Yellow Mealworm Inclusion in Diets for Heavy-Size Broiler Chickens: Implications for Intestinal Microbiota and Mucin Dynamics

 , ,
, ,  ,
,  ,
,  , , and
, , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Birds and Experimental Design
2.2. Intestinal Sampling and Processing
2.3. DNA Extraction and Sequencing and Bioinformatics Analysis
2.4. Histochemical Staining
2.5. Mucin Staining Intensity
2.6. Statistical Analysis
3. Results
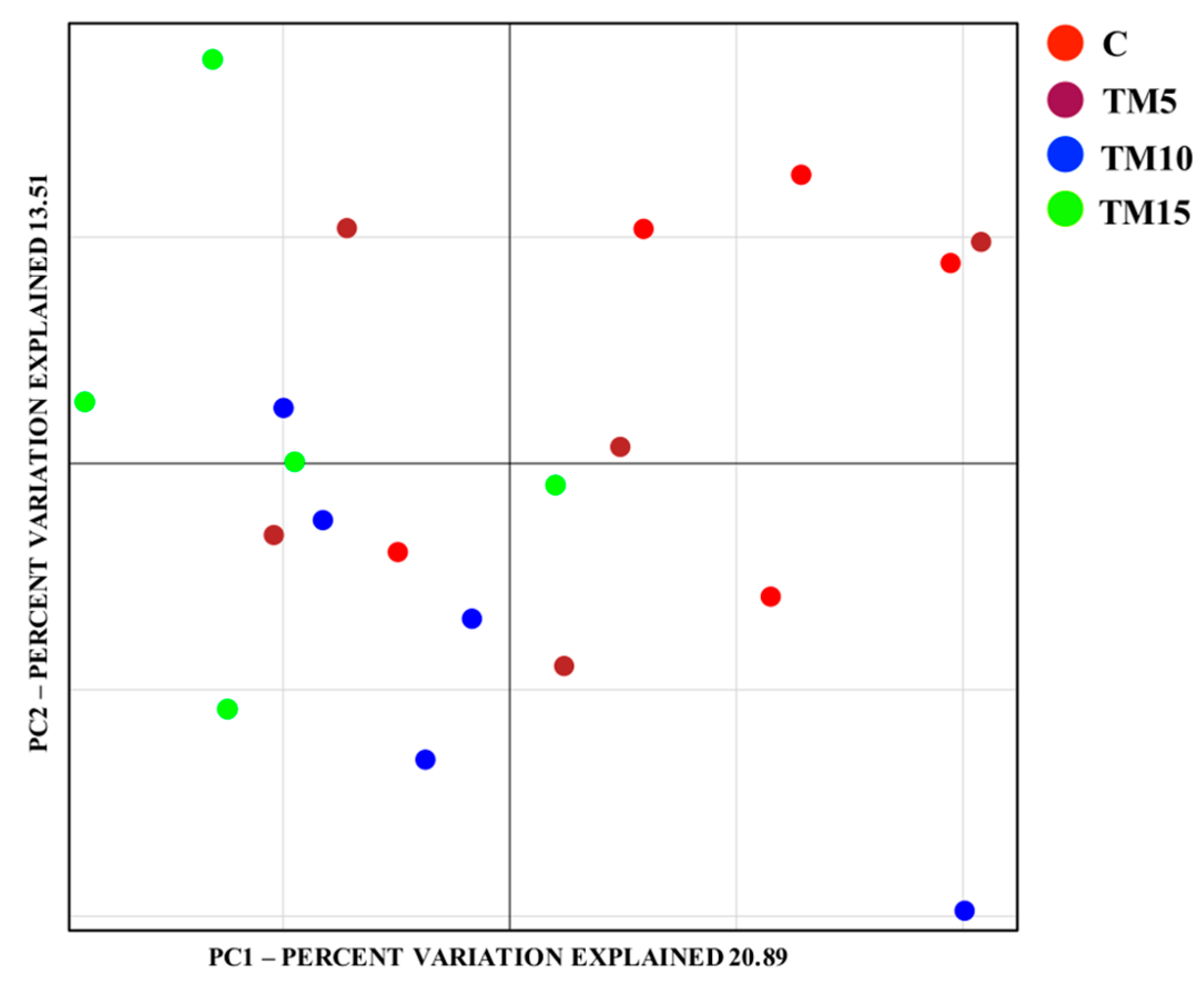
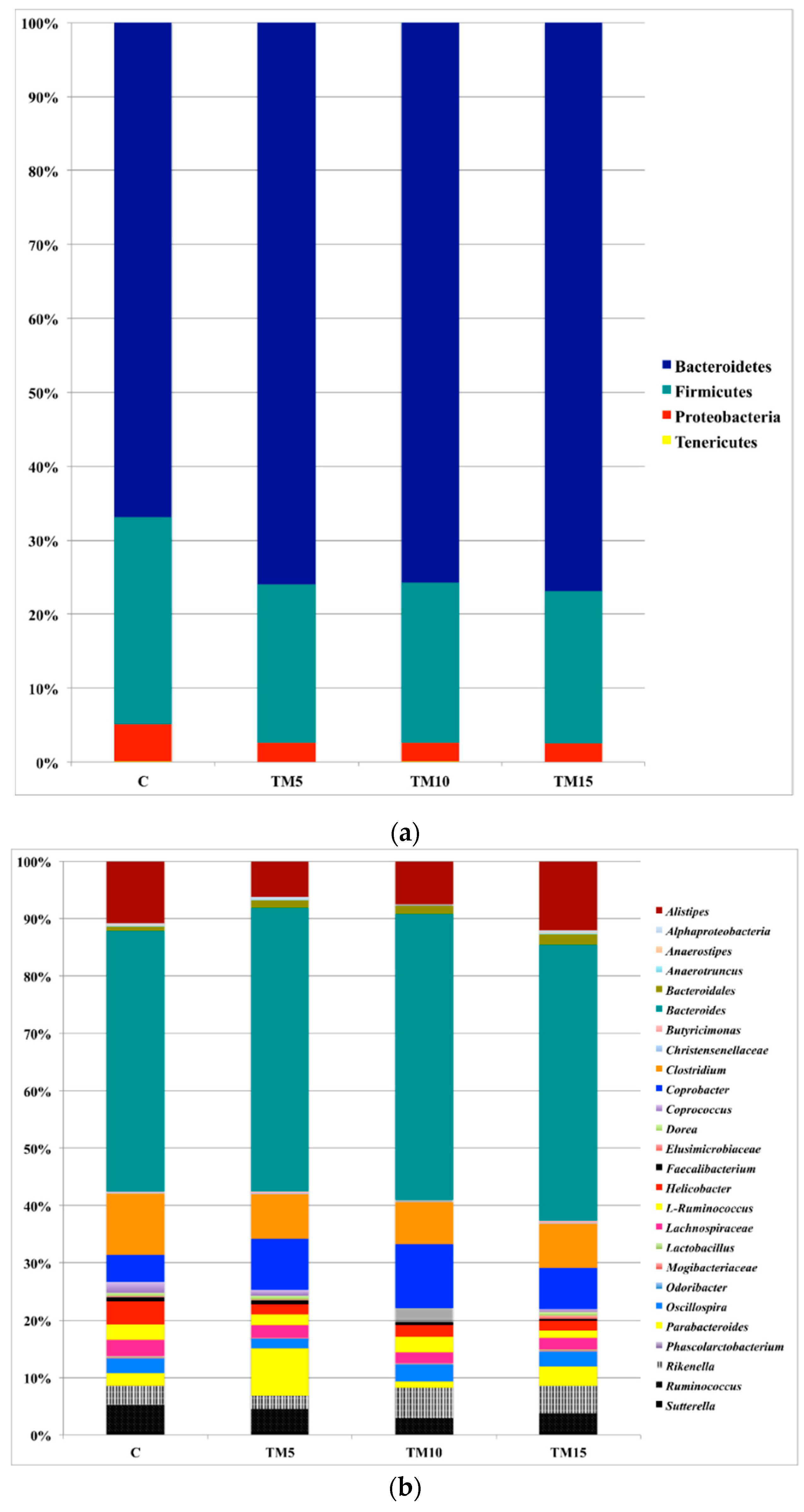
3.1. Cecal Microbiota
3.2. Intestinal Mucin Composition
4. Discussion
4.1. Cecal Microbiota
4.2. Intestinal Mucin Composition
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bianchi, M.; Petracci, M.; Sirri, F.; Folegatti, E.; Franchini, A.; Meluzzi, A. The influence of the season and market class of broiler chickens on breast meat quality traits. Poult. Sci. 2007, 86, 959–963. [Google Scholar] [CrossRef] [PubMed]
- Lorenzi, M.; Mudalal, S.; Cavani, C.; Petracci, M. Incidence of white striping under commercial conditions in medium and heavy broiler chickens in Italy. J. Appl. Poult. Res. 2014, 23, 754–758. [Google Scholar] [CrossRef]
- Sirri, F.; Maiorano, G.; Tavaniello, S.; Ken, J.; Petracci, M.; Meluzzi, A. Effect of different levels of dietary zinc, manganese, and copper from organic or inorganic sources on performance, bacterial chondronecrosis, intramuscular collagen characteristics, and occurrence of meat quality defects of broiler chickens. Poult. Sci. 2016, 95, 1813–1824. [Google Scholar] [CrossRef] [PubMed]
- Soglia, F.; Mudalal, S.; Babini, E.; Di Nunzio, M.; Mazzoni, M.; Sirri, F.; Cavani, C.; Petracci, M. Histology, composition, and quality traits of chicken Pectoralis major muscle affected by wooden breast abnormality. Poult. Sci. 2016, 95, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Celi, P.; Cowieson, A.J.; Fru-Nji, F.; Steinert, R.E.; Kluenter, A.M.; Verlha, V. Gastrointestinal functionality in animal nutrition and health: New opportunities for sustainable animal production. Anim. Feed Sci. Technol. 2017, 234, 88–100. [Google Scholar] [CrossRef]
- Diaz Carrasco, J.M.; Casanova, N.A.; Fernández Miyakawa, M.E. Microbiota, Gut Health and Chicken Productivity: What Is the Connection? Microorganisms 2019, 7, 374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vancamelbeke, M.; Vermeire, S. The intestinal barrier: A fundamental role in health and disease. Expert Rev. Gastroenterol. Hepatol. 2017, 11, 821–834. [Google Scholar] [CrossRef] [PubMed]
- Tsirtsikos, P.; Fegeros, K.; Kominakis, A.; Balaskas, C.; Mountzouris, K.C. Modulation of intestinal mucin composition and mucosal morphology by dietary phytogenic inclusion level in broilers. Animal 2012, 6, 1049–1057. [Google Scholar] [CrossRef] [Green Version]
- Makkar, H.P.S.; Tran, G.; Heuzé, V.; Ankers, P. State of the art on use of insects as animal feed. Anim. Feed Sci. Technol. 2014, 197, 1–33. [Google Scholar] [CrossRef]
- Oonincx, D.G.; van Itterbeeck, J.; Heetkamp, M.-J.; van den Brand, H.; van Loon, J.J.; van Huis, A. An exploration on greenhouse gas and ammonia production by insect species suitable for animal or human consumption. PLoS ONE 2010, 5, e14445. [Google Scholar] [CrossRef] [Green Version]
- Oonincx, D.G.; de Boer, I.J. Environmental impact of the production of mealworms as a protein source for humans—A life cycle assessment. PLoS ONE 2012, 7, e51145. [Google Scholar] [CrossRef] [Green Version]
- Meneguz, M.; Schiavone, A.; Gai, F.; Dama, A.; Lussiana, C.; Renna, M.; Gasco, L. Effect of rearing substrate on growth performance, waste reduction efficiency and chemical composition of black soldier fly (Hermetia illucens) larvae. J. Sci. Food Agric. 2018, 98, 5776–5784. [Google Scholar] [CrossRef]
- Gasco, L.; Finke, M.; van Huis, A. Can diets containing insects promote animal health? J. Insects Food Feed. 2018, 4, 1–4. [Google Scholar] [CrossRef]
- Gasco, L.; Biasato, I.; Dabbou, S.; Schiavone, A.; Gai, F. Animals Fed Insect-Based Diets: State-of-the-Art on Digestibility, Performance and Product Quality. Animals 2019, 9, 170. [Google Scholar] [CrossRef] [Green Version]
- Biasato, I.; Ferrocino, I.; Grego, E.; Dabbou, S.; Gai, F.; Gasco, L.; Cocolin, L.; Capucchio, M.T.; Schiavone, A. Gut Microbiota and Mucin Composition in Female Broiler Chickens Fed Diets including Yellow Mealworm (Tenebrio molitor L.). Animals 2019, 9, 213. [Google Scholar] [CrossRef] [Green Version]
- Biasato, I.; Ferrocino, I.; Dabbou, S.; Evangelista, R.; Gai, F.; Gasco, L.; Cocolin, L.; Capucchio, M.T.; Schiavone, A. Black soldier fly and gut health in broiler chickens: Insights into the relationship between cecal microbiota and intestinal mucin composition. J. Anim. Sci. Biotechnol. 2020, 11, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kers, J.G.; Velkers, F.C.; Fischer, E.A.J.; Hermes, G.D.A.; Stegeman, J.A.; Smidt, H. Host and Environmental Factors Affecting the Intestinal Microbiota in Chickens. Front. Microbiol. 2018, 9, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biasato, I.; Gasco, L.; De Marco, M.; Renna, M.; Rotolo, L.; Dabbou, S.; Capucchio, M.T.; Biasibetti, E.; Tarantola, M.; Sterpone, L.; et al. Yellow mealworm larvae (Tenebrio molitor) inclusion in diets for male broiler chickens: Effects on growth performance, gut morphology, and histological findings. Poult. Sci. 2018, 97, 540–548. [Google Scholar] [CrossRef]
- Biasato, I.; Ferrocino, I.; Biasibetti, E.; Grego, E.; Dabbou, S.; Sereno, A.; Gai, F.; Gasco, L.; Schiavone, A.; Cocolin, L.; et al. Modulation of intestinal microbiota, morphology and mucin composition by dietary insect meal inclusion in free-range chickens. BMC Vet. Res. 2018, 14, 383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Dixon, P. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- Wise, M.G.; Siragusa, G.R. Quantitative analysis of the intestinal bacterial community in one- to three-week-old commercially reared broiler chickens fed conventional or antibiotic-free vegetable-based diets. J. Appl. Microbiol. 2007, 102, 1138–1149. [Google Scholar] [CrossRef]
- Tillman, G.E.; Haas, G.J.; Wise, M.G.; Oakley, B.; Smith, M.A.; Siragusa, G.R. Chicken intestine microbiota following the administration of lupulone, a hop-based antimicrobial. FEMS Microbiol. Ecol. 2011, 77, 395–403. [Google Scholar] [CrossRef] [Green Version]
- Wei, S.; Morrison, M.; Yu, Z. Bacterial census of poultry intestinal microbiome. Poult. Sci. 2013, 92, 671–683. [Google Scholar] [CrossRef]
- Oakley, B.B.; Lillehoj, H.S.; Kogut, M.H.; Kim, W.K.; Maurer, J.J.; Pedroso, A.; Lee, M.D.; Collett, S.R.; Johnson, T.J.; Cox, N.A. The chicken gastrointestinal microbiome. FEMS Microbiol. Lett. 2014, 360, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Si, W.; Forster, R.J.; Huang, R.; Yu, H.; Yin, Y.; Yang, C.; Han, Y. 16S rRNA gene-based analysis of mucosa-associated bacterial community and phylogeny in the chicken gastrointestinal tracts: From crops to ceca. FEMS Microbiol. Ecol. 2007, 59, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Danzeisen, J.L.; Kim, H.B.; Isaacson, R.E.; Tu, Z.J.; Johnson, T.J. Modulations of the chicken cecal microbiome and metagenome in response to anticoccidial and growth promoter treatment. PLoS ONE 2011, 6, e27949. [Google Scholar] [CrossRef]
- Stanley, D.; Denman, S.E.; Hughes, R.J.; Geier, M.S.; Crowley, T.M.; Chen, H.; Haring, V.R.; Moore, R.J. Intestinal microbiota associated with differential feed conversion efficiency in chickens. Appl. Microbiol. Biotechnol. 2012, 96, 1361–1369. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.C.; Bessegatto, J.A.; Alfieri, A.A.; Weese, J.S.; Filho, J.A.; Oba, A. Different antibiotic growth promoters induce specific changes in the cecal microbiota membership of broiler chicken. PLoS ONE 2017, 12, e0171642. [Google Scholar] [CrossRef] [PubMed]
- Józefiak, A.; Benzertiha, A.; Kierończyk, B.; Łukomska, A.; Wesołowska, I.; Rawski, M. Improvement of Cecal Commensal Microbiome Following the Insect Additive into Chicken Diet. Animals 2020, 10, 577. [Google Scholar] [CrossRef] [Green Version]
- Borrelli, L.; Coretti, L.; Dipineto, L.; Bovera, F.; Menna, F.; Chiariotti, L.; Nizza, A.; Lembo, F.; Fioretti, A. Insect-based diet, a promising nutritional source, modulates gut microbiota composition and SCFAs production in laying hens. Sci. Rep. 2017, 7, 16269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ducatelle, R.; Goossens, E.; De Meyer, F.; Eeckhaut, V.; Antonissen, G.; Haesebrouck, F.; Van Immerseel, F. Biomarkers for monitoring intestinal health in poultry: Present status and future perspectives. Vet. Res. 2018, 49, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Singh, K.M.; Shah, T.; Deshpande, S.; Jakhesara, S.J.; Koringa, P.G.; Rank, D.N.; Joshi, C.G. High through put 16S rRNA gene-based pyrosequencing analysis of the fecal microbiota of high FCR and low FCR broiler growers. Mol. Biol. Rep. 2012, 39, 10595–10602. [Google Scholar] [CrossRef]
- Pryde, S.E.; Duncan, S.H.; Hold, G.L.; Stewart, C.S.; Flint, H.J. The microbiology of butyrate formation in the human colon. FEMS. Microbiol. Lett. 2002, 217, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, S.; Ding, J.; Dai, R.; He, C.; Xu, K.; Honaker, C.F.; Zhang, Y.; Siegel, P.; Meng, H. Gut microbiota co-microevolution with selection for host humoral immunity. Front. Microbiol. 2017, 8, 1243. [Google Scholar] [CrossRef] [Green Version]
- Onrust, L.; Ducatelle, R.; Van Driessche, K.; De Maesschalck, C.; Vermeulen, K.; Haesebrouck, F.; Eeckhaut, V.; Van Immerseel, F. Steering endogenous butyrate production in the intestinal tract of broilers as a tool to improve gut health. Front. Vet. Sci. 2015, 2, 75. [Google Scholar] [CrossRef]
- Liu, C.; Finegold, S.M.; Song, Y.; Lawson, P.A. Reclassification of Clostridium coccoides, Ruminococcus hansenii, Ruminococcus hydrogenotrophicus, Ruminococcus luti, Ruminococcus productus and Ruminococcus schinkii as Blautia coccoides gen. nov., comb. nov., Blautia hansenii comb. nov., Blautia hydrogenotrophica comb. nov., Blautia luti comb. nov., Blautia producta comb. nov., Blautia schinkii comb. nov. and description of Blautia wexlerae sp. nov., isolated from human faeces. Int. J. Syst. Evol. Microbiol. 2008, 58, 1896–1902. [Google Scholar]
- Sunkara, L.T.; Jiang, W.; Zhang, G. Modulation of antimicrobial host defense peptide gene expression by free fatty acids. PLoS ONE 2012, 7, e49558. [Google Scholar] [CrossRef] [Green Version]
- Rehman, H.U.; Vahjen, W.; Awad, W.A.; Zentek, J. Indigenous bacteria and bacterial metabolic products in the gastrointestinal tract of broiler chickens. Arch. Anim. Nutr. 2007, 61, 319–335. [Google Scholar] [CrossRef]
- Forder, R.E.; Howarth, G.S.; Tivey, D.R.; Hughes, R.J. Bacterial modulation of small intestinal goblet cells and mucin composition during early posthatch development of poultry. Poult. Sci. 2007, 86, 2396–2403. [Google Scholar] [CrossRef]
- Forstner, G.; Forstner, J.F. Gastrointestinal mucus. In Physiology of the Gastrointestinal Tract, 1st ed.; Johnson, L.R., Ed.; Raven Press: New York, NY, USA, 1994; pp. 1255–1284. [Google Scholar]
- Corfield, A.P.; Wagner, S.A.; Clamp, J.R.; Kriaris, M.S.; Hoskins, L.C. Mucin degradation in the human colon: Production of sialidase, sialate O-acetylesterase, N-acetylneuraminate lyase, arylesterase, and glycosulfatase activities by strains of fecal bacteria. Infect. Immun. 1992, 60, 3971–3978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turck, D.; Feste, A.; Lifschitz, C. Age and diet affect the composition of porcine colonic mucins. Pediatr. Res. 1993, 33, 564–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | d.f. 6 | Chi-Square | p7 |
|---|---|---|---|
| Crypts | |||
| Diet 1 | 3 | 12.388 | 0.006 |
| Mucin type 2 | 2 | 18.860 | <0.001 |
| Gut segment 3 | 3 | 75.407 | <0.001 |
| Fragment 4 | 2 | 96.076 | <0.001 |
| Villi | |||
| Diet | 3 | 4.045 | 0.257 |
| Mucin type | 2 | 4.937 | 0.085 |
| Gut segment 5 | 2 | 748.764 | <0.001 |
| Fragment | 2 | 43.135 | <0.001 |
| Mucosal Element | Predictor | Predictor Factors | Mucin Staining Intensity 1,2 |
|---|---|---|---|
| Crypts | Diet | C | 0.98 ± 0.02 ab |
| TM5 | 1.04 ± 0.03 a | ||
| TM10 | 0.94 ± 0.02 b | ||
| TM15 | 0.93 ± 0.02 b | ||
| Mucin type | Neutral | 0.93 ± 0.02 b | |
| Acidic sialylated | 1.05 ± 0.02 a | ||
| Acidic sulfated | 0.96 ± 0.02 b | ||
| Gut segment | Duodenum | 1.09 ± 0.03 a | |
| Jejunum | 0.99 ± 0.02 b | ||
| Ileum | 1.03 ± 0.02 ab | ||
| Cecum | 0.82 ± 0.02 c | ||
| Fragment | Base | 1.15 ± 0.02 a | |
| Midsection | 0.89 ± 0.02 b | ||
| Tip | 0.91 ± 0.09 b | ||
| Villi | Diet | C | 1.70 ± 0.04 |
| TM5 | 1.69 ± 0.04 | ||
| TM10 | 1.59 ± 0.04 | ||
| TM15 | 1.67 ± 0.04 | ||
| Mucin type | Neutral | 1.68 ± 0.04 ab | |
| Acidic sialylated | 1.71 ± 0.04 a | ||
| Acidic sulfated | 1.60 ± 0.04 b | ||
| Gut segment | Duodenum | 1.01 ± 0.02 c | |
| Jejunum | 1.96 ± 0.04 b | ||
| Ileum | 2.31 ± 0.05 a | ||
| Fragment | Base | 1.80 ± 0.04 a | |
| Midsection | 1.71 ± 0.04 a | ||
| Tip | 1.49 ± 0.03 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biasato, I.; Ferrocino, I.; Grego, E.; Dabbou, S.; Gai, F.; Gasco, L.; Cocolin, L.; Capucchio, M.T.; Schiavone, A. Yellow Mealworm Inclusion in Diets for Heavy-Size Broiler Chickens: Implications for Intestinal Microbiota and Mucin Dynamics. Animals 2020, 10, 1909. https://doi.org/10.3390/ani10101909
Biasato I, Ferrocino I, Grego E, Dabbou S, Gai F, Gasco L, Cocolin L, Capucchio MT, Schiavone A. Yellow Mealworm Inclusion in Diets for Heavy-Size Broiler Chickens: Implications for Intestinal Microbiota and Mucin Dynamics. Animals. 2020; 10(10):1909. https://doi.org/10.3390/ani10101909
Chicago/Turabian StyleBiasato, Ilaria, Ilario Ferrocino, Elena Grego, Sihem Dabbou, Francesco Gai, Laura Gasco, Luca Cocolin, Maria Teresa Capucchio, and Achille Schiavone. 2020. "Yellow Mealworm Inclusion in Diets for Heavy-Size Broiler Chickens: Implications for Intestinal Microbiota and Mucin Dynamics" Animals 10, no. 10: 1909. https://doi.org/10.3390/ani10101909
APA StyleBiasato, I., Ferrocino, I., Grego, E., Dabbou, S., Gai, F., Gasco, L., Cocolin, L., Capucchio, M. T., & Schiavone, A. (2020). Yellow Mealworm Inclusion in Diets for Heavy-Size Broiler Chickens: Implications for Intestinal Microbiota and Mucin Dynamics. Animals, 10(10), 1909. https://doi.org/10.3390/ani10101909