1. Introduction
The sensory systems of animals are crucial to detect environmental cues, and they are then processed through the nervous system to generate appropriate behaviours [
1], such as finding food, avoiding predators, identifying conspecifics, locating suitable habitat, and attracting mates [
2,
3,
4]. In aquatic systems, as on land, chemical cues affect not only individual behaviour and population dynamics, but also community organisation and ecosystem function.
Use of sensory modalities may be related to the ecology of the species, as prey or predator. Animals use different sensory modalities to search for food such as chemical, vibrational, tactile, sound, heat, and visual senses [
5]. Among them, while vision enables marine animals to swim directly to food items when they see it, chemoreception is essential to detect and locate food items, especially for animals active at night or in the deep ocean [
6,
7,
8]. Several studies suggest that aquatic species rely more strongly on chemical perception rather than vision one when discriminating between harmless and dangerous heterospecifics [
9,
10,
11,
12]. On the other hand, aquatic environments are particularly prone to the variability of the visual and chemical conditions. For example, turbidity could reduce the efficacy of visual cues even if octopus is capable of polarised vision [
13,
14,
15], whereas currents may disrupt chemical information.
Accordingly, the sensory system’s capabilities in cephalopods have been inextricably associated with their evolutionary success, allowing them to occupy many ecological niches of the sea from shallow waters to the deep sea. Coleoids are endowed with a highly sophisticated nervous system [
16,
17,
18] and an exceptionally large brain that includes more than 30 differentiated lobes [
17,
19]. Among them,
Octopus vulgaris (hereafter octopus)
, well known as an intelligent soft-bodied animal, has a significant number of lobes of the nervous system dedicated to visual, tactile, and chemosensory perception [
20,
21]. Its nervous system has a high degree of cross-connectivity [
17,
20,
22] able to integrate sensory inputs coming from the environment through its well-developed sensory organs [
17,
23,
24,
25].
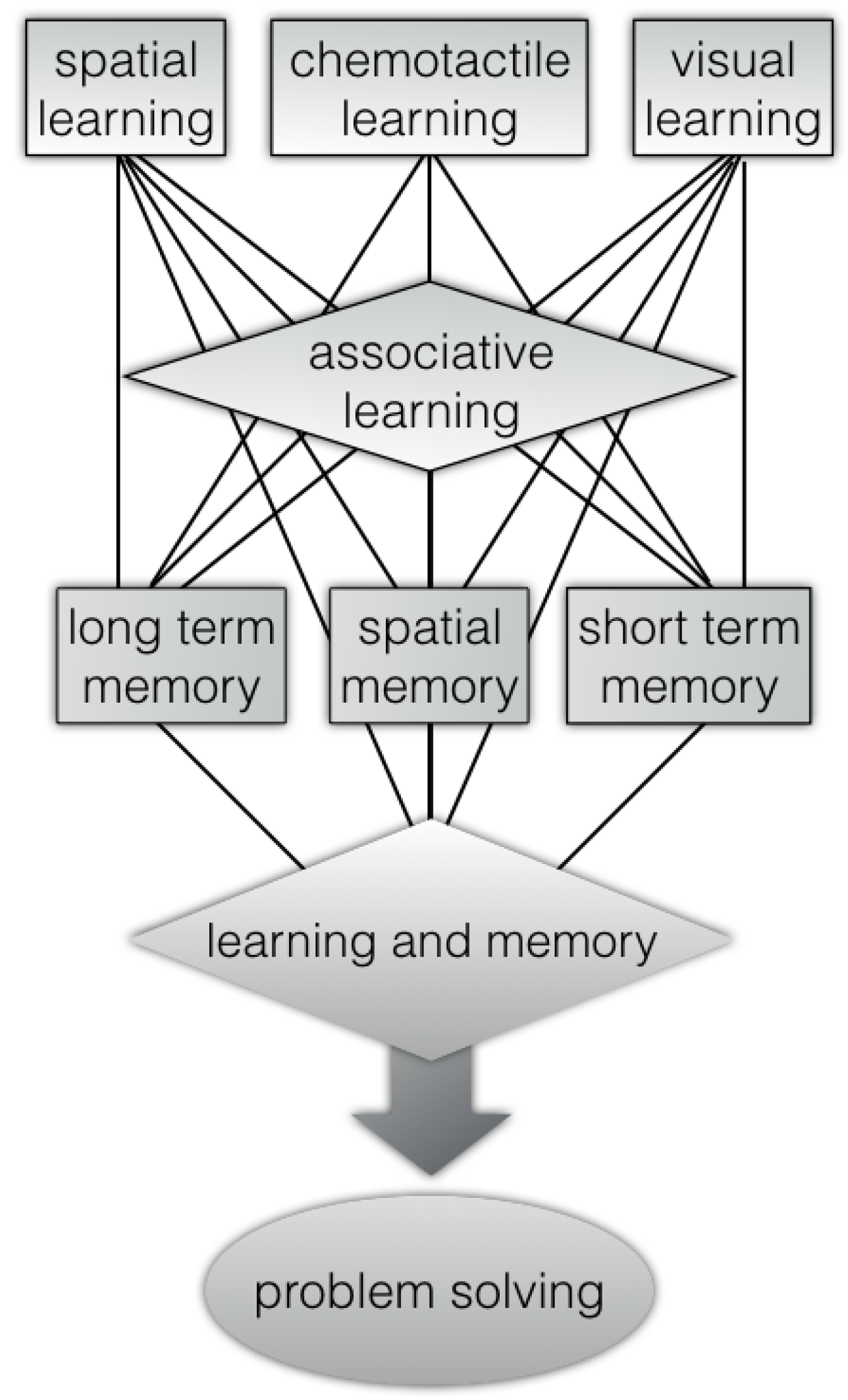
Indeed, octopus has a rich repertoire of complex behaviours (
Figure 1) that includes problem-solving, visual, and chemo-tactile.
In particular, the abilities of coleoids to perceive environmental cues have been mainly attributed to its visual systems. Although, under limited light conditions, the chemical signals are the primary important source as sensory inputs [
23,
55,
56,
57]. Thus, coleoids have remarkable abilities to recognise chemical cues through the buccal lips and mouth [
58], isolated sensory neurons [
59,
60], and arm suckers [
25,
61,
62,
63]. Thus, they may explore their environment by touch and taste, while their olfactory organs are able to perceive at distance [
23,
55,
64,
65,
66,
67,
68,
69], sensing a broad spectrum of chemical signals [
22].
It has been reported that chemosensory cues are important in decision-making in octopuses [
22,
65,
66,
67,
68,
69,
70,
71,
72,
73]. Training experiments for testing chemical discrimination have been done in octopus to demonstrate its ability to distinguish between objects based on their chemical differences using their arm suckers and described this ability as taste by touch [
24,
25], while odour discrimination was tested to assess perceptions of water-born chemical stimuli at distance [
72]. Furthermore, it has been highlighted that the octopus’s olfactory organ is able to change shape, from relaxed to erect to perceive water-soluble compounds such as salts, sugars, amino acids, amines, peptides, proteins, and functionalised hydrocarbons, which allows the animal to orient itself to detect the spatial gradient of these chemical cues, helping in navigation and triggering spatial memories [
23,
57,
74,
75]. Octopuses also possess a self-recognition mechanism, which consists of the attachment reflex inhibition of their own suckers, due to chemical signals in the skin [
76]. Recently, it has been hypothesised that olfaction in octopus is not restricted to the olfactory organ, but it is also extended to other structures such as the suckers, that were traditionally not considered olfactive. In particular, octopus exhibits a peculiar performance that can be defined “smell by touch”, useful to detect odorant molecules that in water are insoluble or have a very low solubility [
55,
74,
77,
78].
However, the octopus has always been described as a predominately “visual” animal with a complex visual system characterised by the presence of highly developed eyes [
22,
79,
80,
81,
82]. Analogously to vertebrate, octopus eyes are equipped with an un-inverted retina, a cornea, an iris, and a lens. Even if they have just one type of receptor cell and only rhodopsin as pigment, octopuses have the ability to recognise the plane of polarised light based on rhabdomeres dichroism. Moreover, it has been proposed that they are able to discriminate colours [
15] within a wide range of light conditions [
79,
80,
81], even if this mechanism is largely discussed due to the turbid aquatic environment and it should be confirmed by behavioural experiments [
83,
84].
Besides their eyes, octopus can detect light to trigger the animal’s colour changes using other visual senses. In fact, they can even perceive light through the skins [
85], and they can camouflage with the high-fidelity colour to natural and artificial backgrounds [
86,
87,
88,
89,
90,
91].
Experiments for testing visual discrimination have been established in octopus [
92]. For example, they can quickly learn to visually discriminate between a series of objects [
49,
51], learn to use vision to direct an arm to a target [
47], and recognise familiar conspecifics using vision [
48,
93]. Octopuses could be visually oriented as well, learning to use visual cues to choose and memorise a den, and take the correct route to return to it [
48].
When both chemical and visual information is available, octopuses combine information from all sensory inputs that they perceive and then the animals can camouflage themselves, escape a predator, or chase prey in the wild, or open jars for food in captivity [
32,
94,
95]. This integration of several sensory inputs may occur at central and/or peripheral levels [
74,
96], but the relative contribution of each sense remains poorly understood.
Our study investigated the priority given to chemical vs. visual perception to establish the sensorial hierarchy in food choice by O. vulgaris.
4. Discussion
In the wild, octopuses are generalist and opportunistic predators that prey on a great variety of species [
100,
101]. There were strong differences in prey preference among individual octopuses and the prey choice could be varied according to several factors such as predation risks, interspecific competition, or local prey abundance [
102].
In captivity, octopuses have shown preferences for selected preys [
103,
104,
105], making their food choice by using their sophisticated sense organs. Among these, much attention has been given to vision [
22,
79,
80,
81,
82]. They also possess sensitive olfactory organs that they use to detect chemicals in the water [
23,
57,
106,
107]. However, they also possess suckers that have excellent tactile and chemical sensitivity to perceive chemicals by touch [
55,
62,
68]. Both chemical and visual information is elaborated and stored in specific brain lobes, located in the supra-oesophageal mass and optic lobes [
17,
92]. Nevertheless, no previous study has addressed the question on which sense has the priority in the food search and choice, although most of the behavioural studies, performed prior to this one, were focused on just their visual capabilities [
47,
51,
64,
108,
109,
110,
111,
112]. Chemical perception is undoubtedly the first sense that evolved, resulting in widespread sensory modality in all animals. The biological system generated an enormous number of receptors genes to detect and recognise chemicals, but chemoperception resulted largely underappreciated by scientists, even in light of sensory drive evolution theory [
113,
114].
Here, we establish the priority given to chemical versus visual perception in octopus’s food choice just using the ethological approach. Our experiments are performed on a small animal group made by four samples that allow us to investigate the octopus’s behaviour, in according with the 3Rs rules (reduction, refinement, replacement) as allowed by the Italian law (European Directive 2010/63 EU L276; the Italian DL. 3 April 2014, no. 26).
On the basis of the octopus’s exploratory behaviours observed during food preference test (FP), animals tested show a clear preference for the anchovies. However, the first food touch was not always consistent with the food preference (
Figure 3), exhibiting a peculiar exploratory behaviour when they approach a new environment [
80,
94,
103,
115,
116,
117,
118,
119]. Although the fact that octopuses did not touch the preferred food immediately is a clear sign that they cannot rely just on visual perception when they approach a prey, evidently, they need to acquire more information about what they see, using other senses like chemical and tactile, to understand the nature of what they are going to eat.
To this end, octopuses are equipped with arms containing a widespread chemotactic sensory system concentrated in the hundreds of suckers [
24,
54,
61,
63,
120,
121]. Thus, food choice in octopuses is driven by multiple sensory cues; nevertheless, a hierarchy in sensory perceptions could be hypothesised.
In our experiments, it was clear that they are mainly attracted by the physical presence of it, without recognising the preferred food by vision at distance. Subsequently, after a random first touch (
Figure 3), octopuses start an evaluation of the food using tactile and chemical senses. This allowed us to recognise a temporal hierarchy, where the octopus uses first visual, tactile, and chemical senses, in this order. Our observations are in agreement with previous studies in which it has been reported that octopuses are visually oriented towards a new given object, and then explored it with their arms [
103,
110,
122]. In this case, we are not able to define which sensory cues are dominant by which they arrive at a decision on food choice, because this behaviour represents the result of the integration of different sensory information coming from visual, tactile, and/or chemical systems sequentially to perform a suitable behaviour.
To understand whether there is a sensorial hierarchy between visual and chemical, to establish which is dominant in decision-making, we tested octopuses with five discrimination tasks. Problem-solving and flexible tool-use are considered hallmarks of cognitive abilities and intelligence [
123,
124]. In the wild, octopuses exhibit behavioural flexibilities in solving many kinds of problem. For instance, the giant octopus, while attached to a rock can use one of the arm tips to attract a seagull, then when the seagull gets close the sea surface and within the range of the animal’s arm, it grabs and draws the bird into the water (
https://youtu.be/LNwegprmtx8). In captivity, octopus also exhibits cognitive abilities in solving problems, when challenged with artificial tasks. Octopuses could retrieve L-shaped food containers from crevices, with or without visual access and independently from the spatial orientation of containers [
32] or learn how to unscrew a jar to reach the food [
95]. The data here discussed clearly show the ability of octopuses to open jars during all five discrimination tasks successfully.
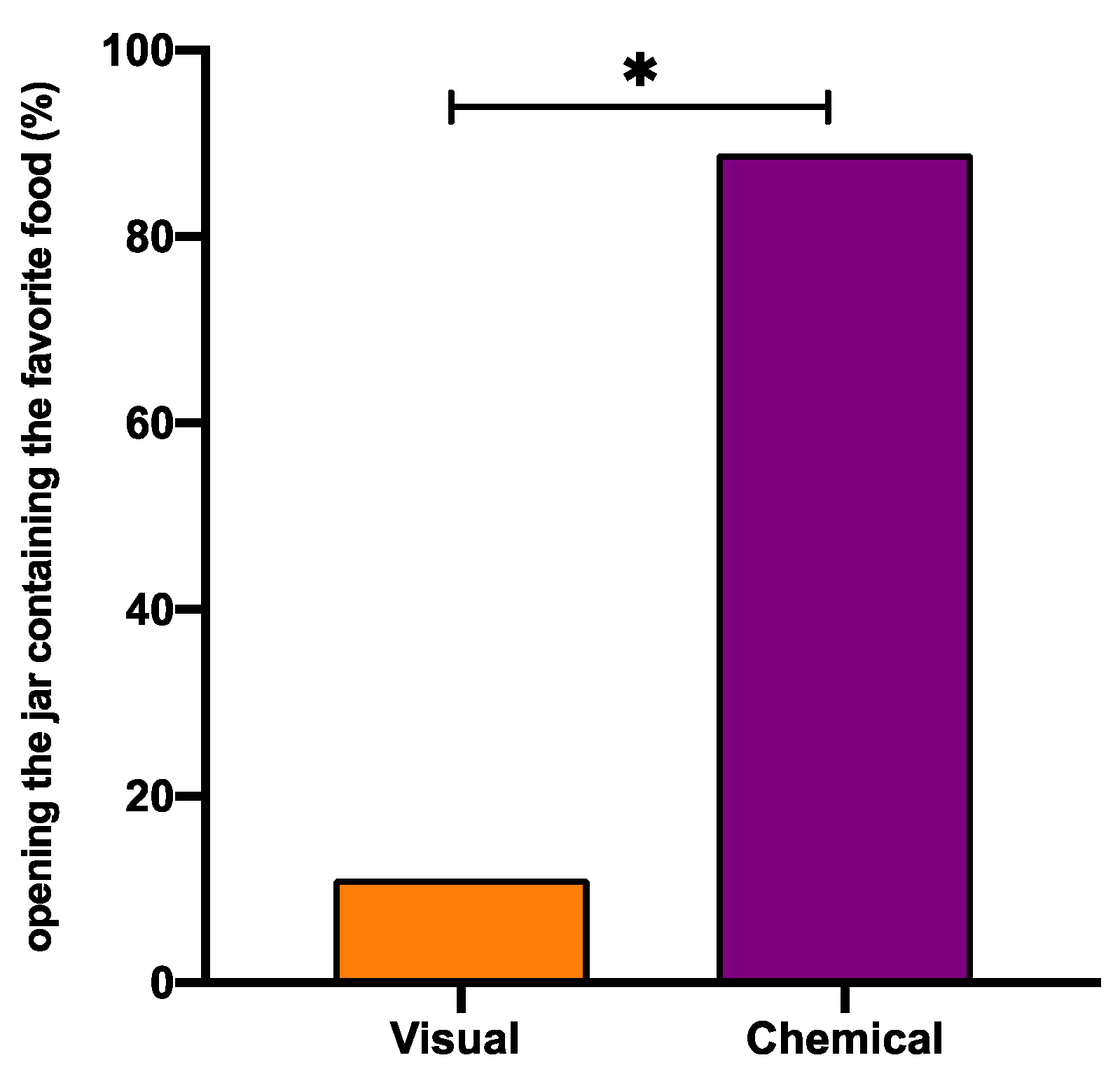
Our findings indicate that octopuses recognise the jar containing the anchovy, that resulted to be the preferred food in FP test (
Table 2 and
Table 3), in all discrimination tasks (T1–T4), with the exception of the negative control (T5) (
Table 4 and
Table 5). However, the task in which it is evident that the dominant sense is the chemical one, it is the confusion task (T4), where, despite the fact that octopuses were cheated with a false picture of the food inside, they picked up anchovy in the 100% of the cases. This evidence is corroborated when we excluded the chemical cues, focusing exclusively on the visual sense (T2), in which the jar containing anchovy was selected in only 50% of the cases. The negative control experiment (T5) reinforces our claim, in fact, that the chemical and visual information was not used by octopuses in solving this task, so the choice was randomly made.
These findings are consistent with Mather and collegues [
125] recently reporting that octopuses did not open the jar to get a small crayfish inside, because chemical cues from herring were smeared on its surface. Our experimental design (
Figure 2) allowed us to demonstrate that in
O. vulgaris both chemical and visual perceptions are essential in food choice; nevertheless, the chemical signals are the most important inputs. On the other hand, when we compared food eaten to jar choice under all different problem-solving tasks, we discovered that the preference to anchovies was maintained, even when the first jar chosen was not containing the anchovy (
Table 4 and
Table 5). In fact, in T2, where octopuses could just see the food inside, in 33.3% of cases, they opened first the jar containing a mussel, but because octopuses do not eat it, they resultantly were forced to look for another “chance to win” the preferred food. Merging the data coming from all tasks (
Table 4 and
Table 5), we observed that the percentage of successful decisions to open as first the jar contained the preferred food, based on chemical cues, were significantly higher than visual one (88.9% vs. 11.1% respectively, Wilcoxon matched-pairs test
p < 0.05).
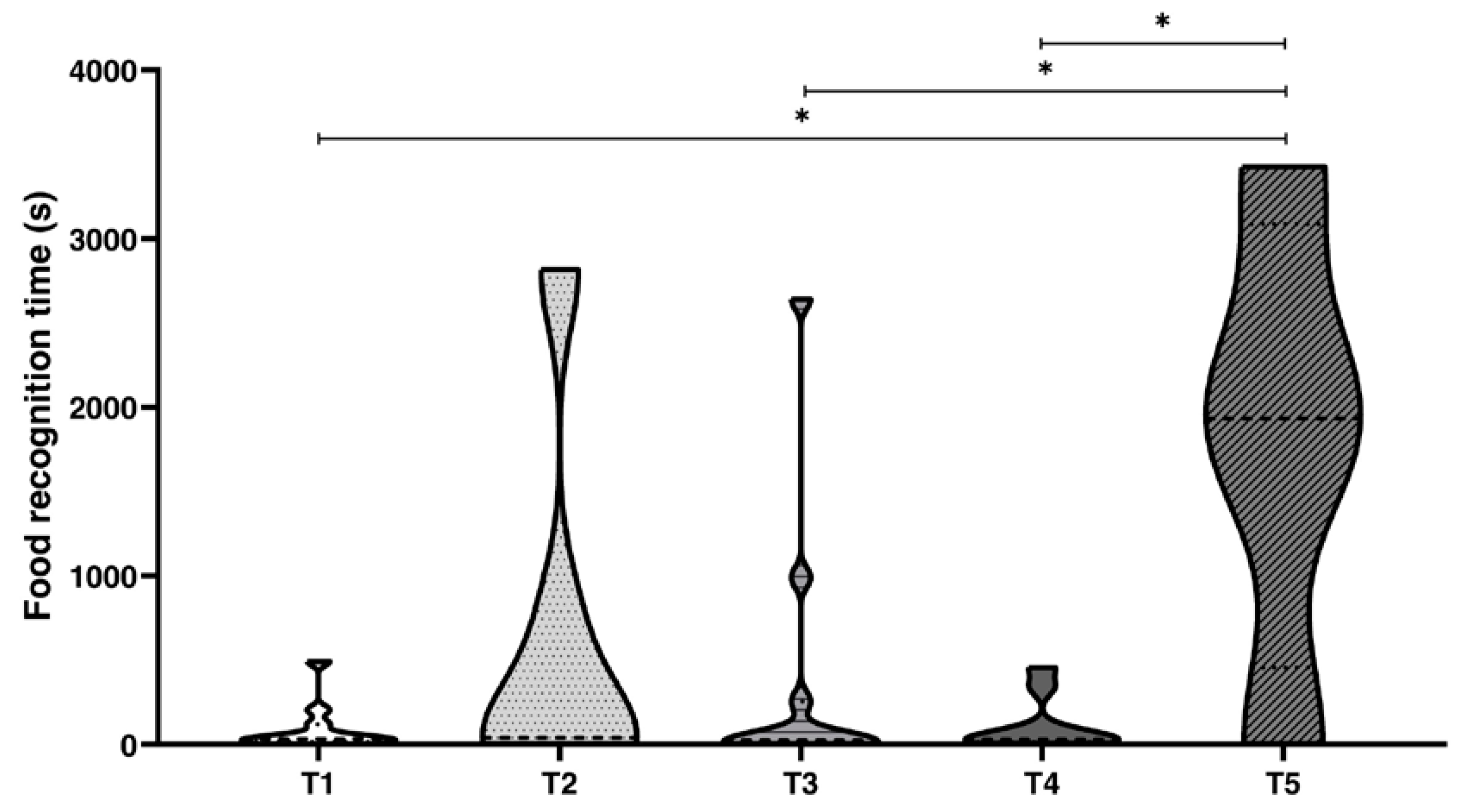
Furthermore, when we considered Δt, octopuses spent more time to visually discriminate the preferred food than either by combined visual and chemical discrimination or by chemical discrimination only (
Figure 4). Despite the fact that the differences encountered in Δt are affected by inter-individual variability, that is well known in this animal, and the limited number of specimens used, these results indicate that octopuses are able to decrease Δt to correctly solve operant tasks based on chemical information (
Figure 4), this might be of importance in predation strategy in the wild where the prey is not closed in a jar, but hidden and ready to escape. However, while we posit that octopus, privileges its chemical stimuli over visual ones, we should appreciate that the two combined increase the probability of success in prey [
74,
95,
96,
126].
Our results demonstrate involvement of chemosensory sense in octopus food choice behaviour, and allows a reassessment of the importance of chemical perception in the ecology of octopus. Octopus is, in fact, able to detect chemical cues with different spatial ranges through either contact or distant chemoreception. Its capability is based on the presence of the olfactory organ, structure mainly dedicated for hydrosoluble molecules, and the numerous chemoreceptors bared on its sucker rims that are essential to perceive and explore bi-dimensional traces consisting of insoluble molecules released on the seafloor in turn by preys, predators, or conspecifics [
74,
75]. These considerations will open new perspectives to study the behaviour of such an intriguing animal that is the octopus.

 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}