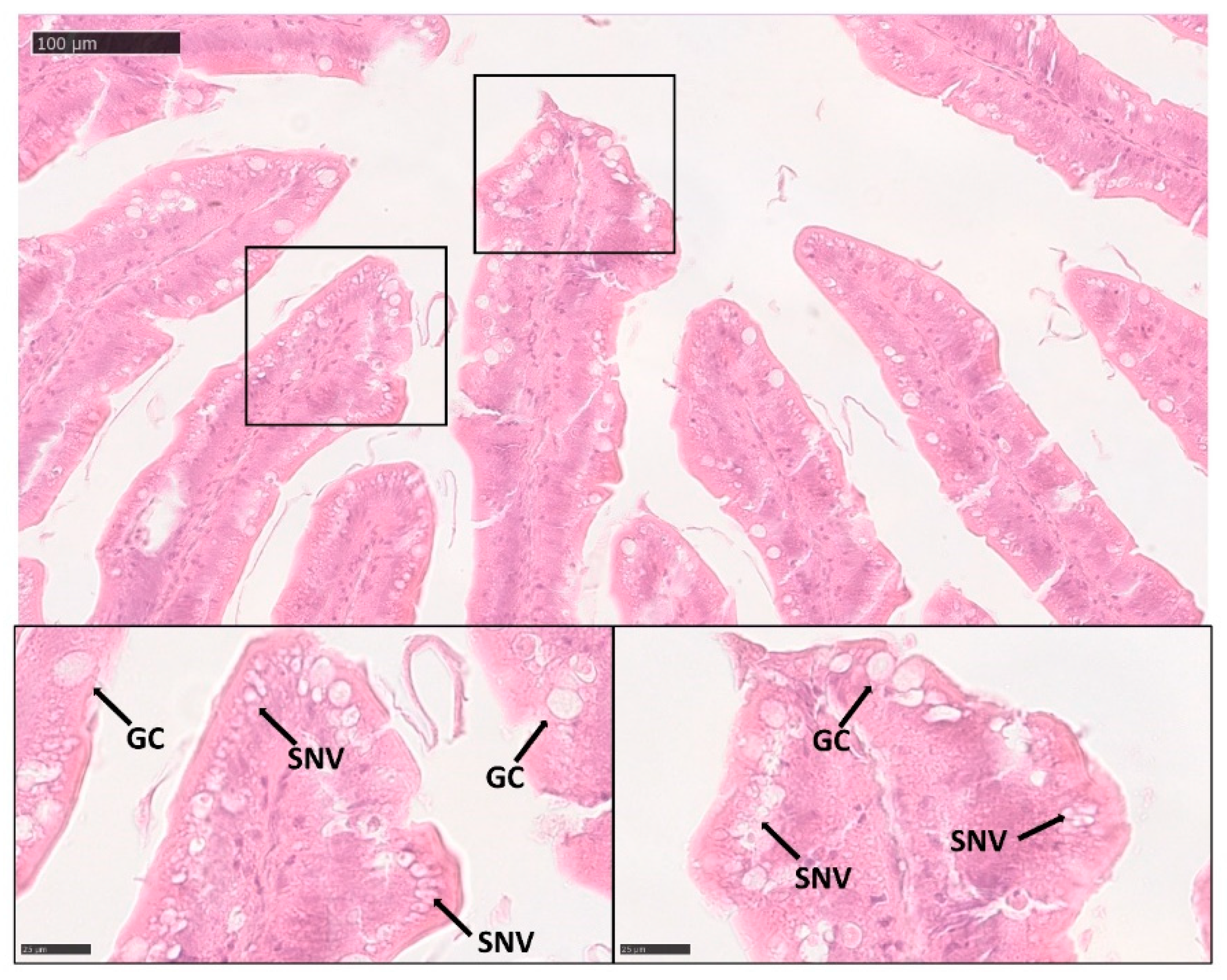
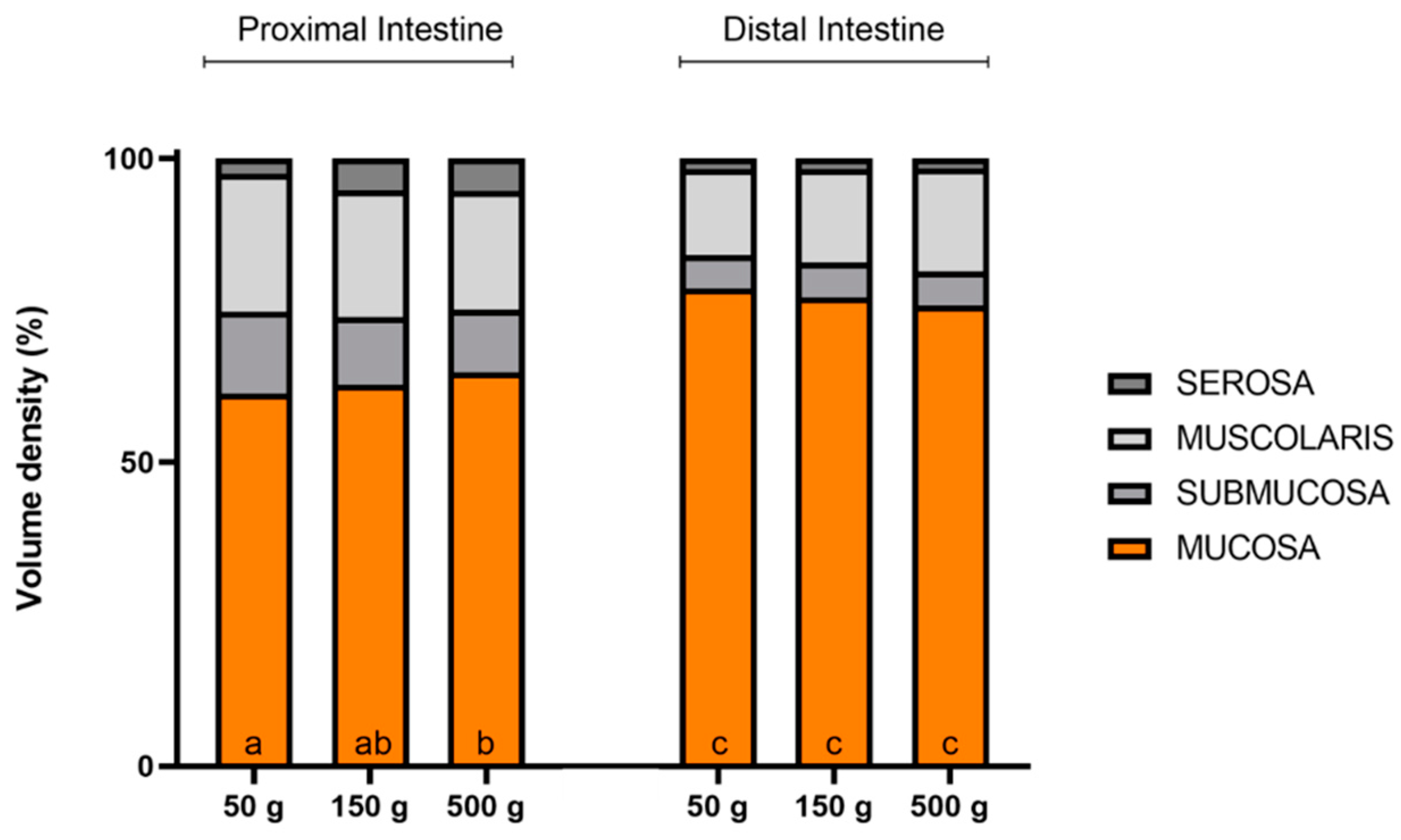
3.3. Quantitative Sterological Analysis
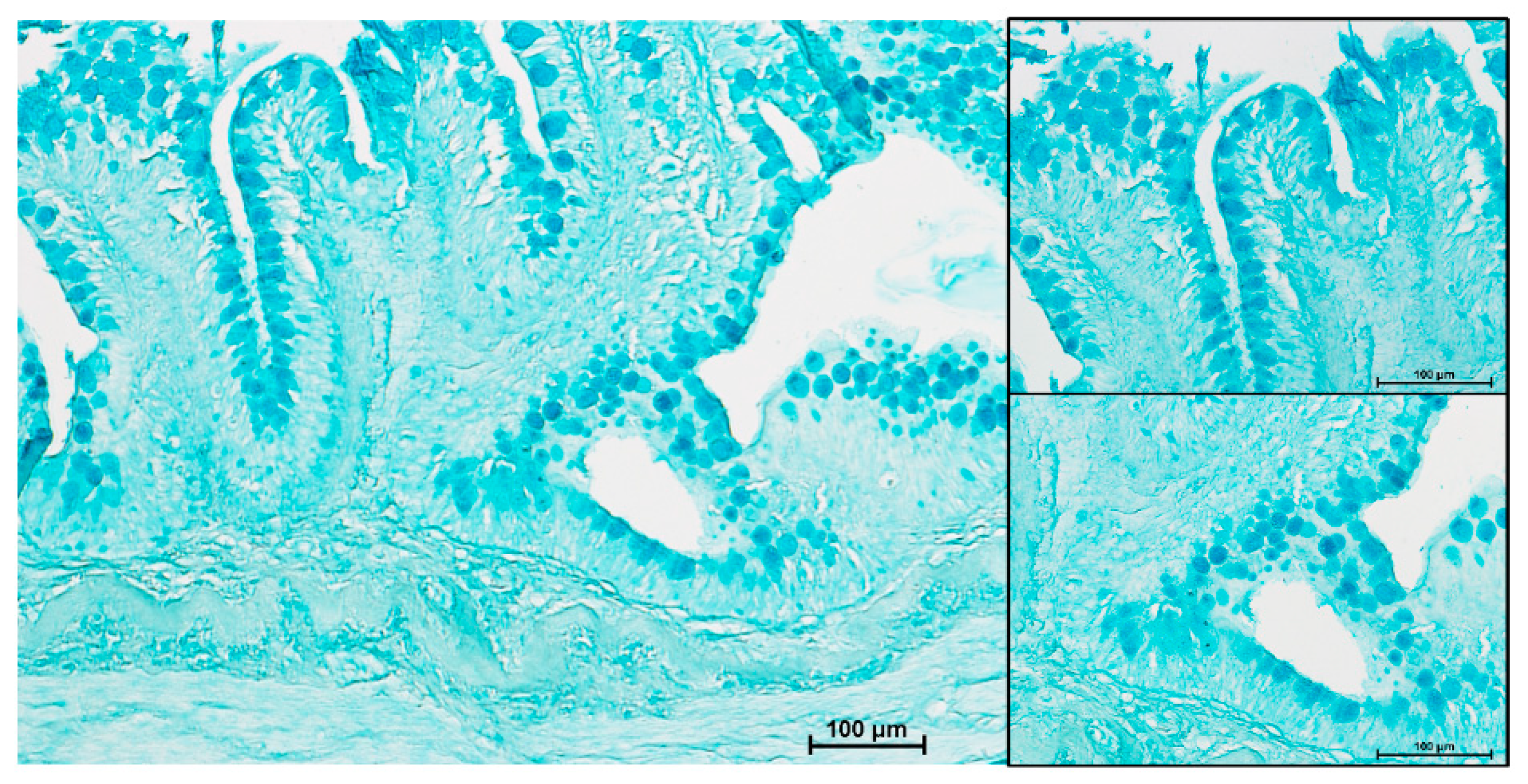
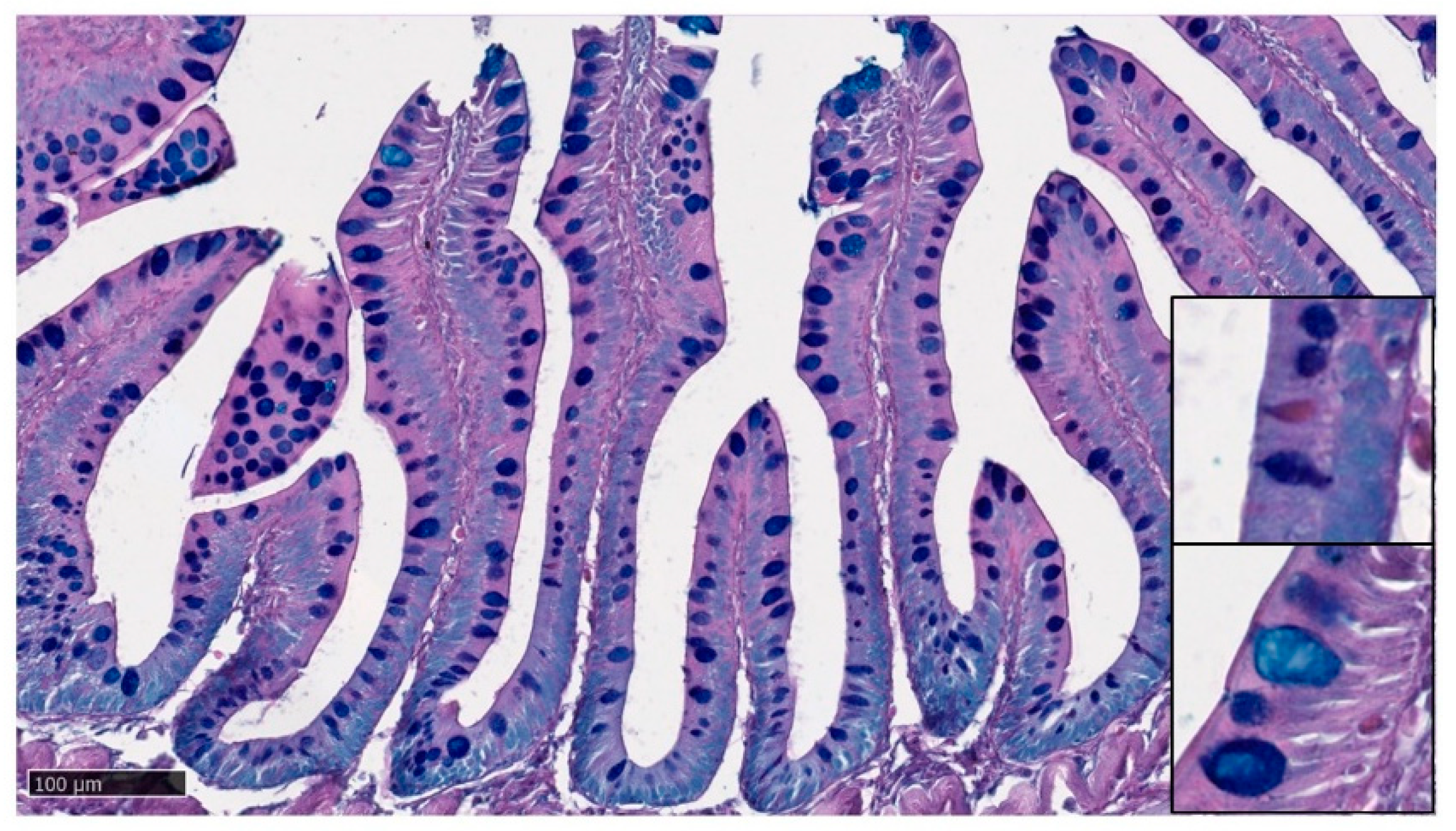
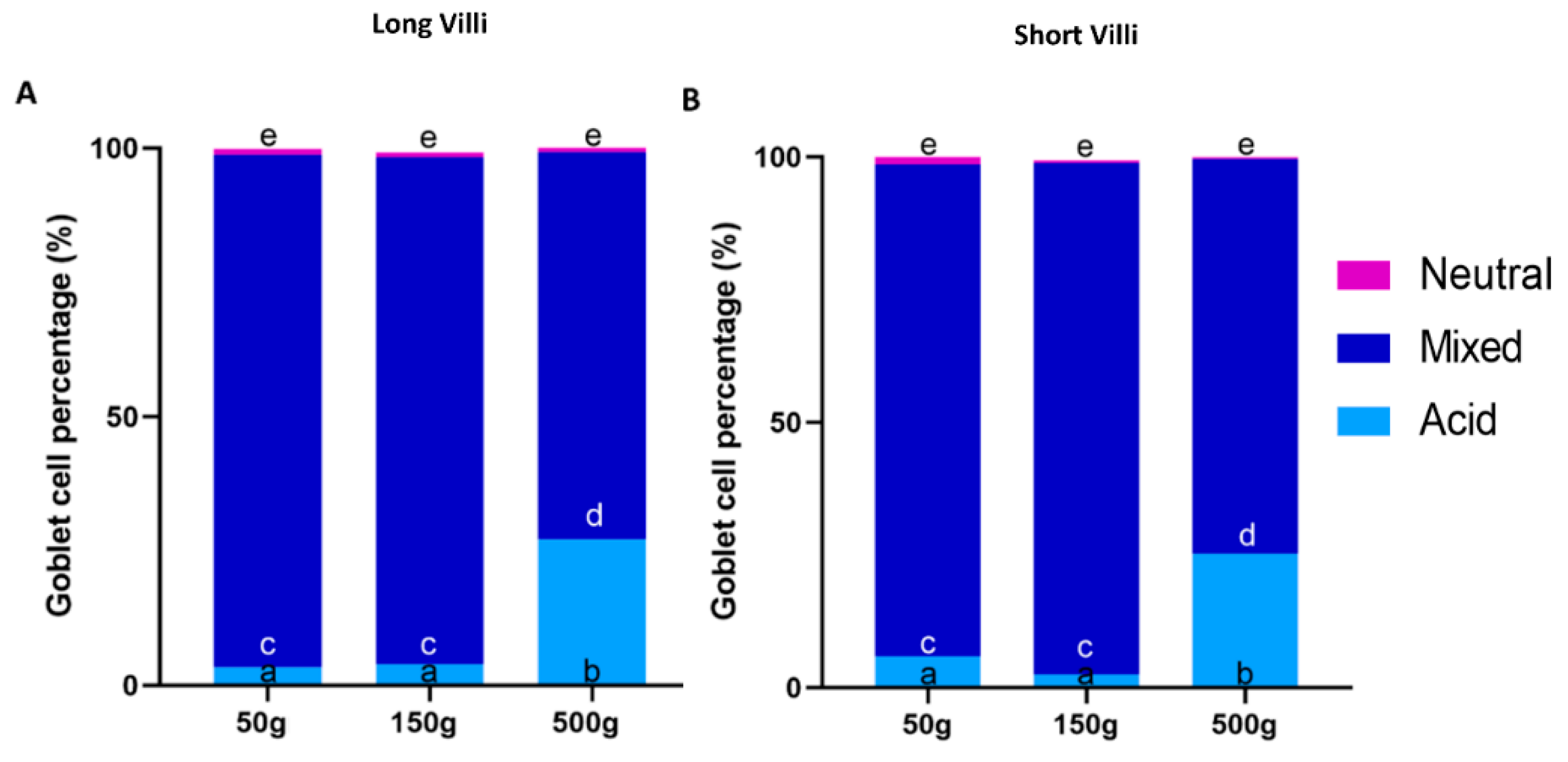
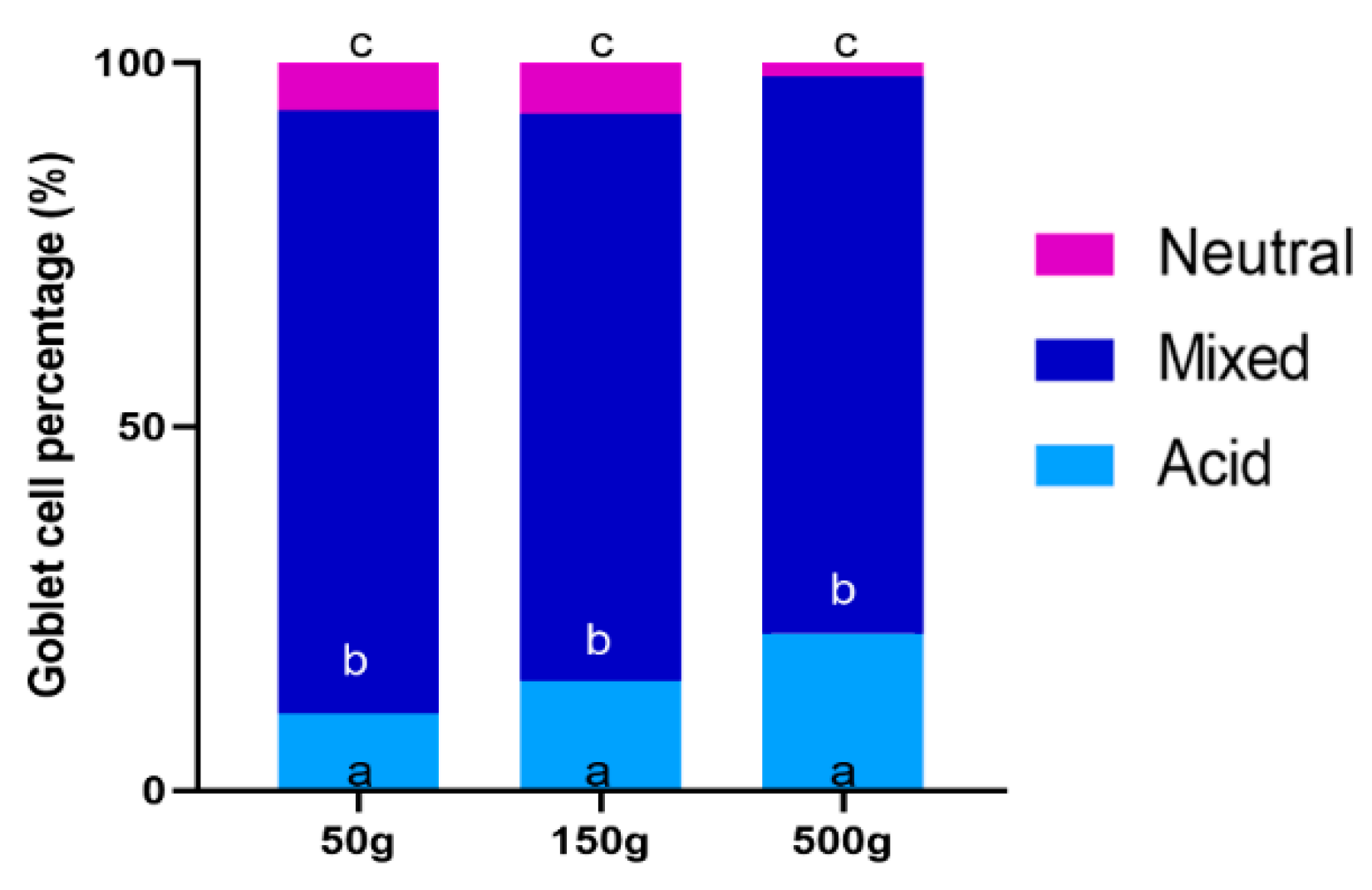
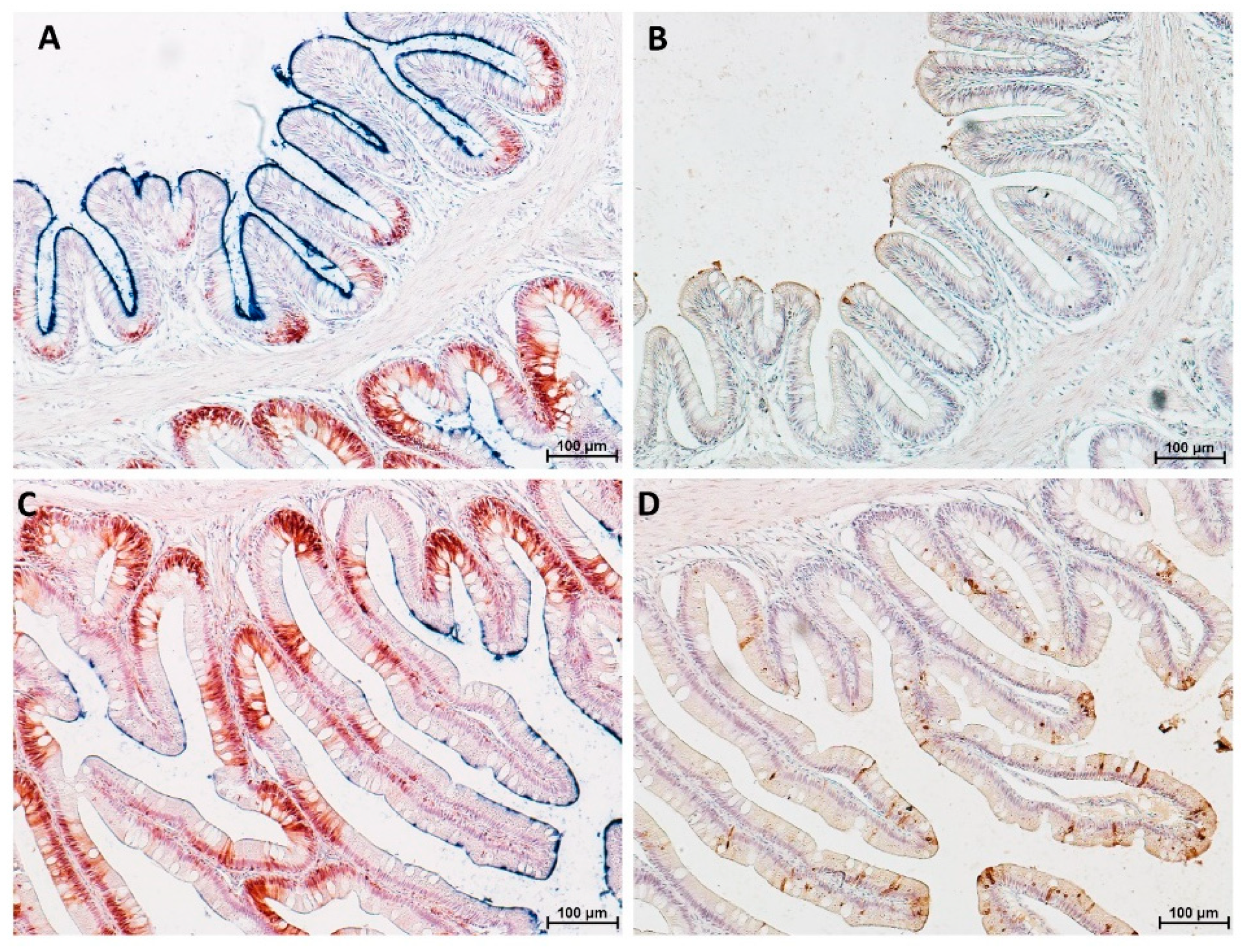
3.4. Goblet Cells
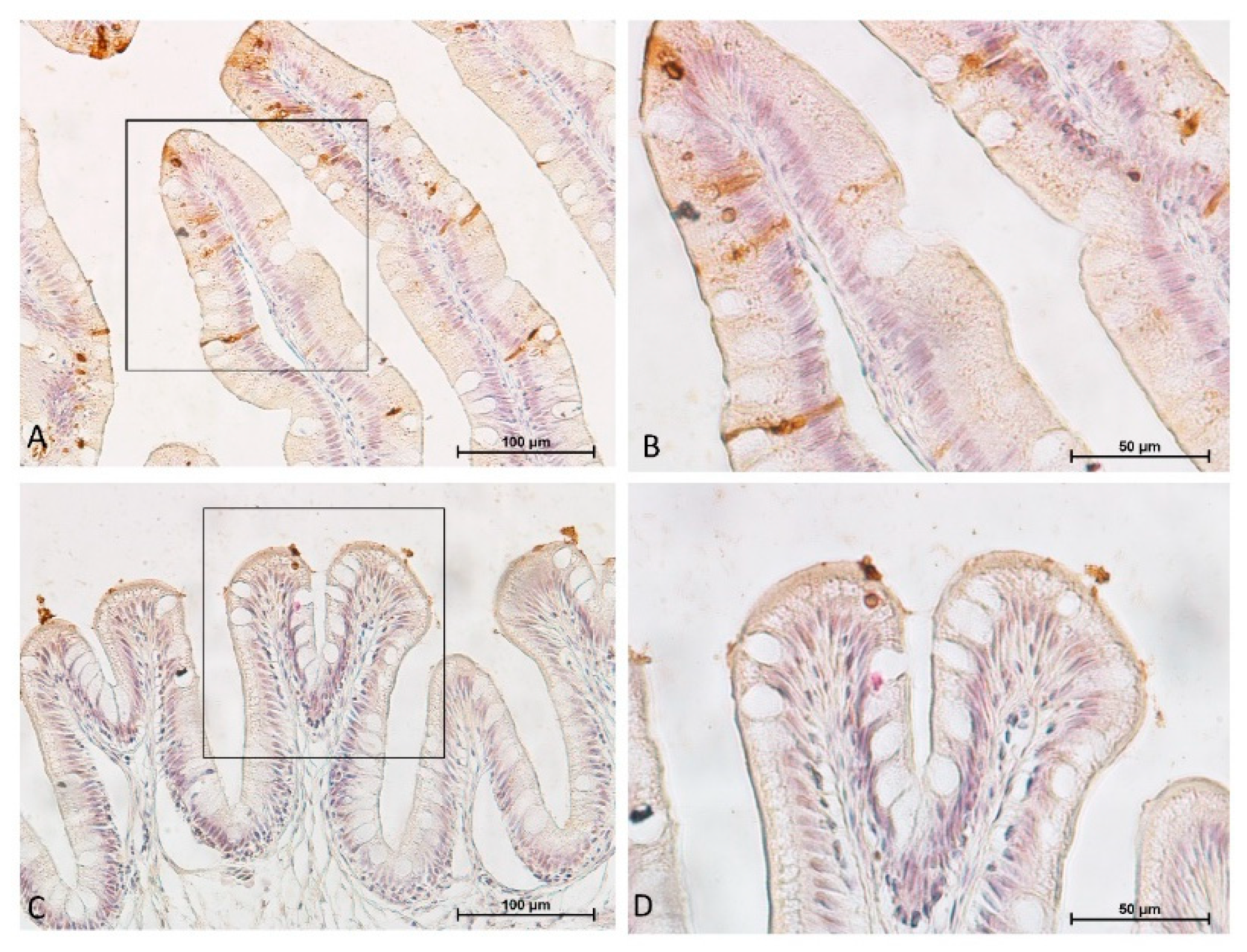
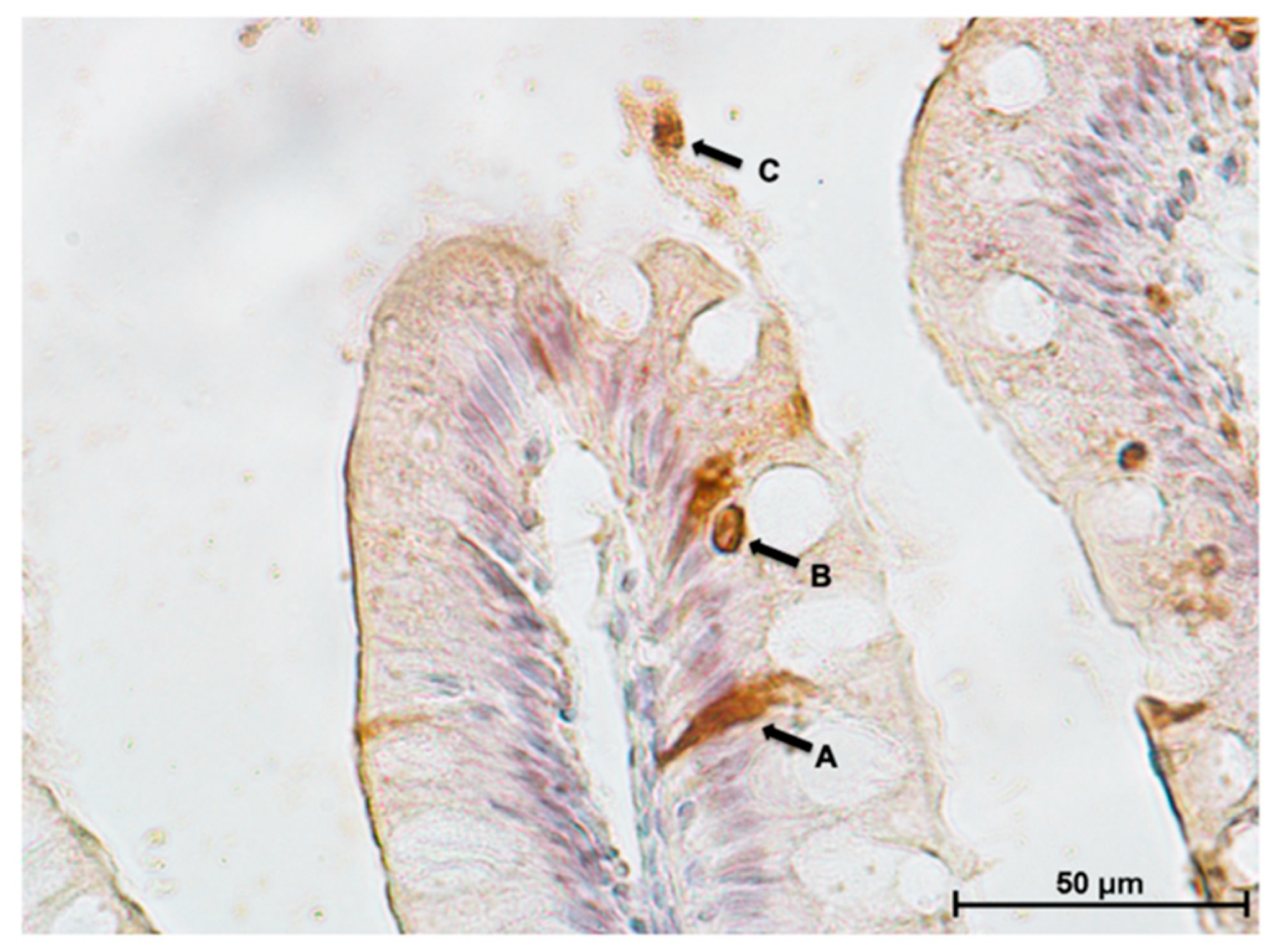
3.5. Intestinal Eosinophilic Granule Cells
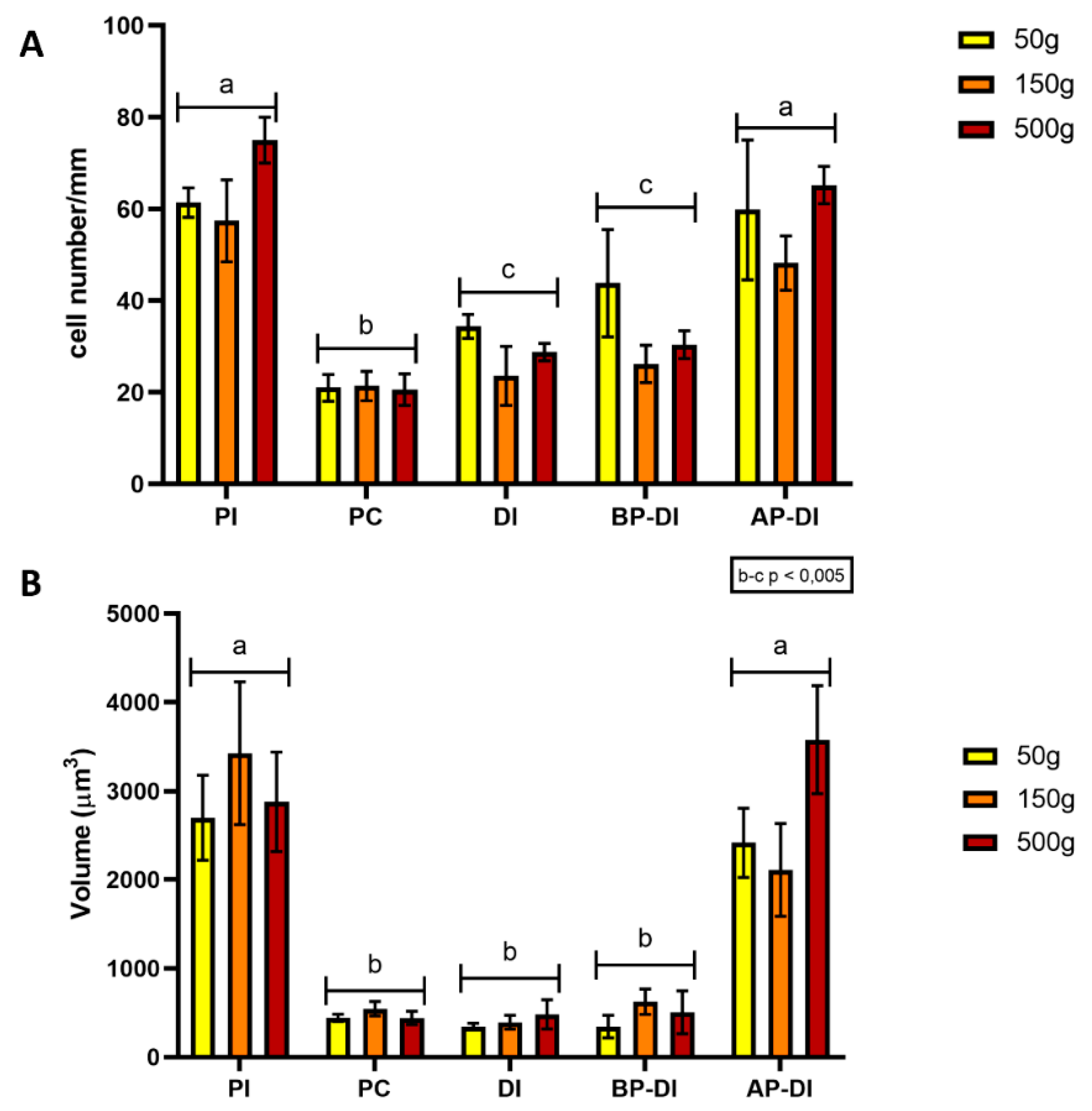
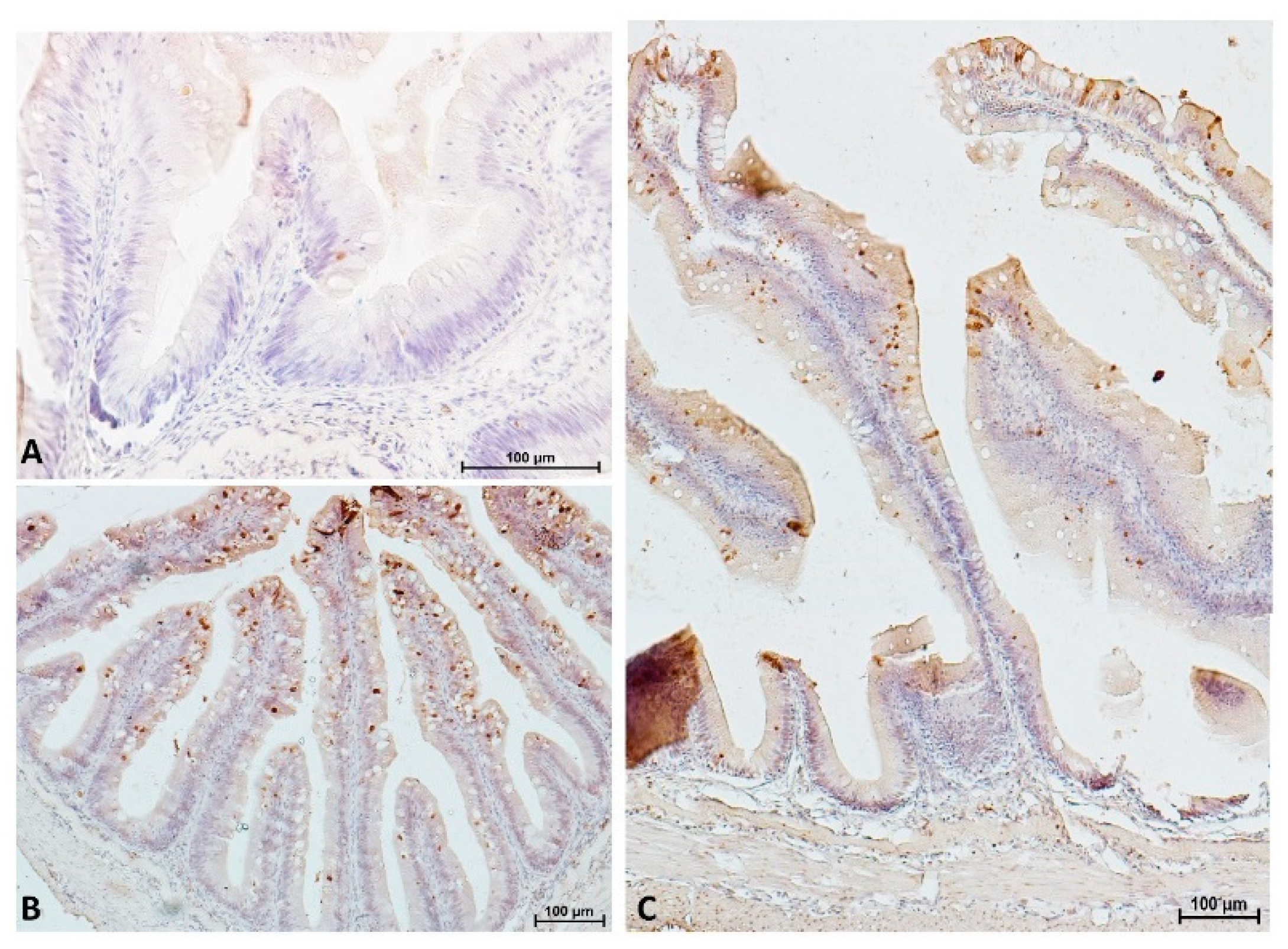
3.6. Proliferation, Differentiation and Apoptosis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Crawford, S.S.; Muir, A.M. Global introductions of salmon and trout in the genus oncorhynchus: 1870–2007. Rev. Fish Biol. Fish. 2008, 18, 313–344. [Google Scholar] [CrossRef]
- Caimi, C.; Gasco, L.; Biasato, I.; Malfatto, V.; Varello, K.; Prearo, M.; Pastorino, P.; Bona, M.C.; Francese, D.R.; Schiavone, A.; et al. Could dietary black soldier fly meal inclusion affect the liver and intestinal histological traits and the oxidative stress biomarkers of Siberian sturgeon (Acipenser baerii) juveniles? Animals 2020, 10, 155. [Google Scholar] [CrossRef] [PubMed]
- Barker, N.; van Oudenaarden, A.; Clevers, H. Identifying the stem cell of the intestinal crypt: Strategies and pitfalls. Cell Stem Cells 2012, 11, 452–460. [Google Scholar] [CrossRef] [PubMed]
- Elia, A.C.; Capucchio, M.T.; Caldaroni, B.; Magara, G.; Dörr, A.J.M.; Biasato, I.; Biasibetti, E.; Righetti, M.; Pastorino, P.; Prearo, M.; et al. Influence of hermetia illucens meal dietary inclusion on the histological traits, gut mucin composition and the oxidative stress biomarkers in rainbow trout (Oncorhynchus mykiss). Aquaculture 2018, 496, 50–57. [Google Scholar] [CrossRef]
- Verdile, N.; Mirmahmoudi, R.; Brevini, T.A.L.; Gandolfi, F. Evolution of pig intestinal stem cells from birth to weaning. Animal 2019, 3, 1319. [Google Scholar] [CrossRef] [PubMed]
- Fagundes, K.R.C.; Rotundo, M.M.; Mari, R.B. Morphological and histochemical characterization of the digestive tract of the puffer fish Sphoeroides testudineus (Linnaeus 1758) (Tetraodontiformes: Tetraodontidae). An. Acad. Bras. Ciênc. 2016, 88, 1615–1624. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.K.; Ringø, E. The gastrointestinal tract of fish. Aquac. Nutr. 2014, 41, 1–13. [Google Scholar] [CrossRef]
- Khojasteh, S.M.B.; Sheikhzadeh, F.; Mohammadnejad, D.; Azami, A. Histological, histochemical and ultrastructural study of the intestine of rainbow trout (Oncorhynchus mykiss). World Appl. Sci. J. 2009, 6, 1525–1531. [Google Scholar]
- Li, Y.; Kortner, T.M.; Chikwati, E.M.; Munang’andu, H.M.; Lock, E.J.; Krogdahl, Å. Gut health and vaccination response in pre-smolt Atlantic salmon (Salmo salar) fed black soldier fly (Hermetia illucens) larvae meal. Fish Shellfish Immunol. 2019, 86, 1106–1113. [Google Scholar] [CrossRef]
- Hamidian, G.; Zirak, K.; Sheikhzadeh, N.; Oushani, K.A.; Shabanzadeh, S.; Divband, B. Intestinal histology and stereology in rainbow trout (Oncorhynchus mykiss) administrated with nanochitosan/zeolite and chitosan/zeolite composites. Aquac. Res. 2018, 49, 1803–1815. [Google Scholar] [CrossRef]
- Dall’aglio, C.; Mercati, F.; Faeti, V.; Acuti, G.; Trabalza-Marinucci, M.; de Felice, E.; Tardella, F.M.; Franciosini, M.P.; Casagrande Proietti, P.; Catorci, D.; et al. Immuno-and glyco-histochemistry as a tool to evaluate the oregano supplemented feed effects in pig gut. Eur. J. Histochem. 2020, 64, 3110. [Google Scholar] [CrossRef]
- Mercati, F.; Dall’Aglio, C.; Acuti, G.; Faeti, V.; Tardella, F.M.; Pirino, C.; de Felice, E.; Scocco, P. Oregano feed supplementation affects glycoconjugates production in swine gut. Animals 2020, 10, 149. [Google Scholar] [CrossRef] [PubMed]
- Albl, B.; Haesner, S.; Braun-Reichhart, C.; Streckel, E.; Renner, S.; Seeliger, F.; Wolf, E.; Wanke, R.; Blutke, A. Tissue sampling guides for porcine biomedical models. Toxicol. Pathol. 2016, 44, 414–420. [Google Scholar] [CrossRef]
- Nyengaard, J.R. Stereologic methods and their application in kidney research. J. Am. Soc. Nephrol. 1999, 10, 1100–1123. [Google Scholar]
- Burnstock, G. The morphology of the gut of the brown trout (Salmo trutta). J. Cell Sci. 1959, 3, 183–198. [Google Scholar]
- Lkka, G.; Austb, L.; Falk, K.; Bjerkås, I.; Koppang, E.O. Intestinal morphology of the wild atlantic salmon (Salmo salar). J. Morphol. 2013, 274, 859–876. [Google Scholar] [CrossRef] [PubMed]
- Hassanpour, M.; Joss, J. Anatomy and histology of the spiral valve intestine in juvenile australian lungfish, neoceratodus forsteri. Open Zool. J. 2009, 2, 62–85. [Google Scholar] [CrossRef]
- Henningsen, A.D.; Whitaker, B.R.; Walker, I.D. Protrusion of the valvular intestine in captive smalltooth sawfish and comments on pristid gastrointestinal anatomy and intestinal valve types. J. Aquat. Anim. Health 2005, 17, 289–295. [Google Scholar] [CrossRef]
- Çinar, K.; Şenol, N. Histological and histochemical characterization of the mucosa of the digestive tract in flower fish (Pseudophoxinus antalyae). J. Vet. Med. Ser. C Anatom. Histol. Embryol. 2006, 35, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Argyriou, T.; Clauss, M.; Maxwell, E.E.; Furrer, H.; Sánchez-Villagra, M.R. Exceptional preservation reveals gastrointestinal anatomy and evolution in early actinopterygian fishes. Sci. Rep. 2016, 6, 18758. [Google Scholar] [CrossRef]
- Glover, C.N.; Petri, D.; Tollefsen, K.E.; Jørum, N.; Handy, R.D.; Berntssen, M.H.G. Assessing the sensitivity of Atlantic salmon (Salmo salar) to dietary endosulfan exposure using tissue biochemistry and histology. Aquat. Toxicol. 2007, 84, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, G.F.; Landis, S.C. The relationship of ultrastructural and cytochemical features to absorptive activity in the goldfish intestine. Anat. Rec. 1972, 172, 675–701. [Google Scholar] [CrossRef]
- Arman, S.; Üçüncü, S.I. Histochemical characterization of convict cichlid (Amatitlania nigrofasciata) intestinal goblet cells. Pak. J. Zool. 2017, 49, 445–453. [Google Scholar] [CrossRef]
- Bosi, G.; Arrighi, S.; di Giancamillo, A.; Domeneghini, C. Histochemistry of glycoconjugates in mucous cells of Salmo trutta uninfected and naturally parasitized with intestinal helminths. Dis. Aquat. Organ. 2005, 64, 45–51. [Google Scholar] [CrossRef]
- Kim, J.J.; Khan, W.I. Goblet cells and mucins: Role in innate defense in enteric infections. Pathogens 2013, 2, 55–70. [Google Scholar] [CrossRef] [PubMed]
- Sveinbjornsson, B.; Olsen, R.; Paulsen, S. Immunocytochemical localization of lysozyme in intestinal eosinophilic granule cells (EGCs) of Atlantic salmon, Salmo salar L. J. Fish Dis. 1996, 19, 349–355. [Google Scholar] [CrossRef]
- Clevers, H.C.; Bevins, C.L. Paneth cells: Maestros of the small intestinal crypts. Annu. Rev. Physiol. 2013, 75, 289–311. [Google Scholar] [CrossRef]
- Krystel-Whittemore, M.; Dileepan, K.N.; Wood, J.G. Mast cell: A multi-functional master cell. Front. Immunol. 2016, 6, 620. [Google Scholar] [CrossRef]
- Reite, O.B.; Evensen, Ø. Inflammatory cells of teleostean fish: A review focusing on mast cells/eosinophilic granule cells and rodlet cells. Fish Shellfish Immunol. 2006, 20, 192–208. [Google Scholar] [CrossRef]
- Dezfuli, B.S.; Giari, L.; Lui, A.; Squerzanti, S.; Castaldelli, G.; Shinn, A.P.; Manera, M.; Lorenzoni, M. Proliferative cell nuclear antigen (PCNA) expression in the intestine of Salmo trutta trutta naturally infected with an acanthocephalan. Parasites Vectors 2012, 5, 198. [Google Scholar] [CrossRef]
- Lallès, J.P. Intestinal alkaline phosphatase: Novel functions and protective effects. Nutr. Rev. 2014, 72, 82–94. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.; Carr, A.; Corner, G.A.; Tögel, L.; Dávaos-Salas, M.; Tran, H.; Chueh, A.C.; Al-Obaidi, S.; Chionh, F.; Naseem, A.; et al. The intestinal epithelial cell differentiation marker intestinal alkaline phosphatase (ALPi) is selectively induced by histone deacetylase inhibitors (HDACi) in colon cancer cells in a kruppel-like factor 5 (KLF5)-defendent manner. J. Biol. Chem. 2014, 289, 25306–25316. [Google Scholar] [CrossRef] [PubMed]
- Moss, S.F.; Attia, L.; Scholes, J.V.; Walters, J.R.F.; Holt, P.R. Increased small intestinal apoptosis in coeliac disease. Gut 1996, 39, 811–817. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; George, S.P.; Roy, S.; Pham, E.; Esmaeilniakooshkghazi, A.; Khurana, S. Both the anti-and pro-apoptotic functions of villin regulate cell turnover and intestinal homeostasis. Sci. Rep. 2016, 6, 35491. [Google Scholar] [CrossRef] [PubMed]




















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trout Weight (g) | N. | Rainbow Trout Length (cm) | Whole Intestinal Length (cm) | Intestinal Segment Length (cm) | |
|---|---|---|---|---|---|
| Proximal | Distal | ||||
| 50 | 5 | 16.8 ± 0.29 | 9.7 ± 0.62 | 6.0 ± 0.30 | 3.6 ± 0.28 |
| 150 | 5 | 22.7 ± 0.58 | 15.1 ± 0.80 | 9.0 ± 0.30 | 6.0 ± 0.50 |
| 500 | 5 | 32.4 ± 0.53 | 26.1 ± 0.76 | 18.2 ± 0.64 | 7.9 ± 0.11 |
| Trout Weight | N. | Pyloric Caeca (µm) | Enterocytes Supranuclear Vacuolization (SNV) |
|---|---|---|---|
| 50 g | 5 | 380.91 a ± 35.85 | − |
| 150 g | 5 | 719.50 b ± 32.45 | − |
| 500 g | 5 | 1011.50 c ± 18.14 | + |
| Trout Weight (g) | Proximal Intestine | ||||
|---|---|---|---|---|---|
| N. | Short Villi Length (µm) | Long Villi Length (µm) | Villi Width (µm) | Villi Branching | |
| 50 | 5 | 251.0 a ± 13.5 | 470.4 a ± 64.6 | 96.0 a ± 11.4 | − |
| 150 | 5 | 310.3 a ± 44.5 | 555.8 a ± 64.2 | 122.9 b ± 10.1 | + |
| 500 | 5 | 250.3 a ± 42.1 | 657.8 b ± 49.1 | 116.9 b ± 14,4 | ++ |
| Distal Intestine | ||||
|---|---|---|---|---|
| Trout Weight (g) | N. | Short Villi Length (µm) | Long Villi Length (µm) | Folds Height (µm) |
| 50 | 5 | − | − | − |
| 150 | 5 | 211.1 ± 3.1 | 592.4 ± 19.3 | 1284.9 ± 51.0 |
| 500 | 5 | 202.4 ± 20.1 | 556.5 ± 37.0 | 1410.1 ± 201.7 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Verdile, N.; Pasquariello, R.; Scolari, M.; Scirè, G.; Brevini, T.A.L.; Gandolfi, F. A Detailed Study of Rainbow Trout (Onchorhynchus mykiss) Intestine Revealed That Digestive and Absorptive Functions Are Not Linearly Distributed along Its Length. Animals 2020, 10, 745. https://doi.org/10.3390/ani10040745
Verdile N, Pasquariello R, Scolari M, Scirè G, Brevini TAL, Gandolfi F. A Detailed Study of Rainbow Trout (Onchorhynchus mykiss) Intestine Revealed That Digestive and Absorptive Functions Are Not Linearly Distributed along Its Length. Animals. 2020; 10(4):745. https://doi.org/10.3390/ani10040745
Chicago/Turabian StyleVerdile, Nicole, Rolando Pasquariello, Marco Scolari, Giulia Scirè, Tiziana A. L. Brevini, and Fulvio Gandolfi. 2020. "A Detailed Study of Rainbow Trout (Onchorhynchus mykiss) Intestine Revealed That Digestive and Absorptive Functions Are Not Linearly Distributed along Its Length" Animals 10, no. 4: 745. https://doi.org/10.3390/ani10040745
APA StyleVerdile, N., Pasquariello, R., Scolari, M., Scirè, G., Brevini, T. A. L., & Gandolfi, F. (2020). A Detailed Study of Rainbow Trout (Onchorhynchus mykiss) Intestine Revealed That Digestive and Absorptive Functions Are Not Linearly Distributed along Its Length. Animals, 10(4), 745. https://doi.org/10.3390/ani10040745