Black Soldier Fly Full-Fat Larvae Meal as an Alternative to Fish Meal and Fish Oil in Siberian Sturgeon Nutrition: The Effects on Physical Properties of the Feed, Animal Growth Performance, and Feed Acceptance and Utilization



Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Insect Meal and Diet Preparation
2.3. Feed Physical Properties
2.4. Animal Experiments
2.4.1. Experiment 1: Growth Performance and Feed Utilization
2.4.2. Experiment 2: Feed Acceptance Test
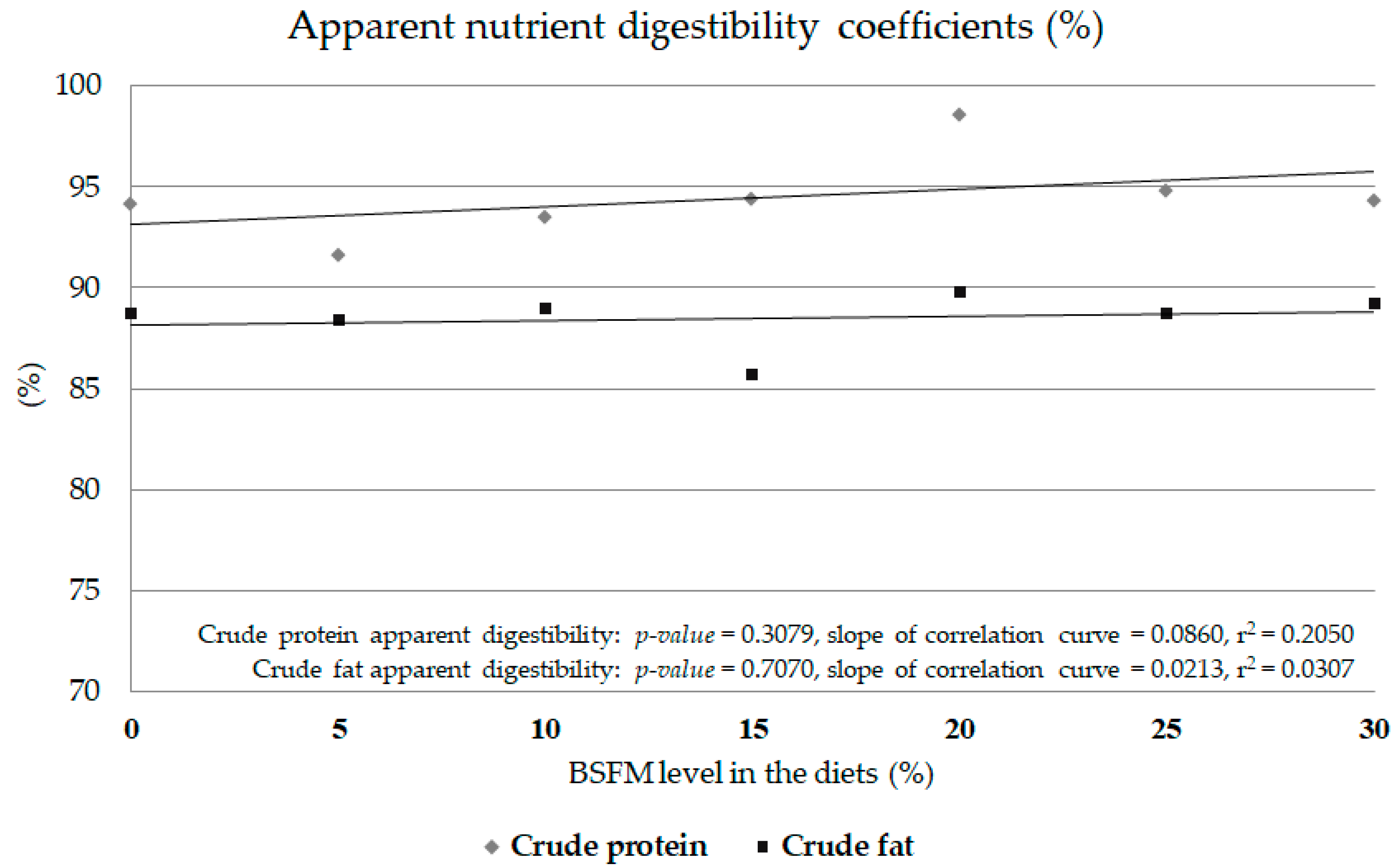
2.4.3. Experiment 3: Nutrient Digestibility Coefficient Assessment
2.5. Statistical Analysis
3. Results
3.1. Raw Materials
3.2. Feed Composition and Physical Properties
3.3. Growth Performance, Feed Utilization, and Acceptance
3.4. Digestibility Coefficients
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gasco, L.; Gai, F.; Maricchiolo, G.; Genovese, L.; Ragonese, S.; Bottari, T.; Caruso, G. Fishmeal alternative protein sources for aquaculture feeds. In Feeds for the Aquaculture Sector: Current Situation and Alternative Sources; Gasco, L., Gai, F., Maricchiolo, G., Genovese, L., Ragonese, S., Bottari, T., Caruso, G., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 1–28. [Google Scholar]
- Makkar, H.P.S.; Tran, G.; Heuzé, V.; Ankers, P. State-of-the-art on use of insects as animal feed. Anim. Feed Sci. Technol. 2014, 197, 1–33. [Google Scholar] [CrossRef]
- Nogales-Mérida, S.; Gobbi, P.; Józefiak, D.; Mazurkiewicz, J.; Dudek, K.; Rawski, M.; Kierończyk, B.; Józefiak, A. Insect meals in fish nutrition. Rev. Aquac. 2019, 11, 1080–1103. [Google Scholar] [CrossRef]
- Sicuro, B. The future of caviar production on the light of social changes: A new dawn for caviar? Rev. Aquac. 2019, 11, 204–219. [Google Scholar] [CrossRef]
- Athwal, N.; Wells, V.K.; Carrigan, M.; Henninger, C.E. Sustainable luxury marketing: A synthesis and research agenda. Int. J. Manag. Rev. 2019, 21, 405–426. [Google Scholar] [CrossRef]
- Hartmann, L.H.; Nitzko, S.; Spiller, A. Segmentation of German consumers based on perceived dimensions of luxury food. J. Food Prod. Mark. 2017, 23, 733–768. [Google Scholar] [CrossRef]
- Tacon, A.G.J. Feed ingredients for carnivorous fish species: Alternatives to fishmeal and other fishery resources. In Sustainable Fish Farming, Proceedings of the First International Symposium on Sustainable Fish Farming, Oslo, Norway, 28–31 August 1994; Reinertsen, H., Haaland, H., Eds.; A.A. Balkema: Rotterdam, The Netherlands; Brookfield, VT, USA, 1995; pp. 89–114. [Google Scholar]
- Lazzarotto, V.; Corraze, G.; Leprevost, A.; Quillet, E.; Dupont-Nivet, M.; Médale, F. Three-year breeding cycle of rainbow trout (Oncorhynchus mykiss) fed a plant-based diet, totally free of marine resources: Consequences for reproduction, fatty acid composition and progeny survival. PLoS ONE 2015, 10, e0117609. [Google Scholar] [CrossRef]
- Williot, P.; Nonnotte, G.; Chebanov, M. The Siberian Sturgeon (Acipenser baerii, Brandt, 1869); Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Gerrity, P.; Guy, C.; Gardner, W. Juvenile Pallid Sturgeon are Piscivorous: A Call for Conserving Native Cyprinids. Trans. Am. Fish. Soc. 2005, 135, 604–609. [Google Scholar] [CrossRef]
- Barroso, F.G.; de Haro, C.; Sánchez-Muros, M.J.; Venegas, E.; Martínez-Sánchez, A.; Pérez-Bañón, C. The potential of various insect species for use as food for fish. Aquaculture 2014, 422–423, 193–201. [Google Scholar] [CrossRef]
- Henry, M.; Gasco, L.; Piccolo, G.; Fountoulaki, E. Review on the use of insects in the diet of farmed fish: Past and future. Anim. Feed Sci. Technol. 2015, 203, 1–22. [Google Scholar] [CrossRef]
- Wang, Y.S.; Shelomi, M. Review of black soldier fly (Hermetia illucens) as animal feed and human food. Foods 2017, 6, 91. [Google Scholar] [CrossRef]
- Józefiak, A.; Nogales-Mérida, S.; Mikołajczak, Z.; Rawski, M.; Kierończyk, B.; Mazurkiewicz, J. The utilization of full-fat insect meal in rainbow trout (Oncorhynchus mykiss) nutrition: The effects on growth performance, intestinal microbiota and gastrointestinal tract histomorphology. Ann. Anim. Sci. 2019, 19, 747–765. [Google Scholar] [CrossRef]
- Józefiak, A.; Nogales-Mérida, S.; Rawski, M.; Kierończyk, B.; Mazurkiewicz, J. Effects of insect diets on the gastrointestinal tract health and growth performance of Siberian sturgeon (Acipenser baerii Brandt, 1869). BMC Vet. Res. 2019, 15, 348. [Google Scholar] [CrossRef] [PubMed]
- Caimi, C.; Renna, M.; Lussiana, C.; Bonaldo, A.; Gariglio, M.; Meneguz, M.; Dabbou, S.; Schiavone, A.; Gai, F.; Elia, A.C.; et al. First insights on black soldier fly (Hermetia illucens L.) larvae meal dietary administration in Siberian sturgeon (Acipenser baerii Brandt) juveniles. Aquaculture 2020, 515, 734539. [Google Scholar] [CrossRef]
- Caimi, C.; Gasco, L.; Biasato, I.; Malfatto, V.; Varello, K.; Prearo, M.; Pastorino, P.; Bona, M.C.; Francese, D.R.; Schiavone, A.; et al. Could dietary black soldier fly meal inclusion affect the liver and intestinal histological traits and oxidative stress biomarkers of Siberian sturgeon (Acipenser baerii) juveniles? Animals 2020, 10, 155. [Google Scholar] [CrossRef] [PubMed]
- Renna, M.; Schiavone, A.; Gai, F.; Dabbou, S.; Lussiana, C.; Malfatto, V.; Prearo, M.; Capucchio, M.; Biasato, I.; Biasebetti, E.; et al. Evaluation of the suitability of a partially defatted black soldier fly (Hermetia illucens L.) larvae meal as ingredient for rainbow trout (Oncorhynchus mykiss Walbaum diets. J. Anim. Sci. Biot. 2017, 8, 57. [Google Scholar] [CrossRef]
- Bruni, L.; Pastorelli, R.; Viti, C.; Gasco, L.; Parisi, G. Characterization of the intestinal microbial communities of rainbow trout (Oncorhynchus mykiss) fed with Hermetia illucens (black soldier fly) partially defatted larva meal as partial dietary protein source. Aquaculture 2018, 487, 56–63. [Google Scholar] [CrossRef]
- Wang, G.; Peng, K.; Hu, J.; Yi, C.; Chen, X.; Wu, H.; Huang, Y. Evaluation of defatted black soldier fly (Hermetia illucens L.) larvae meal as an alternative protein ingredient for juvenile Japanese seabass (Lateolabrax japonicus) diets. Aquaculture 2019, 507, 144–154. [Google Scholar] [CrossRef]
- Dumas, A.; Raggi, T.; Barkhouse, J.; Lewis, E.; Weltzen, E. The oil fraction and partially defatted meal of black soldier fly larvae (Hermetia illucens) affects differently growth performance, feed efficiency, nutrient deposition, blood glucose and lipid digestibility of rainbow trout (Oncorhynchus mykiss). Aquaculture 2018, 492, 24–34. [Google Scholar] [CrossRef]
- Janssen, R.H.; Vincken, J.P.; van den Broek, L.A.; Fogliano, V.; Lakemond, C.M. Nitrogen-to-protein conversion factors for three edible insects: Tenebrio molitor, Alphitobius diaperinus, and Hermetia illucens. J. Agric. Food Chem. 2017, 65, 2275–2278. [Google Scholar] [CrossRef]
- Yildiz, H.; Robaina, L.; Pirhonen, J.; Mente, E.; Dominguez, D.; Parisi, G. Fish welfare in aquaponics systems: Its relation to water quality with emphasis on feed and faeces—A review. Water 2017, 9, 13. [Google Scholar] [CrossRef]
- Topic Popovic, N.; Strunjak-Perovic, I.; Coz-Rakovac, R.; Barisic, J.; Jadan, M.; Persin Berakovic, A.; Sauerborn Klobucar, R. Tricaine methane-sulfonate (MS-222) application in fish anaesthesia. J. Appl. Ichthyol. 2012, 28, 553–564. [Google Scholar] [CrossRef]
- Leary, S.; Underwood, W.; Anthony, R.; Cartner, S. AVMA Guidelines for the Euthanasia of Animals: 2013 Edition; AVMA: Schaumburg, IL, USA, 2013; pp. 67–73. [Google Scholar]
- AOAC. Official Methods of Analysis of AOAC International; AOAC: Washington, DC, USA, 2005. [Google Scholar]
- Myers, W.D.; Ludden, P.A.; Nayigihugu, V.; Hess, B.W. Technical note: A procedure for the preparation and quantitative analysis of samples for titanium dioxide. J. Anim. Sci. 2004, 82, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Short, F.J.; Gorton, P.; Wiseman, J.; Boorman, K.N. Determination of titanium dioxide added as an inert marker in chicken digestibility studies. Anim. Feed Sci. Technol. 1996, 59, 215–221. [Google Scholar] [CrossRef]
- Kierończyk, B.; Sypniewski, J.; Rawski, M.; Czekala, W.; Swiatkiewicz, S.; Józefiak, D. From waste to sustainable feed material: The effect of Hermetia illucens oil on the growth performance, nutrient digestibility, and gastrointestinal tract morphometry of broiler chickens. Ann. Anim. Sci. 2020, 20, 157–177. [Google Scholar] [CrossRef]
- Sypniewski, J.; Kierończyk, B.; Benzertiha, A.; Mikołajczak, Z.; Pruszyńska-Oszmałek, E.; Kołodziejski, P.; Sassek, M.; Rawski, M.; Czekała, W.; Józefiak, D. Replacement of soybean oil by Hermetia illucens fatin turkey nutrition: Effect on performance, digestibility, microbial community, immune and physiological status and final product quality. Brit. Poult. Sci. 2020, 61, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Finke, M.D. Estimate of chitin in raw whole insects. Zoo. Biol. 2007, 26, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Khater, E.G.; Bahnasawy, A.H.; Ali, S.A. Physical and mechanical properties of fish feed pellets. J. Food Process. Technol. 2014, 5, 378. [Google Scholar]
- Irungu, F.; Mutungi, C.; Faraj, A.; Affognon, H.; Kibet, N.; Tanga, C.; Ekesi, S.; Nakimbugwe, D.; Fiaboe, K. Physico-chemical properties of extruded aquafeed pellets containing black soldier fly (Hermetia illucens) larvae and adult cricket (Acheta domesticus) meals. J. Insects Food Feed 2018, 4, 19–30. [Google Scholar] [CrossRef]
- Das, H.K.; Hattula, M.T.; Myllymäki, O.M.; Mälkki, Y. Effects of formulation and processing variables on dry fish feed pellets containing fish waste. J. Sci. Food Agric. 1993, 61, 181–187. [Google Scholar] [CrossRef]
- Umar, S.; Kamarudin, M.S.; Ramezani-Fard, E. Physical properties of extruded aquafeed with a combination of sago and tapioca starches at different moisture contents. Anim. Feed Sci. Technol. 2013, 183, 51–55. [Google Scholar] [CrossRef]
- Hung, S.S.O.; Lutes, P.B.; Shqueir, A.A.; Conte, F.S. Effect of feeding rate and water temperature on growth of juvenile white sturgeon (Acipenser transmontanus). Aquaculture 1993, 115, 297–303. [Google Scholar] [CrossRef]
- Hardy, R.W.; Barrows, F.T. Diet formulation and manufacture. In Fish Nutrition, 3rd ed.; Halver, J.E., Hardy, R.W., Eds.; Academic Press: San Diego, CA, USA, 2002; pp. 506–601. [Google Scholar]
- Allan, G.L.; Rowland, S.J.; Parkinson, S.; Stone, A.J.B.; Jantrarotai, W. Nutrient digestibility for juvenile Silver perch Bidyanus bidyanus development methods. Aquaculture 1999, 170, 131–145. [Google Scholar] [CrossRef]
- Kaczmarek, S.A.; Hejdysz, M.; Kubis, M.; Kasprowicz-Potocka, M.; Rutkowski, A. The nutritional value of yellow lupin (Lupinus luteus L.) for broilers. Anim. Feed Sci. Technol. 2016, 222, 43–53. [Google Scholar] [CrossRef]
- Rawski, M.; Kierończyk, B.; Długosz, J.; Świątkiewicz, S.; Józefiak, D. Dietary probiotics affect gastrointestinal tract microbiota, histological structure and shell mineralization in turtles. PLoS ONE 2016, 11, e1478859. [Google Scholar] [CrossRef]
- Józefiak, D.; Józefiak, A.; Kierończyk, D.; Rawski, M.; Świątkiewicz, S.; Długosz, J.; Engberg, R.M. Insects—A natural nutrient source for poultry—A review. Ann. Anim. Sci. 2016, 16, 297–313. [Google Scholar] [CrossRef]
- Hoffmann, L.; Rawski, M.; Nogales-Mérida, S.; Mazurkiewicz, J. Dietary inclusion of Tenebrio molitor meal in sea trout larvae rearing: Effects on fish growth performance, survival, condition, and git and liver enzymatic activity. Ann. Anim. Sci. 2020, 20, 579–598. [Google Scholar] [CrossRef]
- Mikołajczak, Z.; Rawski, M.; Mazurkiwicz, J.; Kierończyk, B.; Józefiak, D. The effects of hydrolyzed insect meals in sea trout fingerling (Salmo trutta m. trutta) diets on growth performance, microbiota and biochemical blood parameters. Animals 2020, 10, 1031. [Google Scholar] [CrossRef]
- Hung, S.S.O. Recent advances in sturgeon nutrition. Anim. Nutr. 2017, 3, 191–204. [Google Scholar] [CrossRef]
- Guo, Z.; Zhu, X.; Liu, J.; Han, D.; Yang, Y.; Lan, Z.; Xie, S. Effects of dietary protein level on growth performance, nitrogen and energu budget of juvenile hybrid sturgeon Acipenser baerii × A. gueldenstaedtii. Aquaculture 2012, 338, 89–95. [Google Scholar] [CrossRef]
- Xu, Q.Y.; Xu, H.; Wang, C.; Zheng, Q.; Sun, D. Studies on dietary phosphorus requirement of juvenile Siberian sturgeon Acipenser baerii. J. Appl. Ichthyol. 2011, 27, 709–714. [Google Scholar] [CrossRef]
- Sugiura, S.H. Phosphorus, aquaculture and environment. Rev. Fish. Sci. Aquac. 2018, 28, 515–521. [Google Scholar] [CrossRef]
- Ilo, S.; Schoenlechner, R.; Berghofe, E. Role of lipids in the extrusion cooking processes. Grasas Aceites 2000, 51, 97–110. [Google Scholar] [CrossRef]
- Weththasinghe, P.; Hansen, J.; Nøkland, D.; Lagos, L.; Rawski, M.; Øverland, M. Full-fat black soldier fly larvae (Hermetia illucens) meal and paste in extruded diets for Atlantic salmon (Salmo salar): Effect on physical pellet quality, nutrient digestibility, nutrient utilization and growth performances. Aquaculture 2021, 530, 735785. [Google Scholar] [CrossRef]
- Faller, J.Y.; Heymann, H. Sensory and physical properties of extruded potato puffs. J. Sens. Stud. 1996, 11, 227–245. [Google Scholar] [CrossRef]
- Harper, J.M. The Technology of Extrusion Cooking; Frame, N., Ed.; Blackie Academic & Professional: London, UK, 1994. [Google Scholar]
- Murofushi, S.; Ina, K. Survey of feeding stimulants for the sea bream present in the dried pupae of silkworms. Agric. Biol. Chem. 1981, 45, 1501–1504. [Google Scholar] [CrossRef]
- Kasumyan, A.O.; Sidorov, S.S.; Kazhlaev, A.A.; Pashchenko, N.I. Behavioral responses of young stellate sturgeon to smell and taste of artificial feeds and their components. In Proceedings of the International Symposium on Stugeons, Moscow, Russia, 6–11 September 1993; Gershanovich, A.D., Smith, T.I.J., Eds.; VNIRO Publishing: Moscow, Russia, 1995; pp. 278–288. [Google Scholar]
- Kierończyk, B.; Rawski, M.; Pawełczyk, P.; Różyńska, J.; Golusik, J.; Mikołajczak, Z.; Józefiak, D. Do insects smell attractive to dogs? A comparison of dog reactions to insects and commercial feed aromas—A preelimnary study. Ann. Anim. Sci. 2018, 18, 795–800. [Google Scholar] [CrossRef]
- Kasumyan, A.O. Olfaction and Gustation in Acipenseridae, with special references to the Siberian sturgeon. In The Siberian sturgeon (Acipenser baerii, Brandt 1869) Volume 1—Biology, 1st ed.; Williot, P., Nonnotte, G., Vizziano-Cantonnet, D., Chebanov, D., Eds.; Springer: Cham, Switzerland, 2018; Volume 1, pp. 173–205. [Google Scholar]
- Kasumyan, A.O.; Taufik, L.R. Behavioral reaction of juvenile sturgeons (Acipenseridae) to amino acids. J. Ichthyol. 1994, 34, 90–103. [Google Scholar]
- Kanost, M. Haemolymph. In Encyclopedia of Insects, 2nd ed.; Resh, V., Carde, R., Eds.; Academic Press, An Elsevier Science Imprint: Cambridge, MA, USA, 2009; pp. 446–449s. [Google Scholar]
- Mintah, B.; He, E.; Dabbour, M.; Xiang, J.; Hui, J.; Agyekum, A.; Ma, H. Characterization of edible soldier fly protein and hydrolysate altered by multiple frequency ultrasound: Structural, physical, and functional attributes. Proc. Biochem. 2020, 95, 157–165. [Google Scholar] [CrossRef]
- Kasumyan, A.O. Taste attractiveness of free amino acids and their physiochemical and biological properties (as exemplified by fishes). J. Evol. Biochem. Physiol. 2016, 52, 271–281. [Google Scholar] [CrossRef]
- Park, S.; Kim, J.; Yoe, S. Purification and characterization of a novel antibacterial peptide from black soldier fly (Hermetia illucens) larvae. Dev. Comp. Immunol. 2015, 52, 98–106. [Google Scholar] [CrossRef]
- Moretta, A.; Salvia, R.; Scieuzo, C.; Di Somma, A.; Vogel, H.; Pucci, P.; Sgambato, A.; Wolff, M.; Falabella, P. A bioinformatic study on antimicrobial peptides identified in the black soldier fly (BSF) Hermetia illucens (Diptera: Stratiomyidae). Sci. Rep. 2020, 10, 16875. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Yun, E.; Goo, T. Antimicrobial activity of an extract of Hermetia illucens larvae immunized with Lactobacillus casei against Salmonella species. Insects 2020, 10, 704. [Google Scholar] [CrossRef] [PubMed]
- Gasco, L.; Finke, M.; Van Huis, A. Can diets containing insects promote animal health? J. Insects Food Feed 2018, 4, 1–4. [Google Scholar] [CrossRef]
- Belghit, I.; Liland, N.S.; Waagbø, R.; Biancarosa, I.; Pelusio, N.; Li, Y.; Krogdahl, Å.; Lock, E.J. Potential of insect-based diets for Atlantic salmon (Salmo salar). Aquaculture 2018, 491, 72–81. [Google Scholar] [CrossRef]
- Li, S.; Ji, H.; Zhang, B.; Tian, J.; Zhou, J.; Yu, H. Influence of black soldier fly (Hermetia illucens) larvae oil on growth performance in juvenile Jian Carp (Cyprinus carpio var. Jian). Aquaculture 2016, 465, 43–52. [Google Scholar] [CrossRef]
- Shiau, S.Y.; Yu, Y.P. Dietary supplementation of chitin and chitosan depresses growth in tilapia, Oreochromis niloticus × O. aureus. Aquaculture 1999, 179, 439–446. [Google Scholar] [CrossRef]
- Su, J.; Gong, Y.; Cao, S.; Lu, F.; Han, D.; Liu, H.; Jin, J.; Yang, Y.; Zhu, X.; Xie, S. Effects of dietary Tenebrio molitor meal on the growth performance, immune response and disease resistance of yellow catfish (Pelteobagrus fulvidraco). Fish Shellfish Immunol. 2017, 69, 59–66. [Google Scholar] [CrossRef]
- Volkman, E.T.; Pangle, K.L.; Rajchel, D.A.; Sutton, T.M. Hatchery performance attributes of juvenile lake sturgeon fed two natural food types. N. Am. J. Aquac. 2004, 66, 105–112. [Google Scholar] [CrossRef]


{kind=link}
| Nutrient- | BSFL | FM |
|---|---|---|
| g/1000 g of dry matter | ||
| Crude protein | 350 | 618 |
| Crude fat | 298 | 165 |
| Crude fibre | 79.0 | 0 |
| Crude ash | 53.0 | 175 |
| Nitrogen free extract | 221 | 42.0 |
| Amino acid | g/100 g of crude protein | |
| Aspartic acid | 7.30 | 9.40 |
| Glutamic acid | 13.1 | 14.5 |
| Serine | 4.88 | 4.17 |
| Glycine | 6.15 | 6.41 |
| Histidine | 3.25 | 2.09 |
| Arginine | 5.47 | 6.07 |
| Threonine | 4.43 | 4.10 |
| Alanine | 8.21 | 6.87 |
| Proline | 6.68 | 4.28 |
| Tyrosine | 6.71 | 3.00 |
| Valine | 6.79 | 5.79 |
| Methionine | 2.12 | 2.53 |
| Cystine | 0.76 | 9.59 |
| Isoleucine | 4.73 | 4.24 |
| Leucine | 7.83 | 7.48 |
| Phenylalanine | 7.76 | 3.07 |
| Lysine | 6.82 | 6.63 |
| Fatty Acid | BSFL | FM |
|---|---|---|
| g/100 g of crude fat | ||
| C12:0 (lauric acid) | 21.00 | <0.1 |
| C13:0 (tridecylic acid) | <0.1 | <0.1 |
| C14:0 (myristic acid) | 3.70 | 5.30 |
| C15:0 (pentadecanoic acid) | <0.1 | 1.10 |
| C16:0 (palmitic acid) | 5.30 | 5.30 |
| C17:0 (margaric acid) | <0.1 | 1.40 |
| C18:0 (stearic acid) | 0.80 | 4.40 |
| C20:0 (arachidic acid) | <0.1 | 0.20 |
| C21:0 (heneicosanoic acid) | <0.1 | 0.20 |
| C22:0 (behenic acid) | <0.1 | 0.20 |
| C23:0 (tricosylic acid) | <0.1 | 0.10 |
| C24:0 (lignoceric acid) | <0.1 | 0.20 |
| C14:1 (myristoleic acid) | <0.1 | 0.20 |
| C15:1 (ginkgolic acid) | <0.1 | <0.1 |
| C16:1 n7 (palmitoleic acid) | 1.60 | 5.30 |
| C17:1 (margaroleic acid) | <0.1 | <0.1 |
| C18:1 n9 (trans-elaidic acid) | <0.1 | 0.20 |
| C18:1 n9 (oleic acid) | 6.10 | 13.00 |
| C18:1 n7 (vaccenic acid) | 0.20 | <0.1 |
| C20:1 n9 (eicosenoic acid) | 0.10 | 1.50 |
| C22:1 n11 (gadoleic acid) | <0.1 | <0.1 |
| C22:1 n9 (erucic acid) | <0.1 | 0.20 |
| C24:1 n9 (nervonic acid) | <0.1 | <0.1 |
| C16:2 n4 (hexadecadienoic acid) | <0.1 | <.01 |
| C16:3 n4 (hexadecatrienoic acid) | <0.1 | <0.1 |
| C18:2 n6 (trans-linolelaidic acid) | <0.1 | 0.20 |
| C18:2 n6 (linoleic acid) | 3.80 | 1.70 |
| C18:3 n6 (γ-linolenic acid) | <0.1 | 0.10 |
| C18:3 n4 (octadecatrienoic acid) | <0.1 | <0.1 |
| C18:3 n3 (α-linolenic acid) | 0.50 | 1.00 |
| C18:4 n3 (stearidonic acid) | <0.1 | <0.1 |
| C20:2 n6 (eicosadienoic acid) | <0.1 | 0.30 |
| C20:3 n6 (dihomo-γ-linolenic acid) | <0.1 | <0.1 |
| C20:3 n3 (eicosatrienoic acid) | <0.1 | <0.1 |
| C20:4 n6 (arachidonic acid) | <0.1 | 1.20 |
| C20:4 n3 (eicosatetraenoic acid) | <0.1 | <0.1 |
| C20:5 n3 (eicosapentaenoic acid) | 0.20 | 7.90 |
| C22:2 n6 (docosadienoic acid) | <0.1 | 1.10 |
| C22:5 n3 (docosapentaenoic acid) | <0.1 | <0.1 |
| C22:6 n3 (docosahexaenoic acid) | <0.1 | 18.30 |
| Treatment | CON | H5 | H10 | H15 | H20 | H25 | H30 |
|---|---|---|---|---|---|---|---|
| Diet composition (g/1000 g) | |||||||
| Fish meal | 261 | 234 | 208 | 181 | 155 | 128 | 101 |
| Red blood cells | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| BSFL | 0 | 50 | 100 | 150 | 200 | 250 | 300 |
| Soy protein isolate | 100 | 100 | 100 | 100 | 100 | 100 | 100 |
| Wheat gluten | 150 | 150 | 150 | 150 | 150 | 150 | 150 |
| Wheat meal | 145 | 130 | 117 | 104 | 89 | 76 | 63 |
| Maltodextrin | 130 | 130 | 130 | 130 | 130 | 130 | 130 |
| Fish oil | 65 | 55 | 44 | 34 | 24 | 14 | 3 |
| Lecithin | 10 | 10 | 10 | 10 | 10 | 10 | 10 |
| Premix 1 | 15 | 15 | 15 | 15 | 15 | 15 | 15 |
| Vitamin premix 2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Choline chloride | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| Limestone | 18 | 18 | 16 | 14 | 3 | 11 | 10 |
| Phosphate 1-Ca | 0 | 2 | 4 | 6 | 8 | 10 | 12 |
| TiO2 | 3 | 3 | 3 | 3 | 3 | 3 | 3 |
| Total | 1000 | 1000 | 1000 | 1000 | 1000 | 1000 | 1000 |
| Levels of replacements by BSFL (%) | |||||||
| Fish meal | 0 | 10.3 | 20.3 | 30.7 | 40.6 | 50.9 | 61.3 |
| Fish oil | 0 | 15.4 | 32.3 | 47.7 | 61.3 | 78.5 | 95.4 |
| Analysed chemical feed composition (g/1000 g) | |||||||
| Dry matter | 938.4 | 937.4 | 934.5 | 934.7 | 934.9 | 936.0 | 935.9 |
| Crude protein | 485.5 | 487.7 | 491.4 | 497.9 | 503.5 | 507.4 | 509.7 |
| Crude fat | 99.7 | 102.4 | 101 | 95.5 | 92.1 | 92.3 | 91.0 |
| Crude fibre | 6.7 | 9.9 | 13.3 | 16.7 | 19.9 | 23.2 | 26.5 |
| Ash | 82.1 | 82.3 | 82.2 | 80.9 | 81.2 | 80.1 | 80.1 |
| Nitrogen-free extract | 264.4 | 255.1 | 246.6 | 243.7 | 238.2 | 233.0 | 228.6 |
| Ca | 13.5 | 14.3 | 14.3 | 14.3 | 14.7 | 14.7 | 15.0 |
| P | 8.1 | 8.1 | 8.1 | 8.1 | 8.1 | 8.1 | 8.1 |
| TiO2 | 3.0 | 2.8 | 2.9 | 3.2 | 3.1 | 3.2 | 3.4 |
| Gross energy (MJ/kg) | 20.39 | 20.44 | 20.44 | 20.43 | 20.42 | 20.42 | 20.43 |
| Calcium:phosphorus ratio (g:g) | |||||||
| Ca:P ratio | 1.67:1 | 1.77:1 | 1.76:1 | 1.76:1 | 1.81:1 | 1.81:1 | 1.85:1 |
| ITEM | CON | H5 | H10 | H15 | H20 | H25 | H30 | SEM | p-Value |
|---|---|---|---|---|---|---|---|---|---|
| D (mm) | 3.70 c | 3.72 b,c | 3.79 a,b,c | 3.85 a,b,c | 3.91 a | 3.87 a | 3.93 a | 0.02 | 0.0017 |
| ER (%) | 23.4 c | 24.1 b,c | 26.4 a,b,c | 28.4 a,b | 30.4 a | 29.0 a | 31.1 a | 0.62 | 0.0017 |
| PD (g/dm3) | 603 a | 603 a | 592 b | 586 c | 571 d | 546 e | 532 f | 6.21 | <0.0001 |
| SV (s/100 cm) | 10.9 d | 10.8 d | 11.3 c,d | 11.7 b,c,d | 12.2 b,c | 12.5 a,b | 13.3 a | 0.15 | <0.0010 |
| WS (%) | 97.8 a | 97.3 a | 97.4 a | 97.7 a | 96.9 a,b | 95.8 b | 96.3 b | 1.64 | <0.0001 |
| VI (%) | 24.8 c | 25.2 c | 29.6 c | 36.4 b | 38.0 b | 38.4 b | 48.8 a | 0.24 | <0.0001 |
| Parameter and Day | CON | H5 | H10 | H15 | H20 | H25 | H30 | SEM | p-Value |
|---|---|---|---|---|---|---|---|---|---|
| Growth performance parameters (Experiment 1) | |||||||||
| BW, day 50 | 83.8 c | 94.8 b | 105.7 a | 105.8 a | 106.6 a | 109.2 a | 109.1 a | 1.51 | <0.0001 |
| BWG, days 1–50 | 69.4 c | 80.4 b | 91.3 a | 91.4 a | 94.8 a | 92.2 a | 94.7 a | 1.49 | <0.0001 |
| SGR, days 1–50 | 1.91 c | 2.04 b | 2.17 a | 2.16 a | 2.20 a | 2.17 a | 2.20 a | 0.02 | <0.0001 |
| PWG, days 1–50 | 481 c | 558 b | 635 a | 634 a | 658 a | 641 a | 658 a | 10.0 | <0.0001 |
| Feed utilization parameters (Experiment 1) | |||||||||
| FI, days 1–50 | 61 | 63.3 | 63.6 | 64.3 | 64.6 | 63.3 | 64.25 | 0.50 | 0.2915 |
| FCR, days1–50 | 0.88 a | 0.79 b | 0.69 c | 0.70 c | 0.68 c | 0.68 c | 0.68 c | 0.01 | <0.0001 |
| PER, days 1–50 | 2.34 c | 2.60 b | 2.92 a | 2.86 a | 2.91 a | 2.88 a | 2.90 a | 0.18 | <0.0001 |
| Feed acceptance (Experiment 2) | |||||||||
| FA (% of BW) | 0.21 b | 0.21 b | 0.44 a | 0.49 a | 0.54 a | 0.62a | 0.61 a | 0.04 | 0.0016 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rawski, M.; Mazurkiewicz, J.; Kierończyk, B.; Józefiak, D. Black Soldier Fly Full-Fat Larvae Meal as an Alternative to Fish Meal and Fish Oil in Siberian Sturgeon Nutrition: The Effects on Physical Properties of the Feed, Animal Growth Performance, and Feed Acceptance and Utilization. Animals 2020, 10, 2119. https://doi.org/10.3390/ani10112119
Rawski M, Mazurkiewicz J, Kierończyk B, Józefiak D. Black Soldier Fly Full-Fat Larvae Meal as an Alternative to Fish Meal and Fish Oil in Siberian Sturgeon Nutrition: The Effects on Physical Properties of the Feed, Animal Growth Performance, and Feed Acceptance and Utilization. Animals. 2020; 10(11):2119. https://doi.org/10.3390/ani10112119
Chicago/Turabian StyleRawski, Mateusz, Jan Mazurkiewicz, Bartosz Kierończyk, and Damian Józefiak. 2020. "Black Soldier Fly Full-Fat Larvae Meal as an Alternative to Fish Meal and Fish Oil in Siberian Sturgeon Nutrition: The Effects on Physical Properties of the Feed, Animal Growth Performance, and Feed Acceptance and Utilization" Animals 10, no. 11: 2119. https://doi.org/10.3390/ani10112119
APA StyleRawski, M., Mazurkiewicz, J., Kierończyk, B., & Józefiak, D. (2020). Black Soldier Fly Full-Fat Larvae Meal as an Alternative to Fish Meal and Fish Oil in Siberian Sturgeon Nutrition: The Effects on Physical Properties of the Feed, Animal Growth Performance, and Feed Acceptance and Utilization. Animals, 10(11), 2119. https://doi.org/10.3390/ani10112119