Detection of Novel Goose Parvovirus Disease Associated with Short Beak and Dwarfism Syndrome in Commercial Ducks



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Samples and Samples’ Preparation
2.3. Polymerase Chain Reaction (PCR)
2.3.1. Viral DNA Purification
2.3.2. PCR Amplification
2.4. DNA Sequence
2.5. Virus Isolation
3. Results
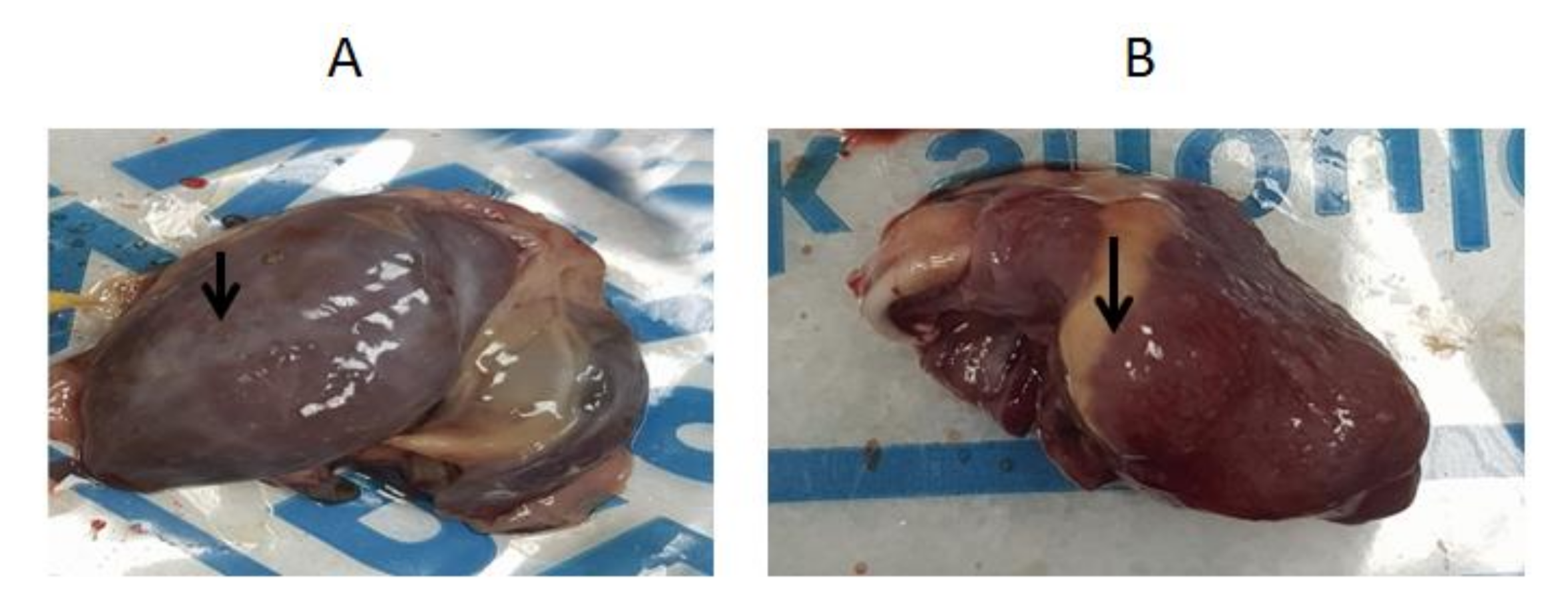
3.1. Clinical Picture and Postmortem Findings
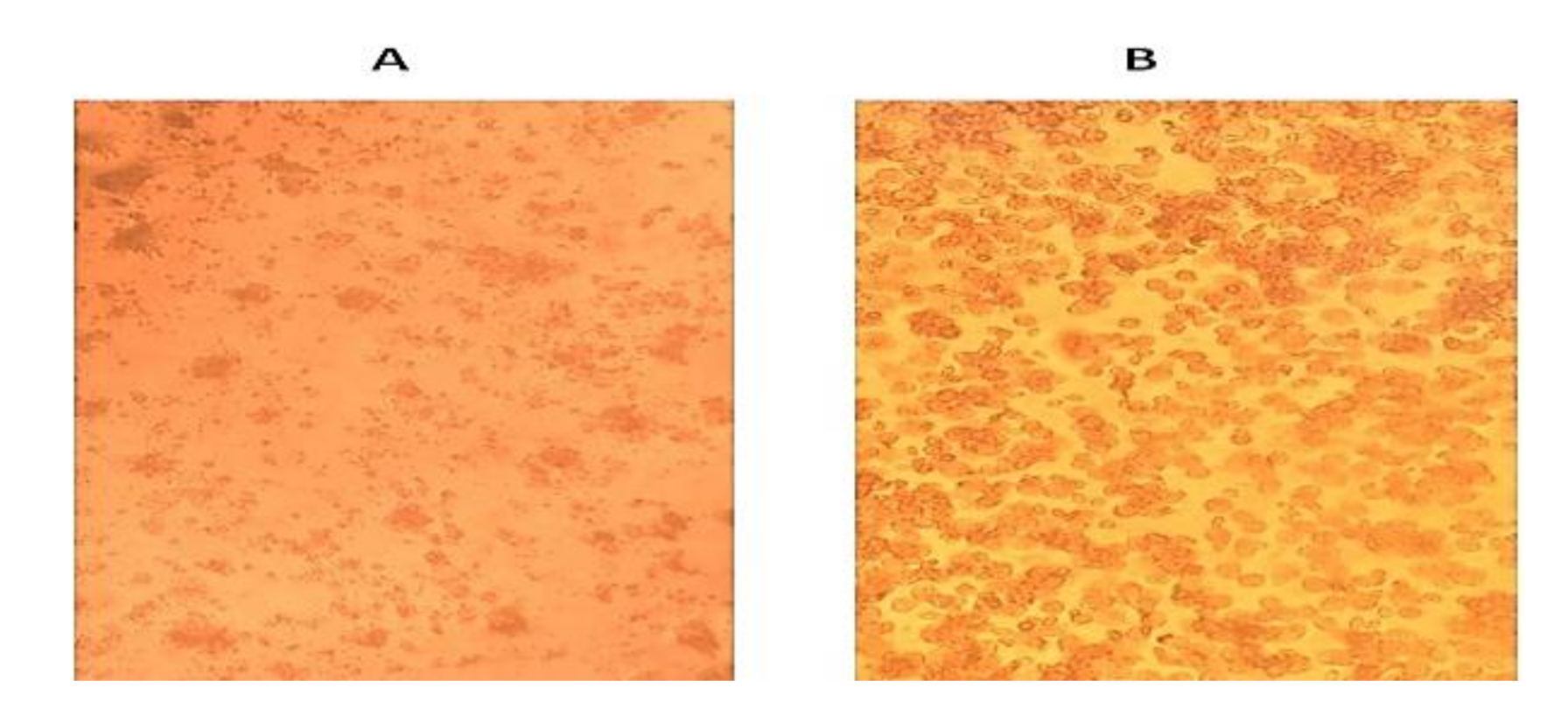
3.2. Identification and Isolation of the Causative Agent
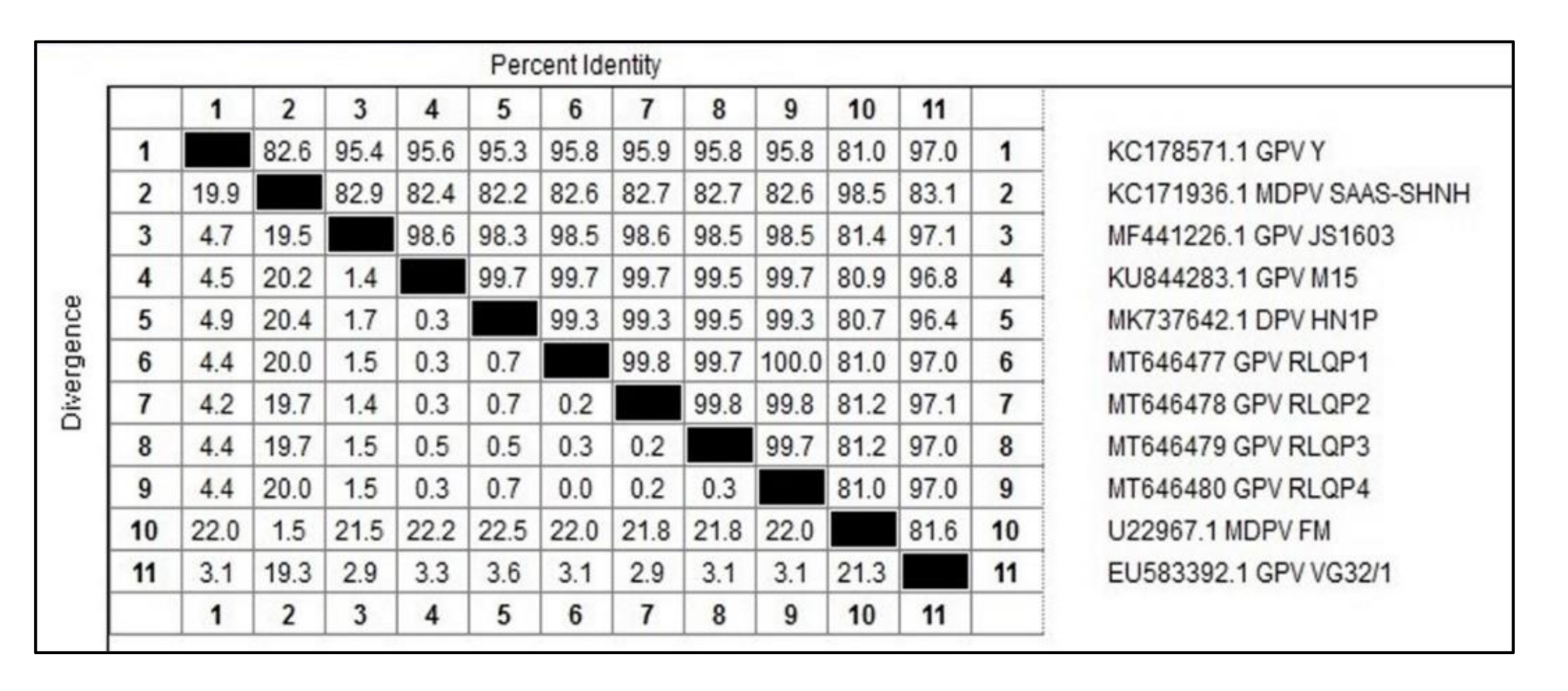
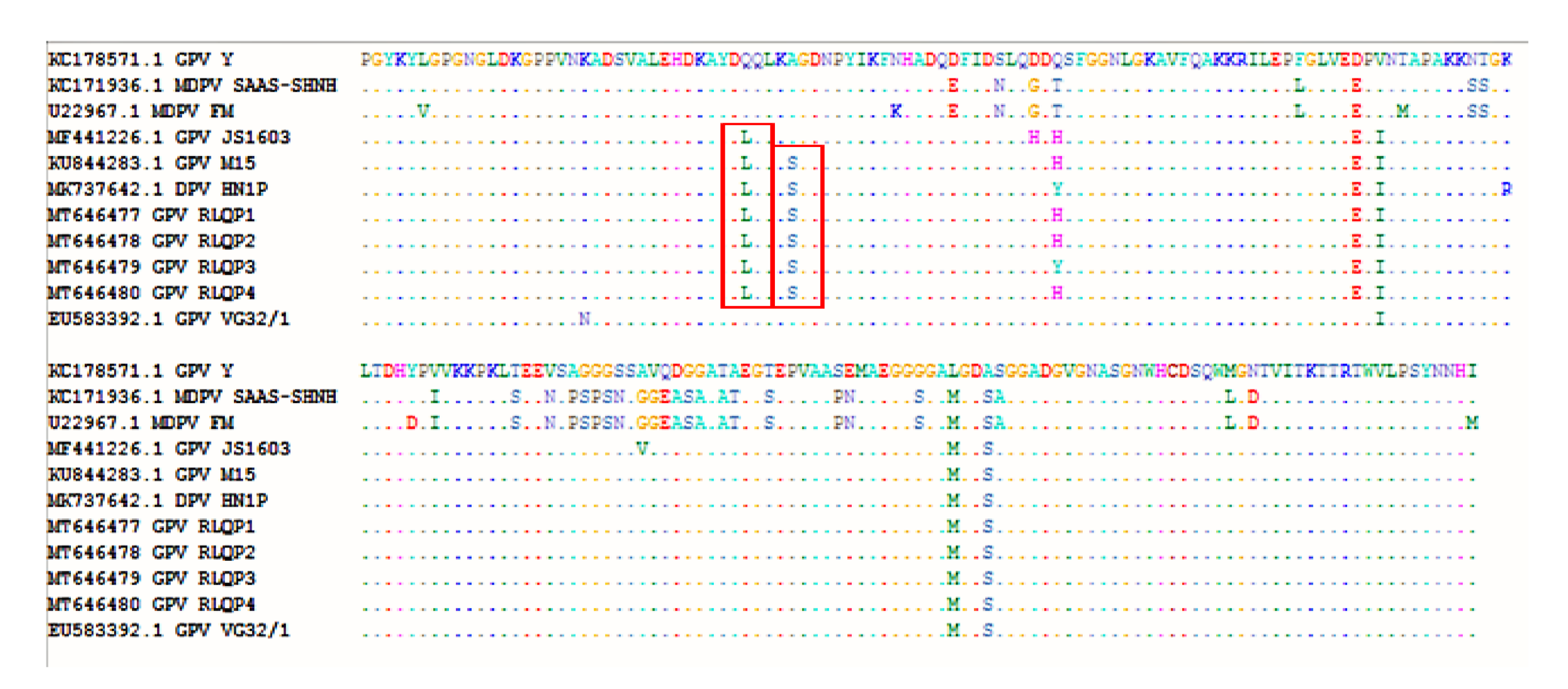
3.3. DNA Sequence and Phylogenetic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Derzsy, D. A viral disease of goslings. I. Epidemiological, clinical, pathological and aetiological studies. Acta Vet. Acad. Sci. Hung. 1967, 17, 443–448. [Google Scholar] [PubMed]
- Palya, V.J. Parvovirus infections of water fowl. In Diseases of Poultry, 13th ed.; Ames, I.A., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2013; pp. 444–451. [Google Scholar]
- Chang, P.C.; Shien, J.H.; Wang, M.S.; Shieh, H.K. Phylogenetic analysis of parvoviruses isolated in Taiwan from ducks and geese. Avian Pathol. 2000, 29, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Le Gall-Reculé, G.; Jestin, V. Biochemical and genomic characterization of muscovy duck parvovirus. Arch. Virol. 1994, 139, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Zádori, Z.; Stefancsik, R.; Rauch, T.; Kisary, J. Analysis of the complete nucleotide sequences of goose and muscovy duck parvoviruses indicates common ancestral origin with adeno-associated virus 2. Virology 1995, 212, 562–573. [Google Scholar] [CrossRef] [PubMed]
- Takehara, K.; Hyakutake, K.; Imamura, T.; Mutoh, K.; Yoshimura, M. Isolation, identification, and plaque titration of parvovirus from Muscovy ducks in Japan. Avian Dis. 1994, 38, 810–815. [Google Scholar] [CrossRef] [PubMed]
- Glávits, R.; Zolnai, A.; Szabó, E.; Ivanics, E.; Zarka, P.; Mató, T.; Palya, V. Comparative pathological studies on domestic geese (Anseranser domestica) and Muscovy ducks (Cairina moschata) experimentally infected with parvovirus strains of goose and Muscovy duck origin. Acta. Vet. Hung. 2005, 53, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Jansson, D.S.; Feinstein, R.; Kardi, V.; Mató, T.; Palya, V. Epidemiologic investigation of an outbreak of goose parvovirus infection in Sweden. Avian Dis. 2007, 51, 609–613. [Google Scholar] [CrossRef]
- Woźniakowski, G.; Samorek-Salamonowicz, E.; Kozdruń, W. Quantitative analysis of waterfowl parvoviruses in geese and Muscovy ducks by real-time polymerase chain reaction: Correlation between age, clinical symptoms and DNA copy number of waterfowl parvoviruses. BMC Vet. Res. 2012, 8, 1–10. [Google Scholar] [CrossRef]
- Wan, C.; Chen, C.; Cheng, L.; Chen, H.; Fu, Q.; Shi, S.; Fu, G.; Liu, R.; Huang, Y. Specific detection of Muscovy duck parvovirus infection by TaqMan-based real-time PCR assay. BMC Vet. Res. 2018, 14, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Poonia, B.; Dunn, P.A.; Lu, H.; Jarosinski, K.W.; Schat, K.A. Isolation and molecular characterization of a new Muscovy duck parvovirus from Muscovy ducks in the USA. Avian Pathol. 2006, 35, 435–441. [Google Scholar] [CrossRef]
- Shien, J.H.; Wang, Y.S.; Chen, C.H.; Shieh, H.K.; Hu, C.C.; Chang, P.C. Identification of sequence changes in live attenuated goose parvovirus vaccine strains developed in Asia and Europe. Avian Pathol. 2008, 37, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Palya, V.; Zolnai, A.; Benyeda, Z.; Kovács, E.; Kardi, V.; Mató, T. Short beak and dwarfism syndrome of mule duck is caused by a distinct lineage of goose parvovirus. Avian Pathol. 2009, 38, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.; Ma, X.; Sheng, Z.; Qi, L.; Liu, C.; Wang, D.; Huang, B.; Li, F.; Song, M. Identification of Goose-Origin Parvovirus as a Cause of Newly Emerging Beak Atrophy and Dwarfism Syndrome in Ducklings. J. Clin. Microbiol. 2016, 54, 1999–2007. [Google Scholar] [CrossRef]
- Lu, Y.S.; Lin, D.F.; Lee, Y.L.; Liao, Y.K.; Tsai, H.J. Infectious bill atrophy syndrome caused by parvovirus in a co-outbreak with duck viral hepatitis in ducklings in Taiwan. Avian Dis. 1993, 37, 591–596. [Google Scholar] [CrossRef]
- Li, C.; Li, Q.; Chen, Z.; Liu, G. Novel duck parvovirus identified in Cherry Valley ducks (Anas platyrhynchos domesticus), China. Infect. Genet. Evol. 2016, 44, 278–280. [Google Scholar] [CrossRef] [PubMed]
- Ning, K.; Wang, M.; Qu, S.; Lv, J.; Yang, L.; Zhang, D. Pathogenicity of Pekin duck- and goose-origin parvoviruses in Pekin ducklings. Vet. Microbiol. 2017, 210, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Sui, N.; Zhang, R.; Lan, J.; Li, P.; Lian, C.; Li, H.; Xie, Z.; Jiang, S. Coinfection of novel goose parvovirus-associated virus and duck circovirus in feather sacs of Cherry Valley ducks with feathers shedding syndrome. Poult. Sci. 2020, 99, 4227–4234. [Google Scholar] [CrossRef] [PubMed]
- Irvine, R.; Holmes, P. Diagnosis and control of goose parvovirus. Practice 2010, 32, 382–386. [Google Scholar] [CrossRef]
- Fan, W.; Sun, Z.; Shen, T.; Xu, D.; Huang, K.; Zhou, J.; Song, S.; Yan, L. Analysis of Evolutionary Processes of Species Jump in Waterfowl Parvovirus. Front. Microbiol. 2017, 8, 1–9. [Google Scholar] [CrossRef]
- Chen, H.; Tang, Y.; Dou, Y.; Zheng, X.; Diao, Y. Evidence for Vertical Transmission of Novel Duck-Origin Goose Parvovirus-Related Parvovirus. Transbound. Emerg. Dis. 2016, 63, 243–247. [Google Scholar] [CrossRef]
- Wan, C.; Chen, C.; Cheng, L.; Liu, R.; Shi, S.; Fu, G.; Chen, H.; Fu, Q.; Huang, Y. Specific detection and differentiation of classic goose parvovirus and novel goose parvovirus by TaqMan real-time PCR assay, coupled with host specificity. BMC Vet. Res. 2019, 15, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Wang, S.; Cheng, X.; Xiao, S.; Zhu, X.; Lin, F.; Wu, N.; Wang, J.; Huang, M.; Zheng, M.; et al. Isolation and characterization of a distinct duck-origin goose parvovirus causing an outbreak of duckling short beak and dwarfism syndrome in China. Arch. Virol. 2016, 161, 2407–2416. [Google Scholar] [CrossRef] [PubMed]
- Salganik, M.; Aydemir, F.; Nam, H.J.; McKenna, R.; Agbandje-McKenna, M.; Muzyczka, N. Adeno-associated virus capsid proteins may play a role in transcription and second-strand synthesis of recombinant genomes. J. Virol. 2014, 88, 1071–1079. [Google Scholar] [CrossRef] [PubMed]
- Naguib, M.M.; Verhagen, J.H.; Samy, A.; Eriksson, P.; Fife, M.; Lundkvist, Å.; Ellström, P.; Järhult, J.D. Avian influenza viruses at the wild-domestic bird interface in Egypt. Infect. Ecol. Epidemiol. 2019, 9, 1575687. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic. Acids. Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Gough, R.E. Goose parvovirus (Derzsy’s disease). In A Laboratory Manual for the Isolation and Identification of Avian Pathogens, 4th ed.; American Association of Avian Pathologists, University of Pennsylvania: Philadelphia, PA, USA, 1998; pp. 219–223. [Google Scholar]
- Woźniakowski, G.; Kozdruń, W.; Samorek-Salamonowicz, E. Genetic variance of Derzsy’s disease strains isolated in Poland. J. Mol. Genet. Med. 2009, 3, 210–216. [Google Scholar] [CrossRef]
- Jin, M.; Feng, C.; Wang, X.; Zhang, D. Molecular evidence of goose-parvovirus-related abnormal molting in Pekin ducks. Arch. Virol. 2019, 164, 2837–2841. [Google Scholar] [CrossRef]
- Kapgate, S.S.; Kumanan, K.; Vijayarani, K.; Barbuddhe, S.B. Avian parvovirus: Classification, phylogeny, pathogenesis and diagnosis. Avian Pathol. 2018, 47, 536–545. [Google Scholar] [CrossRef]
- Li, P.; Lin, S.; Zhang, R.; Chen, J.; Sun, D.; Lan, J.; Song, S.; Xie, Z.; Jiang, S. Isolation and characterization of novel goose parvovirus-related virus reveal the evolution of waterfowl parvovirus. Transbound. Emerg. Dis. 2018, 65, 284–295. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Zhang, R.; Chen, J.; Sun, D.; Lan, J.; Lin, S.; Song, S.; Xie, Z.; Jiang, S. Development of a duplex semi-nested PCR assay for detection of classical goose parvovirus and novel goose parvovirus-related virus in sick or dead ducks with short beak and dwarfism syndrome. J. Virol. Methods 2017, 249, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Bian, G.; Ma, H.; Luo, M.; Gong, F.; Li, B.; Wang, G.; Mohiuddin, M.; Liao, M.; Yuan, J. Identification and genomic analysis of two novel duck-origin GPV-related parvovirus in China. BMC Vet. Res. 2019, 15, 88. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.; Chen, S.; Cheng, X.; Lin, F.; Wang, S.; Zhu, X.; Yu, B.; Huang, M.; Wang, J.; Wu, N.; et al. The newly emerging duck-origin goose parvovirus in China exhibits a wide range of pathogenicity to main domesticated waterfowl. Vet. Microbiol. 2017, 203, 252–256. [Google Scholar] [CrossRef] [PubMed]
- Berns, K.I. Parvovirus replication. Microbiol. Rev. 1990, 54, 316–329. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Erfan, A.M.; Selim, A.A.; Moursi, M.K.; Nasef, S.A.; Abdelwhab, E.M. Epidemiology and molecular characterisation of duck hepatitis A virus from different duck breeds in Egypt. Vet. Microbiol. 2015, 177, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Selim, A.A.; Erfan, A.M.; Hagag, N.; Zanaty, A.; Samir, A.H.; Samy, M.; Abdelhalim, A.; Arafa, A.A.; Soliman, M.A.; Shaheen, M.; et al. Highly Pathogenic Avian Influenza Virus (H5N8) Clade 2.3.4.4 Infection in Migratory Birds, Egypt. Emerg. Infect. Dis. 2017, 23, 1048–1051. [Google Scholar] [CrossRef]
- Zanaty, A.M.; Erfan, A.M.; Mady, W.H.; Amer, F.; Nour, A.A.; Rabie, N.; Samy, M.; Selim, A.A.; Hassan, W.M.M.; Naguib, M.M. Avian influenza virus surveillance in migratory birds in Egypt revealed a novel reassortant H6N2 subtype. Avian Res. 2019, 10, 41–50. [Google Scholar] [CrossRef]
- Rossmann, M.G. The canyon hypothesis. Hiding the host cell receptor attachment site on a viral surface from immune surveillance. J. Biol. Chem. 1989, 264, 14587–14590. [Google Scholar]
- Tsao, J.; Chapman, M.S.; Agbandje, M.; Keller, W.; Smith, K.; Wu, H.; Luo, M.; Smith, T.J.; Rossmann, M.G.; Compans, R.W.; et al. The three-dimensional structure of canine parvovirus and its functional implications. Science 1991, 251, 1456–1464. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soliman, M.A.; Erfan, A.M.; Samy, M.; Mahana, O.; Nasef, S.A. Detection of Novel Goose Parvovirus Disease Associated with Short Beak and Dwarfism Syndrome in Commercial Ducks. Animals 2020, 10, 1833. https://doi.org/10.3390/ani10101833
Soliman MA, Erfan AM, Samy M, Mahana O, Nasef SA. Detection of Novel Goose Parvovirus Disease Associated with Short Beak and Dwarfism Syndrome in Commercial Ducks. Animals. 2020; 10(10):1833. https://doi.org/10.3390/ani10101833
Chicago/Turabian StyleSoliman, Mohamed A., Ahmed M. Erfan, Mohamed Samy, Osama Mahana, and Soad A. Nasef. 2020. "Detection of Novel Goose Parvovirus Disease Associated with Short Beak and Dwarfism Syndrome in Commercial Ducks" Animals 10, no. 10: 1833. https://doi.org/10.3390/ani10101833
APA StyleSoliman, M. A., Erfan, A. M., Samy, M., Mahana, O., & Nasef, S. A. (2020). Detection of Novel Goose Parvovirus Disease Associated with Short Beak and Dwarfism Syndrome in Commercial Ducks. Animals, 10(10), 1833. https://doi.org/10.3390/ani10101833