Influence of Age and Immunostimulation on the Level of Toll-Like Receptor Gene (TLR3, 4, and 7) Expression in Foals

 , ,
, ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Feeding
2.2. Experimental Design
- The control group (Group C) (n = 9)—foals without any pharmacological and feed additives that may influence immune system;
- The experimental group (Group E) (n = 7)—foals that were administered an immunostimulating agent.
2.3. Blood Sampling and Blood Analysis
2.4. Statistical Analysis
3. Results
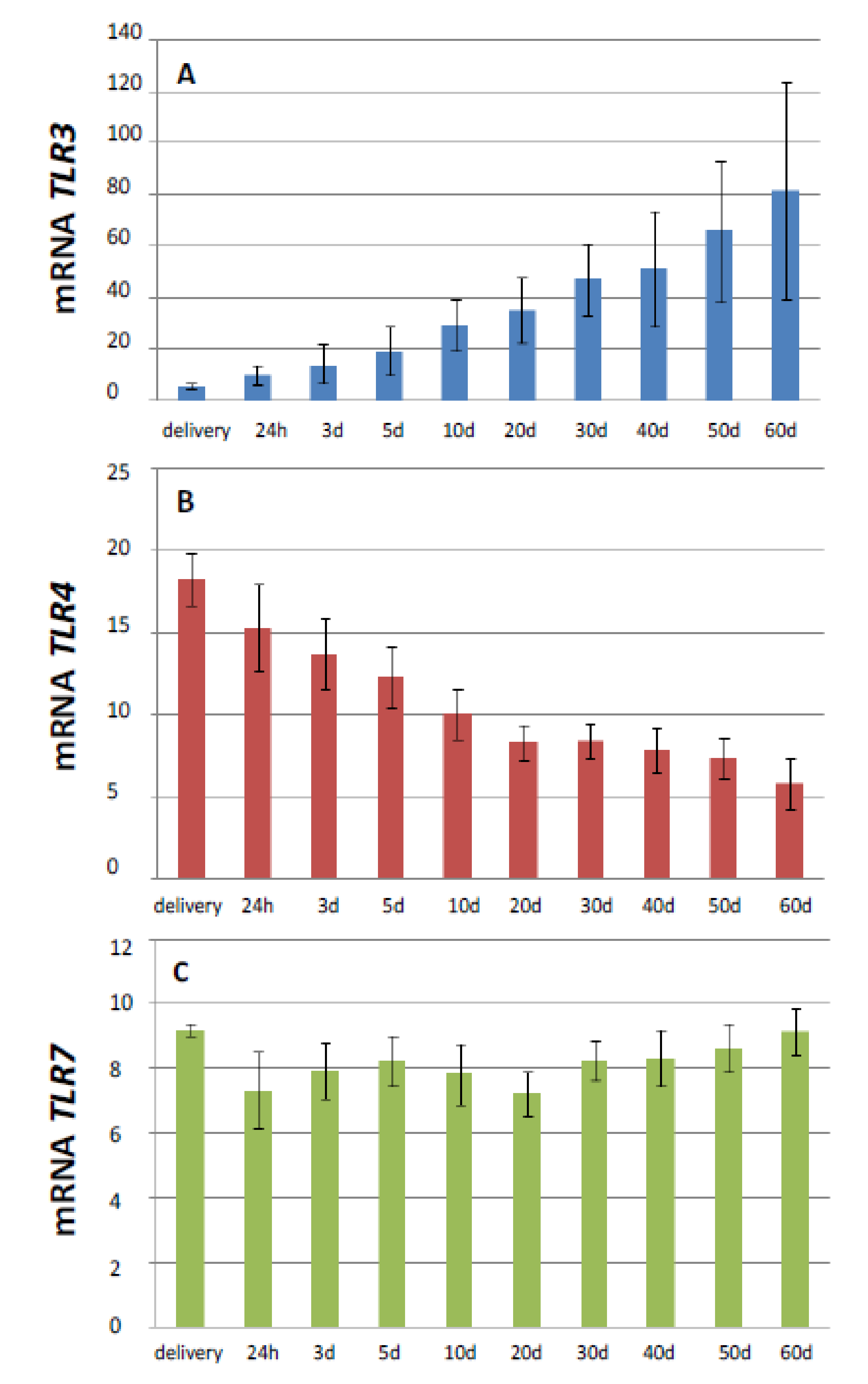
3.1. Influence of Age on the Expression of TLR3, 4, and 7 mRNA
3.2. Influence of Stimulation with Biotropina on the Expression of TLR3, 4, and 7 mRNA
3.3. Influence of Age and Stimulation with Biotropina on the Level of Blood Morphotic Elements
- -
- A significant influence of age on the hematocrit level;
- -
- A highly significant influence of age on the hemoglobin level;
- -
- A significant influence of age on the level of erythrocytes;
- -
- A highly significant influence of age and immunostimulation on the level of leukocytes;
- -
- A highly significant influence of age and immunostimulation on the level of lymphocytes;
- -
- A highly significant influence of immunostimulation on the number of monocytes;
- -
- A significant influence of age and immunostimulation on the number of neutrophils.
- -
- A highly significant influence of immunostimulation, significant influence of age on the number of basophils;
- -
- A highly significant influence of immunostimulation and age on the number of basophils.
4. Discussion
4.1. Changes in TLR4 Gene Expression
4.2. Changes in TLR3 Gene Expression
4.3. Changes in TLR7 Gene Expression
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Tallmadge, R.; McLaughlin, K.; Secor, E.; Ruano, D.; Matychak, M.; Julia, M.; Flaminio, B.F. Expression of essential Bcell genes and immunoglobulin isotypes suggests active development and gene recombination during equine gestation. Dev. Comp. Immunol. 2009, 33, 1027–1038. [Google Scholar] [CrossRef] [PubMed]
- Perkins, G.A.; Wagner, B. The development of equine immunity: Current knowledge on immunology in the young horse. Equine Vet. J. 2015, 47, 267–274. [Google Scholar] [CrossRef]
- Flamino, M.J.; Rush, B.R.; Davis, E.G.; Hennessy, K.; Shuman, W.; Wilkerson, M.J. Characterization of peripheral blood and pulmonary leukocyte function in healthy foal. Vet. Immunol. Immunopathol. 2000, 73, 267–285. [Google Scholar] [CrossRef]
- Miyara, M.; Sakaguchi, S. Natural regulatory T cells: Mechanisms of suppression. Trends Mol. Med. 2007, 13, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Janeway, C.A., Jr.; Medzhitov, R. Innate immune recognition. Annu. Rev. Immunol. 2002, 20, 197–216. [Google Scholar] [CrossRef]
- Lemaitre, B.; Nicolas, E.; Michaut, L.; Reichhart, J.M.; Hoffmann, J.A. The dorsoventral regulatory gene cassette spätzle/Toll/cactus controls the potent antifungal response in Drosophila adults. Cell 1996, 6, 973–983. [Google Scholar] [CrossRef]
- Medzhitov, R.; Preston-Hurlburt, P.; Janeway, C., Jr. A human homologue of the Drosophila Toll protein signals activation of adaptive immunity. Nature 1997, 388, 394–397. [Google Scholar] [CrossRef]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef]
- Harrington, J.R.; Wilkerson, C.P.; Brake, C.N.; Cohen, N.D. Effects of age and R848 stimulation on expression of Toll-like receptor 8mRNA by foal neutrophils. Vet. Immunol. Immunopathol. 2012, 150, 10–18. [Google Scholar] [CrossRef]
- Hayashi, F.; Means, T.K.; Luster, A.D. Toll-like receptors stimulate human neutrophil function. Blood 2003, 102, 2660–2669. [Google Scholar] [CrossRef]
- Kulisa, M.; Makieła, K.; Długosz, B.; Gaj, M. Thoroughbred foals’ mortality causes during first six months of life. PartII. Diseases and injuries. Roczniki Naukowe Polskiego Towarzystwa Zootechnicznego 2009, 5, 79–84. [Google Scholar]
- Haas, S.D.; Bristol, F.; Card, C.E. Risk factors associated with the incidence of foal mortality in a managed mare herd. Can. Vet. J. 1996, 37, 91–95. [Google Scholar] [PubMed]
- Wilson, W.D.; Mihalyi, J.E.; Hussey, S.; Lunn, D.P. Passive transfer of maternal immunoglobulin isotype antibodies against tetanus and influenza and their effect on the response of foals to vaccination. Equine Vet. J. 2001, 33, 644–650. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, D.M.; Eicker, S.W.; Yeagar, A.E.; Sweeney, C.R.; Viel, L.; Tesarowski, D.; Lavoie, J.P.; Hoffman, A.; Paradis, M.R.; Reed, S.M.; et al. Associations between physical examination, laboratory, and radiographic findings and outcome and subsequent racing performance of foals with Rhodococcus equi infection: 115 cases (1984–1992). J. Am. Vet. Med. Assoc. 1998, 213, 510–515. [Google Scholar] [PubMed]
- Breathnach, C.C.; Sturgill-Wright, T.; Stiltner, J.L.; Adams, A.A.; Lunn, D.P.; Horohov, D.W. Foals are interferon gamma-deficient at birth. Vet. Immunol. Immunopathol. 2006, 112, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Boyd, N.K.; Cohen, N.D.; Lim, W.S.; Martens, R.J.; Chaffin, M.K.; Ball, J.M. Temporal changes in cytokine expression of foals during the first month of life. Vet. Immunol. Immunopathol. 2003, 92, 75–85. [Google Scholar] [CrossRef]
- Prescott, J.F.; Nicholson, V.M.; Patterson, M.C.; Zandona Meleiro, M.C.; de Caterino, A.A.; Yager, J.A.; Holmes, M.A. Use of Rhodococcus equi virulence-associated protein for immunization of foals against R. equi pneumonia. Am. J. Vet. Res. 1997, 58, 356–359. [Google Scholar]
- Fleer, A.; Krediet, T.G. Innate immunity: Toll-like receptors and some more. Neonatology 2007, 92, 145–157. [Google Scholar] [CrossRef]
- Levy, O. Innate immunity of the newborn: Basic mechanisms and clinical correlates. Nat. Rev. Immunol. 2007, 7, 379–390. [Google Scholar] [CrossRef]
- Jaworski, Z. Tablice Genealogiczne Koników Polskich Genealogical Tables of the Polish Primitive Horse; Stacja Badawcza Rolnictwa Ekologicznego i Hodowli Zachowawczej Zwierzat PAN: Popielno, Poland, 1997. (In Polish) [Google Scholar]
- Hoehler, D. The Institute for Animal Nutrition and Metabolic Physiology; Kiel University: Kiel, Germany, 1997. [Google Scholar]
- Flaminio, M.; Rush, B.; Shuman, W. Peripheral Blood Lymphocyte Subpopulations and Immunoglobulin Concentrations in Healthy Foals and Foals with Rhodococcus Equi Pneumonia. J. Vet. Intern. Med. 1999, 13, 206–212. [Google Scholar] [CrossRef]
- Tallmadge, R.; Wang, M.; Sun, Q.; Felippe, M.J.B. Transcriptome analysis of immune genes in peripheral blood mononuclear cells of young foals and adult horses. PLoS ONE 2018, 13, e0202646. [Google Scholar] [CrossRef] [PubMed]
- Vendrig, J.C.; Coffeng, L.E.; Fink-Gremmels, J. Effects of Separate and Concomitant TLR-2 and TLR-4 Activation in Peripheral Blood Mononuclear Cells of Newborn and Adult Horses. PLoS ONE 2013, 8, e66897. [Google Scholar] [CrossRef] [PubMed]
- Tessier, L.; Bienzle, D.; Williams, L.B.; Koch, T.G. Phenotypic and Immunomodulatory Properties of Equine Cord Blood-Derived Mesenchymal Stromal Cells. PLoS ONE 2015, 10, e0122954. [Google Scholar] [CrossRef] [PubMed]
- Hansen, S.; Baptiste, K.; Fjeldborg, J.; Betancourt, A.; Horohov, D. A comparison of pro-inflammatory cytokine mRNA expression in equine bronchoalveolar lavage (BAL) and peripheral blood. Vet. Immunol. Immunopathol. 2014, 158, 238–243. [Google Scholar] [CrossRef]
- Osorio, J.; Trevisi, E.; Ballou, M.; Bertoni, G.; Drackley, J.; Loor, J. Effect of the level of maternal energy intake prepartum on immune metabolic markers, polymorphonuclear leukocyte function, and neutrophil gene network expression in neonatal Holstein heifer calves. J. Dairy Sci. 2013, 96, 3573–3587. [Google Scholar] [CrossRef]
- Strong, R.; Silva, E.; Cheng, H.; Eicher, S.D. Acute brief heat stress in late gestation alters neonatal calf innate immune functions. J. Dairy Sci. 2015, 98, 7771–7783. [Google Scholar] [CrossRef]
- Yerkovich, S.T.; Wikstrom, M.E.; Suriyaarachchi, D.; Prescott, S.L.; Upham, J.W.; Holt, P.G. Postnatal Development of Monocyte Cytokine Responses to Bacterial Lipopolysaccharide. Pediatr. Res. 2007, 62, 547–552. [Google Scholar] [CrossRef]
- Levy, E.; Xanthou, G.; Petrakou, E.; Zacharioudaki, V.; Tsatsanis, C.; Fotopoulos, S.; Xanthou, M. Distinct Roles of TLR4 and CD14 in LPS-Induced Inflammatory Responses of Neonates. Pediatr. Res. 2009, 66, 179–184. [Google Scholar] [CrossRef]
- Boehmer, E.D.; Goral, J.; Faunce, D.E.; Kovacs, E.J. Age-dependent decrease in Toll-like receptor 4-mediated proinflammatory cytokine production and mitogen-activated protein kinase expression. J. Leukoc. Biol. 2003, 75, 342–349. [Google Scholar] [CrossRef]
- Chelvarajan, R.L.; Collins, S.M.; Van Willigen, J.M.; Bondada, S. The unresponsiveness of aged mice to polysaccharide antigens is a result of a defect in macrophage function. J. Leukoc. Biol. 2005, 77, 503–512. [Google Scholar] [CrossRef]
- Förster-Waldl, E.; Sadeghi, K.; Tamandl, D.; Gerhold, B.; Hallwirth, U.; Meistersinger, K.; Hayde, M.; Prusa, A.R.; Herkner, K.; Boltz-Nitulescu, G.; et al. Monocyte toll-like receptor 4 expression and LPS induced cytokine production increase during gestational aging. Pediatr. Res. 2005, 58, 121–124. [Google Scholar] [CrossRef] [PubMed]
- Levy, O.; Zarember, K.A.; Roy, R.M.; Cywes, C.; Godowski, P.J.; Wessels, M.R. Selective impairment of TLR-mediated innate immunity in human newborns: Neonatal blood plasma reduces monocyte TNF-alpha induction by bacterial lipopeptides, lipopolysaccharide, and imiquimod, but preserves the response to R-848. J. Immunol. 2004, 173, 4627–4634. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.R.; Qing, G.; Byers, D.M.; Stadnyk, A.W.; Al-Hertani, W.; Bortolussi, R. Role of MyD88 in Diminished Tumor Necrosis Factor Alpha Production by Newborn Mononuclear Cells in Response to Lipopolysaccharide. Infect. Immun. 2004, 72, 1223–1229. [Google Scholar] [CrossRef]
- Porrás, A.; Kozar, S.; Russanova, V.; Salpea, P.; Hirai, T.; Sammons, N.; Mittal, P.; Kim, J.Y.; Ozato, K.; Romero, R.; et al. Developmental and epigenetic regulation of the human TLR3 gene. Mol. Immunol. 2008, 46, 27–36. [Google Scholar] [CrossRef]
- Zhang, J.; Wei, H.; Wu, D.; Tian, Z. Toll-like receptor 3 agonist induces impairment of uterine vascular remodeling and fetal losses in CBA × DBA/2 mice. J. Reprod. Immunol. 2007, 74, 61–67. [Google Scholar] [CrossRef]
- Gibbons, D.L.; Haque, S.F.; Silberzahn, T.; Hamilton, K.; Langford, C.; Ellis, P.; Carr, R.; Hayday, A.C. Neonates harbour highly active gamma delta T cells with selective impairments in preterm infants. Eur. J. Immunol. 2009, 39, 1794–1806. [Google Scholar] [CrossRef] [PubMed]
- Pott, J.; Stockinger, S.; Torow, N.; Smoczek, A.; Lindner, C.; McInerney, G.; Bäckhed, F.; Baumann, U.; Pabst, O.; Bleich, A.; et al. Age-Dependent TLR3 Expression of the Intestinal Epithelium Contributes to Rotavirus Susceptibility. PLoS Pathog. 2012, 8, e1002670. [Google Scholar] [CrossRef]
- Slavica, L.; Nordström, I.; Karlsson, M.N.; Valadi, H.; Kacerovsky, M.; Jacobsson, B.; Eriksson, K. TLR3 impairment in human newborns. J. Leukoc. Biol. 2013, 94, 1003–1011. [Google Scholar] [CrossRef]
- Hussey, G.S.; Ashton, L.V.; Quintana, A.M.; Lunn, P.D.; Goehring, L.S.; Annis, K.; Landolt, G. Innate immune responses of airway epithelial cells to infection with Equine herpesvirus-1. Vet. Microbiol. 2014, 170, 28–38. [Google Scholar] [CrossRef]
- Asquith, M.; Haberthur, K.; Brown, M.; Engelmann, F.; Murphy, A.; Al-Mahdi, Z.; Messaoudi, I. Age-dependent changes in innate immune phenotype and function in rhesus macaques (Macaca mulatta). Pathobiol. Aging Age Relat. Dis. 2012, 2, 3331. [Google Scholar] [CrossRef]
- Kwon, S.; Vandenplas, M.L.; Figueiredo, M.D.; Salter, C.E.; Andrietti, A.L.; Robertson, T.P.; Moore, J.N.; Hurley, D.J. Differential induction of Toll-like receptor gene expression in equine monocytes activated by Toll-like receptor ligands or TNF-α. Vet. Immunol. Immunopathol. 2010, 138, 213–217. [Google Scholar] [CrossRef]
- Belnoue, E.; Fontannaz, P.; Rochat, A.-F.; Tougne, C.; Bergthaler, A.; Lambert, P.-H.; Pinschewer, D.D.; Siegrist, C.-A. Functional Limitations of Plasmacytoid Dendritic Cells Limit Type I Interferon, T Cell Responses and Virus Control in Early Life. PLoS ONE 2013, 8, e85302. [Google Scholar] [CrossRef]


{kind=link}
| Gen. | Full Name of the Gene | Access Number GenBank | TaqMan Gene Expression Assay ID | Dye |
|---|---|---|---|---|
| TLR3 | Toll-like receptor 3 | NC_009170.2 | Ec03467747_m1 | FAM |
| TLR4 | Toll-like receptor 4 | NC_009168.2 | Ec03468993_m1 | FAM |
| TLR7 | Toll-like receptor 7 | NC_009175.2 | Ec03467530_m1 | VIC |
| SDHA | Succinate dehydrogenase complex subunit A | XM_001490889 | Ec03470479_m1 | VIC |
| HPRT | Hypoxanthinephosphoribosyl transferase | AY372182.1 | Ec03470217_m1 | VIC |
| Age | TLR3 | TLR4 | TLR7 |
|---|---|---|---|
| Delivery 1 | 6.2 A ± 0.9 2 | 18.3 A ± 2.6 | 9.2 ± 1.2 |
| 24 h | 9.8 B ± 1.7 | 15.3 B ± 2.2 | 7.3 ± 0.9 |
| 3 days | 14.1 A,C ± 2.1 | 13.7 C ± 1.8 | 7.9 ± 0.8 |
| 5 days | 19.1 A,B,D ± 2.2 | 12.3 A,D ± 1.5 | 8.2 ± 0.9 |
| 10 days | 29.3 A,B,C,E ± 2.8 | 10.1 A ± 1.1 | 7.8 ± 0.7 |
| 20 days | 35.1 A,B,C,F ± 3.1 | 8.3 A,B ± 1.1 | 7.2 ± 0.6 |
| 30 days | 48.4 A,B,C,D,G ± 4.9 | 8.4 A,B ± 14 | 8.3 ± 0.9 |
| 40 days | 53.6 ABCDEF ± 5.8 | 7.8 A,B,C ± 1.2 | 8.3 ± 0.7 |
| 50 days | 80.4 ABCDEFG ± 8.7 | 7.3 A,B,C ± 1.5 | 8.6 ± 0.7 |
| 60 days | 87.9 A,B,C,D,E,F,G ± 8.0 | 5.8 A,B,C,D ± 0.9 | 9.2 ± 07 |
| Age | TLR3 | TLR4 | TLR7 | |||
|---|---|---|---|---|---|---|
| Group C | Group E | Group C | Group E | Group C | Group E | |
| Delivery 1 | 8.3 ** ± 1.8 2 | 4.1 ** ± 0.55 | 19.9 * ± 4.0 | 13.7 * ± 0.5 | 12.9 * ± 2.9 | 7.6 * ± 0.9 |
| 24 h | 14.7 ** ± 3.8 | 6.5 ** ± 1.24 | 17.5 * ± 3.3 | 11.5 * ± 0.6 | 10.2 ± 2.1 | 7.0 ± 0.8 |
| 3 days | 20.0 ** ± 4.9 | 8.7 ** ± 2.24 | 16.7 ** ± 2.4 | 9.9 ** ± 1.1 | 9.6 ± 1.7 | 8.3 ± 1.1 |
| 5 days | 28.0 ± 2.6 | 15.1 * ± 1.70 | 14.5 * ± 1.9 | 9.7 * ± 1.1 | 11.4 * ± 2.2 | 7.4 * ± 0.9 |
| 10 days | 32.7 ± 3.3 | 30.1 ± 5.43 | 10.9 ± 0.7 | 7.1 ± 1.3 | 9.7 ± 1.4 | 7.4 ± 0.6 |
| 20 days | 36.7 ± 4.6 | 36.4 ± 4.93 | 8.0 ± 1.2 | 7.2 ± 1.5 | 8.7 ± 1.5 | 7.0 ± 0.7 |
| 30 days | 62.2 * ± 9.6 | 39.6 * ± 7.43 | 8.2 ± 2.3 | 7.1 ± 1.9 | 10.0 ± 2.0 | 7.1 ± 0.9 |
| 40 days | 78.2 ** ± 6.8 | 23.1 ** ± 8.82 | 6.5 * ± 1.3 | 10.0 * ± 2.4 | 9.7 ± 1.6 | 7.9 ± 0.8 |
| 50 days | 107.0 ** ± 15.3 | 26.0 ** ± 9.71 | 5.5 ± 1.9 | 6.8 ± 1.9 | 11.2 * ± 1.2 | 6.9 * ± 0.9 |
| 60 days | 116.2 ** ± 13.9 | 44.6 ** ± 10.20 | 4.2 ± 1.4 | 6.5 ± 1.7 | 11.5 ± 1.2 | 7.6 ± 0.7 |
| Gene | Age | TLR4 | TLR7 |
|---|---|---|---|
| TLR3 | <1 h | −0.160 (0.6273) | 0.62857 (0.1631) |
| 24 h | −0.191 (0.4199) | 0.462 (0.1400) | |
| 3 days | −0.100 (0.6726) | 0.492 (0.1276) | |
| 5 days | −0.095 (0.6912) | 0.328 (0.1582) | |
| 10 days | −0.350 (0.0299) | 0.567 (0.1917) | |
| 20 days | −0.582 (0.0071) | 0.423 (0.1634) | |
| 30 days | −0.04361 (0.0085) | 0.368 (0.1098) | |
| 40 days | −0.56992 (0.0087) | 0.472 (0.0355) | |
| 50 days | −0.46466 (0.0039) | 0.76541 (0.0251) | |
| 60 days | −0.35338 (0.0026) | 0.82105 (0.0341) | |
| TLR4 | <1 h | −0.340 (0.1376) | |
| 24 h | −0.385 (0.0936) | ||
| 3 days | −0.472 (0.1355) | ||
| 5 days | −0.341 (0.1408) | ||
| 10 days | −0.191 (0.4199) | ||
| 20 days | −0.319 (0.1707) | ||
| 30 days | −0.271 (0.2468) | ||
| 40 days | −0.53083 (0.1600) | ||
| 50 days | −0.67519 (0.0111) | ||
| 60 days | −0.360 (0.0116) | ||
| Age | <1 h 1 | 24 h | 3 Days | 5 Days | 10 Days | 20 Days | 30 Days | 40 Days | 50 Days | 60 Days | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Parameters | |||||||||||
| Hematocrit (PCV) % | C | 50.00 2 ± 1.2 | 43.33 ± 1.2 | 39.83 ± 1.5 | 41.11 ± 1.1 | 41.56 ± 1.9 | 34.44 ± 1.6 | 37.33 ± 1.1 | 36.33 ± 1.3 | 37.72 ± 1.2 | 37.56 ± 1.7 |
| E | 50.50 ± 1.2 | 44.50 ± 1.8 | 43.67 ± 1.7 | 39.58 ± 1.5 | 41.17 ± 0.9 | 38.20 ± 1.1 | 39.75 ± 2.9 | 36.50 ± 1.7 | 37.25 ± 1.3 | 37.75 ± 0.9 | |
| Hemoglobin (g/dL) | C | 15.46 ± 0.7 | 14.55 ± 0.7 | 13.26 ± 0.5 | 13.75 ± 0.7 | 13.63 ± 0.7 | 14.57 * ± 1.2 | 15.11 * ± 0.9 | 14.85 * ± 1.3 | 13.30 ± 0.6 | 13.98 * ± 1.1 |
| E | 13.94 ± 0.9 | 13.04 ± 1.7 | 13.50 ± 0.8 | 12.71 ± 0.6 | 12.39 ± 0.2 | 13.03 ± 0.9 | 11.80 * ± 0.3 | 11.70 * ± 0.4 | 12.39 ± 0.5 | 10.08 * ± 0.3 | |
| RBC count (106/µL) | C | 10.93 ± 0.9 | 10.62 ± 0.7 | 9.99 ± 0.6 | 10.55 ± 0.6 | 9.53 ± 0.4 | 10.38 ± 1.3 | 11.24 ± 1.5 | 9.98 ± 0.8 | 10.43 ± 0.8 | 9.86 ± 0.8 |
| E | 11.50 ± 0.5 | 10.01 ± 0.3 | 9.17 ± 0.4 | 10.24 ± 0.9 | 9.33 ± 0.8 | 9.04 ± 0.7 | 9.59 ± 1.0 | 9.69 ± 0.7 | 10.29 ± 0.3 | 10.22 ± 0.3 | |
| WBC count (103/µL) | C | 7.35 ± 0.8 | 7.71 ± 0.7 | 10.35 ± 1.2 | 9.86 ± 1.0 | 11.03 ± 0.8 | 11.30 ± 0.8 | 12.60 ± 1.0 | 14.31 ** ± 0.8 | 14.62 * ± 0.7 | 13.07 ** ± 0.7 |
| E | 6.25 ± 0.6 | 8.17 ± 0.5 | 9.15 ± 0.8 | 10.64 ± 1.4 | 12.40 ± 1.1 | 12.48 ± 0.9 | 10.91 * ± 1.6 | 21.75 ** ± 0.5 | 18.98 * ± 0.4 | 14.89 ** ± 0.2 | |
| Eosinophils (/µL) | C | 105 ** ± 2.2 | 116 ** ± 2.4 | 72 ** ± 1.5 | 296 ** ± 6.2 | 110 ** ± 2.3 | 226 ** ± 4.7 | 315 ** ± 6.6 | 286 ** ± 6.0 | 292 ** ± 6.1 | 327 ** ± 6.9 |
| E | 0 ** ± 0.0 | 16 ** ± 0.3 | 22 ** ± 0.4 | 31 ** ± 0.6 | 37 ** ± 0.7 | 42 ** ± 0.8 | 55 ** ± 1.0 | 206 ** ± 3.7 | 105 ** ± 1.9 | 133 ** ± 2.4 | |
| Basophils (/µL) | C | 44 ± 0.9 | 39 ± 0.8 | 31 ± 0.6 | 99 ± 2.1 | 55 ± 1.2 | 113 * ± 2.4 | 94 * ± 1.9 | 72 ** ± 1.5 | 146 ** ± 3.1 | 196 ** ± 4.1 |
| E | 18 ± 0.3 | 29 ± 0.5 | 38 ± 0.7 | 39 ± 0.7 | 52 ± 0.9 | 64 * ± 1.1 | 75 * ± 1.3 | 308 ** ± 5.5 | 205 ** ± 3.7 | 76 ** ± 1.4 | |
| Neutrophils (/µL) | C | 4471 * ± 59.5 | 4488 * ± 59.3 | 7041 ± 93.4 | 6903 ± 91.6 | 5898 * ± 78.3 | 6104 ± 81.0 | 7150 * ± 94.9 | 8583 ± 113.9 | 8186 ± 108.6 | 6403 * ± 84.9 |
| E | 5650 ± 50.8 | 6942 ± 62.5 | 7374 ± 66.4 | 7645 ± 68.8 | 8804 * ± 79.2 | 7773 ± 69.9 | 5705 * ± 51.3 | 9499 * ± 85.5 | 9109 ± 81.9 | 7523 ± 67.7 | |
| Lymphocytes (/µL) | C | 2554 ** ± 35.7 | 2852 * ± 39.9 | 3003 * ± 42.0 | 2465 ± 34.5 | 4686 ** ± 65.6 | 4635 * ± 64.9 | 4945 ** ± 69.2 | 5150 ** ± 72.1 | 5701 ** ± 79.8 | 5880 ** ± 82.3 |
| E | 470 ** ± 4.2 | 1090 * ± 9.8 | 1609 ** ± 14.5 | 2802 ± 25.2 | 3110 ** ± 28.0 | 4280 * ± 38.5 | 4850 ** ± 43.6 | 10953 ** ± 98.6 | 9355 ** ± 84.2 | 7054 ** ± 63.5 | |
| Myelocytes (/µL) | C | 162 ± 2.3 | 231 ** ± 3.2 | 207 * ± 2.9 | 99 ± 1.4 | 276 * ± 3.7 | 226 * ± 3.2 | 94 ** ± 1.3 | 215 ** ± 3.0 | 292 * ± 4.1 | 261 * ± 3.7 |
| E | 112 ± 1.0 | 93 **± 0.8 | 107 *± 0.9 | 123 ± 1.1 | 397 * ± 3.6 | 321 *± 2.9 | 225 *± 2.0 | 784 **± 7.1 | 206 * ± 1.8 | 104 * ± 0.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Migdał, A.; Migdał, Ł.; Oczkowicz, M.; Okólski, A.; Chełmońska-Soyta, A. Influence of Age and Immunostimulation on the Level of Toll-Like Receptor Gene (TLR3, 4, and 7) Expression in Foals. Animals 2020, 10, 1966. https://doi.org/10.3390/ani10111966
Migdał A, Migdał Ł, Oczkowicz M, Okólski A, Chełmońska-Soyta A. Influence of Age and Immunostimulation on the Level of Toll-Like Receptor Gene (TLR3, 4, and 7) Expression in Foals. Animals. 2020; 10(11):1966. https://doi.org/10.3390/ani10111966
Chicago/Turabian StyleMigdał, Anna, Łukasz Migdał, Maria Oczkowicz, Adam Okólski, and Anna Chełmońska-Soyta. 2020. "Influence of Age and Immunostimulation on the Level of Toll-Like Receptor Gene (TLR3, 4, and 7) Expression in Foals" Animals 10, no. 11: 1966. https://doi.org/10.3390/ani10111966
APA StyleMigdał, A., Migdał, Ł., Oczkowicz, M., Okólski, A., & Chełmońska-Soyta, A. (2020). Influence of Age and Immunostimulation on the Level of Toll-Like Receptor Gene (TLR3, 4, and 7) Expression in Foals. Animals, 10(11), 1966. https://doi.org/10.3390/ani10111966