The Overexpression of Tβ4 in the Hair Follicle Tissue of Alpas Cashmere Goats Increases Cashmere Yield and Promotes Hair Follicle Development

 ,
,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Design and Construction of the Vectors
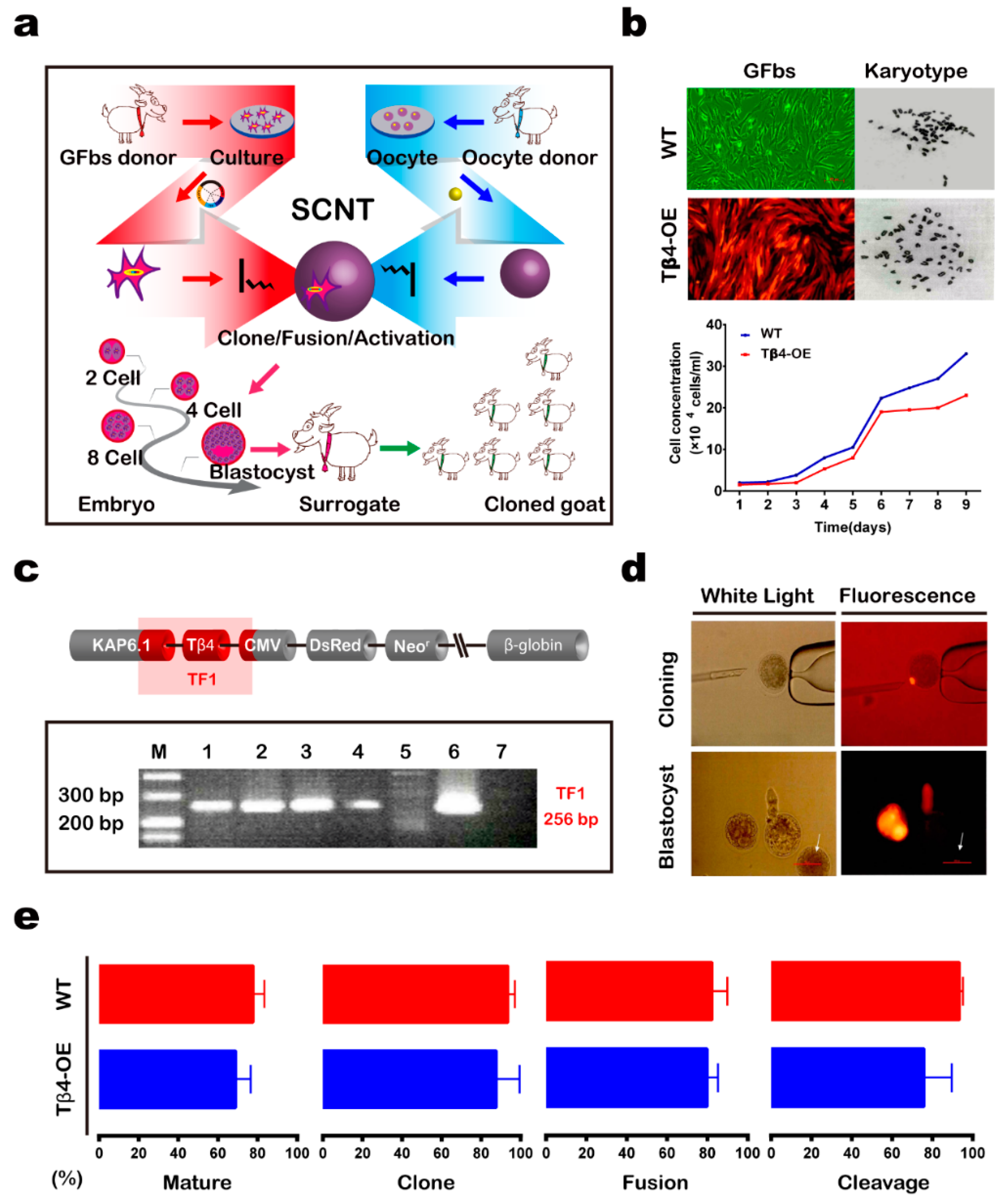
2.3. Cell Culture, Transfection, and Selection
2.4. Karyotype Analysis
2.5. Cell Growth Curves
2.6. Generation of Cloned Goats via SCNT
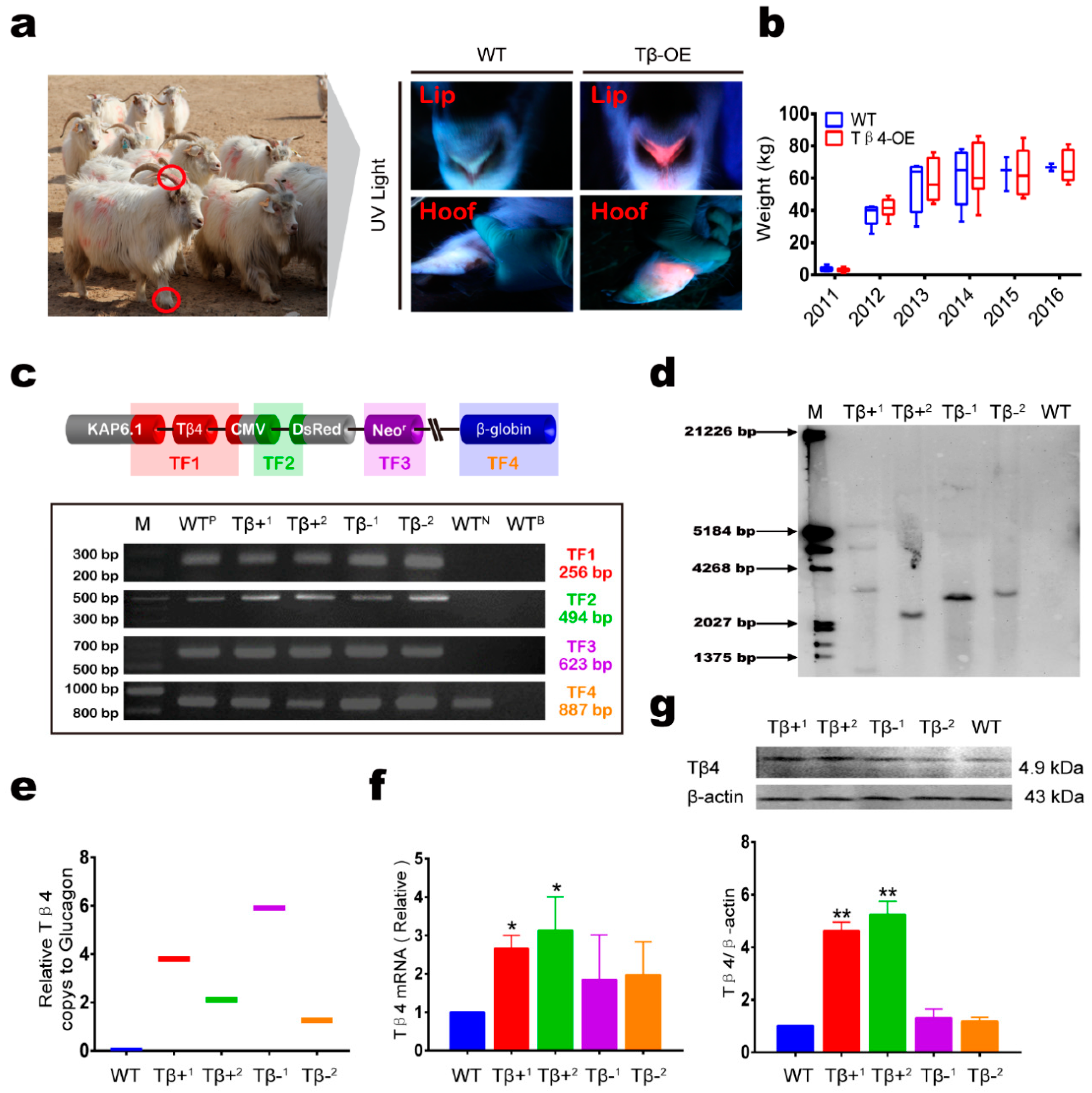
2.7. PCR Analysis
2.8. Southern Blot Analysis
2.9. DNA Extraction and qRT-PCR
2.10. RNA Extraction and qRT-PCR
2.11. Western Blot Analysis
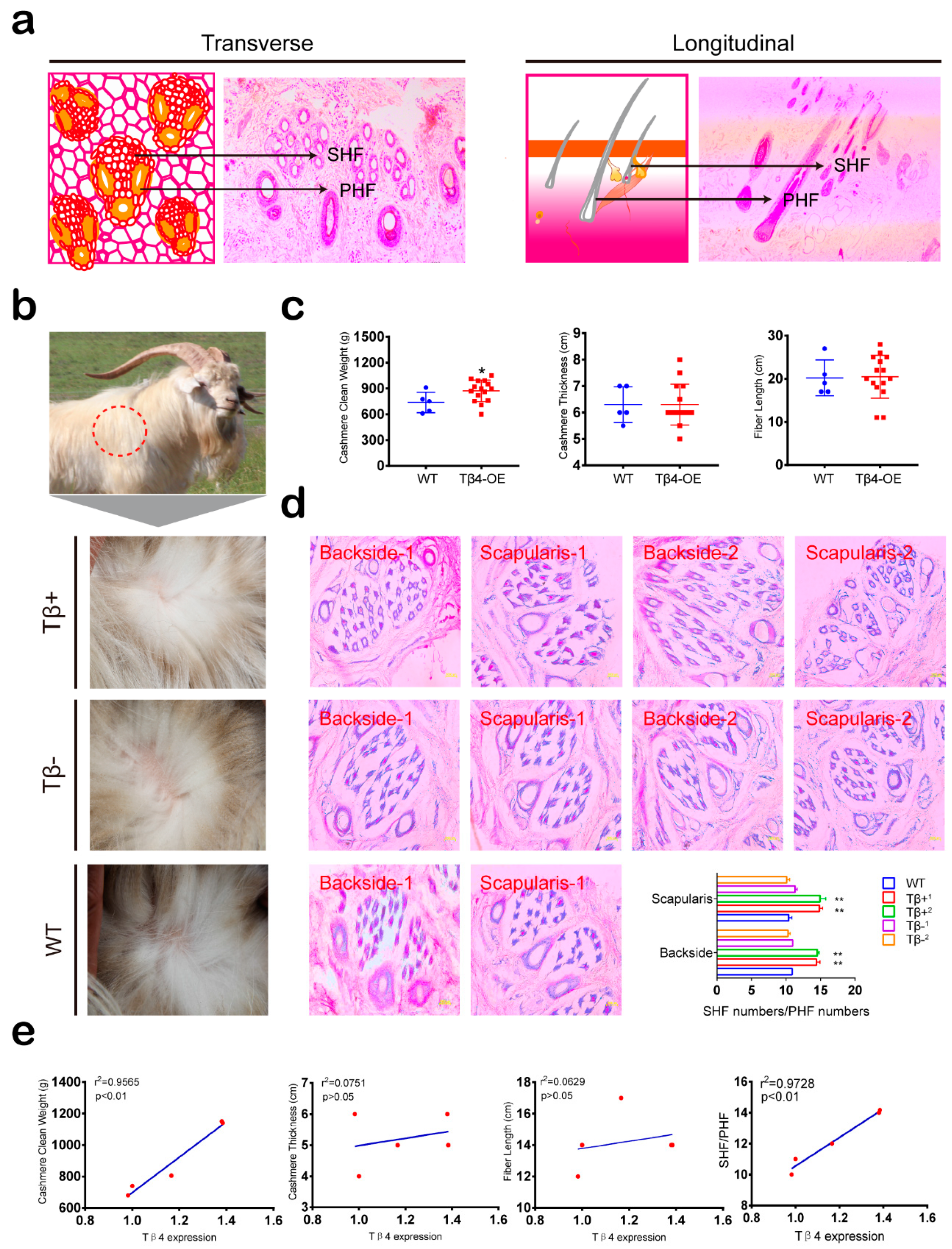
2.12. Hematoxylin-Eosin (H&E) Staining
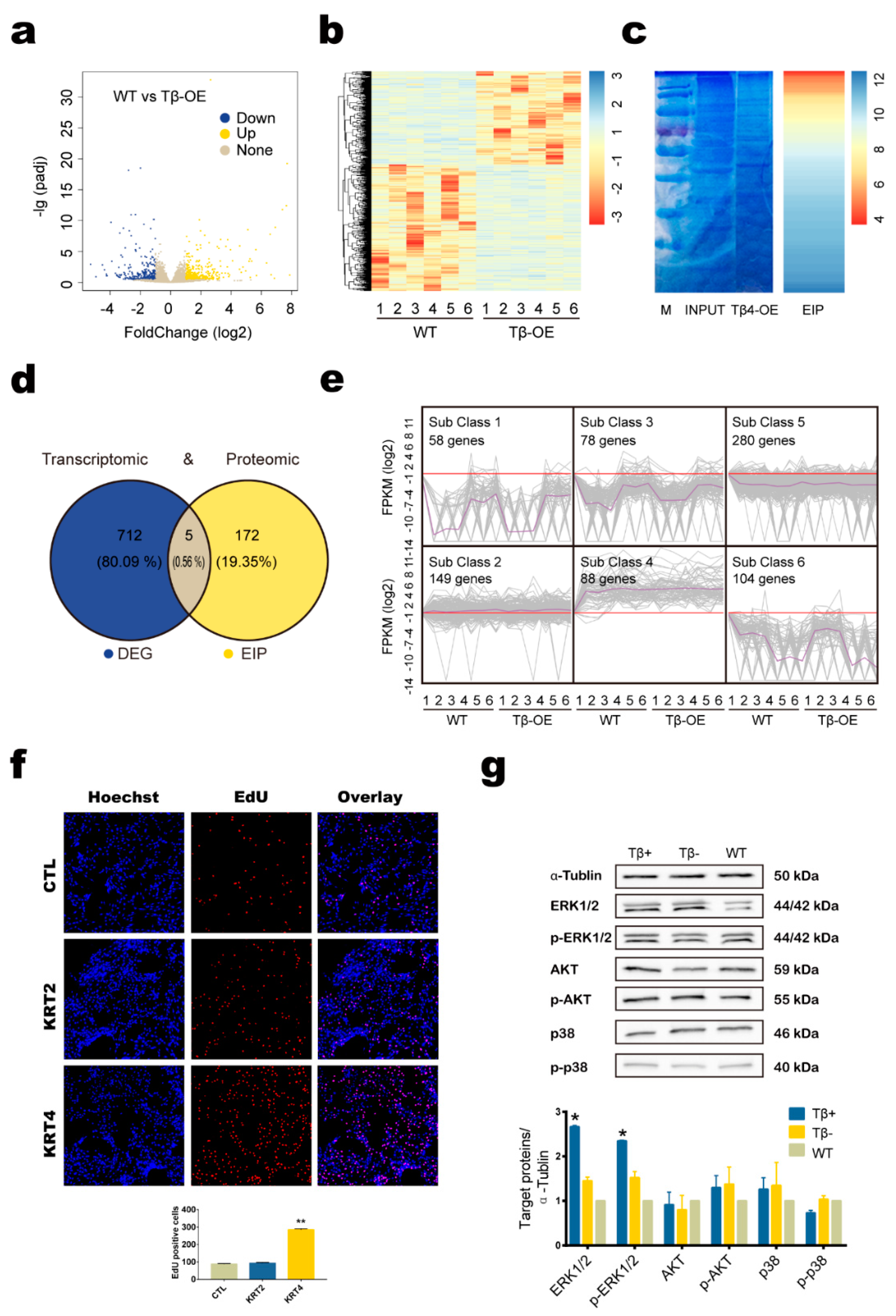
2.13. RNA Sequencing Alignment and Transcriptomic Analysis
2.14. Co-Immunoprecipitation (Co-IP)
2.15. Protein Mass Spectrometry (MS) Analysis
2.16. Cell Proliferation Assay
2.17. Animal Management and Measurements
3. Results
3.1. Generation of Tβ4-OE Alpas Cashmere Goats
3.2. Ectopic Expression of Tβ4 in Tβ4-OE Alpas Cashmere Goats
3.3. Cashmere Yield and SHF/PHF Ratio Increased in Tβ4-OE Alpas Cashmere Goats
3.4. Tβ4 May Affect Hair Growth by Interacting with KRT4 to Mediate ERK Signaling Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zeder, M.A.; Hesse, B. The initial domestication of goats (capra hircus) in the zagros mountains 10,000 years ago. Science 2000, 287, 2254–2257. [Google Scholar] [CrossRef] [PubMed]
- Bai, W.L. Multivariate statistic analysis of morphological and ecological characters of cashmere goat populations in china. J. Anhui Agric. Sci. 2006, 34, 489. [Google Scholar]
- Xue-Feng, L.V.; Zheng, W.X. Research progress and perspective in skin follicle of cashmere goat. China Anim. Husb. Vet. Med. 2010, 37, 25–28. [Google Scholar]
- Bai, J.; Zhang, Q.; Li, J.; Dao, E.J.; Jia, X. Estimates of genetic parameters and genetic trends for production traits of inner mongolian white cashmere goat. Asian-Australas. J. Anim. Sci. 2005, 19, 15–18. [Google Scholar] [CrossRef]
- MacHugh, D.E.; Bradley, D.G. Livestock genetic origins: Goats buck the trend. Proc. Natl. Acad. Sci. USA 2001, 98, 5382–5384. [Google Scholar] [CrossRef] [PubMed]
- Mao, C.; Xu, Y.; Shi, L.; Guo, S.; Jin, X.; Yan, S.; Shi, B. Effects of photoperiod change on melatonin secretion, immune function and antioxidant status of cashmere goats. Animals (Basel) 2019, 9, 766. [Google Scholar] [CrossRef]
- Berger, J.; Buuveibaatar, B.; Mishra, C. Globalization of the cashmere market and the decline of large mammals in central Asia. Conserv. Biol. 2013, 27, 679–689. [Google Scholar] [CrossRef]
- Wu, G.; He, Y. Identification of Varieties of Cashmere by Vis/Nir Spectroscopy Technology Based on Pca-Svm; IEEE: Piscataway, NJ, USA, 2009; Volume 29, pp. 1541–1544. [Google Scholar]
- Bai, D.P.; Yang, M.M.; Qu, L.; Chen, Y.L. Generation of a transgenic cashmere goat using the piggybac transposition system. Theriogenology 2017, 93, 1–6. [Google Scholar] [CrossRef]
- Li, X.; Hao, F.; Hu, X.; Wang, H.; Dai, B.; Wang, X.; Liang, H.; Cang, M.; Liu, D. Generation of tbeta4 knock-in cashmere goat using crispr/cas9. Int. J. Biol. Sci. 2019, 15, 1743–1754. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, J.; Yang, W.; Cui, M.; Dai, B.; Dong, Y.; Yang, J.; Zhang, X.; Liu, D.; Liang, H.; et al. Comparison of gene editing efficiencies of crispr/cas9 and talen for generation of mstn knock-out cashmere goats. Theriogenology 2019, 132, 1–11. [Google Scholar] [CrossRef]
- Wang, X.; Cai, B.; Zhou, J.; Zhu, H.; Niu, Y.; Ma, B.; Yu, H.; Lei, A.; Yan, H.; Shen, Q.; et al. Disruption of fgf5 in cashmere goats using crispr/cas9 results in more secondary hair follicles and longer fibers. PLoS ONE 2016, 11, e0164640. [Google Scholar]
- Ge, W.; Wang, S.H.; Sun, B.; Zhang, Y.L.; Shen, W.; Khatib, H.; Wang, X. Melatonin promotes cashmere goat (capra hircus) secondary hair follicle growth: A view from integrated analysis of long non-coding and coding rnas. Cell Cycle 2018, 17, 1255–1267. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Song, S.; Dong, K.; Chen, X.; Liu, X.; Rouzi, M.; Zhao, Q.; He, X.; Pu, Y.; Guan, W.; et al. Skin transcriptome reveals the intrinsic molecular mechanisms underlying hair follicle cycling in cashmere goats under natural and shortened photoperiod conditions. Sci. Rep. 2017, 7, 13502. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Xie, M.; Jiang, Y.; Xiao, N.; Du, X.; Zhang, W.; Tosser-Klopp, G.; Wang, J.; Yang, S.; Liang, J.; et al. Sequencing and automated whole-genome optical mapping of the genome of a domestic goat (capra hircus). Nat. Biotechnol. 2012, 31, 135–141. [Google Scholar] [CrossRef]
- Rile, N.; Liu, Z.; Gao, L.; Qi, J.; Zhao, M.; Xie, Y.; Su, R.; Zhang, Y.; Wang, R.; Li, J.; et al. Expression of vimentin in hair follicle growth cycle of inner mongolian cashmere goats. BMC Genom. 2018, 19, 38. [Google Scholar] [CrossRef]
- Stenn, K.S.; Paus, R. Controls of hair follicle cycling. Physiol. Rev. 2001, 81, 449–494. [Google Scholar] [CrossRef]
- Bai, W.L.; Dang, Y.L.; Wang, J.J.; Yin, R.H.; Wang, Z.Y.; Zhu, Y.B.; Cong, Y.Y.; Xue, H.L.; Deng, L.; Guo, D.; et al. Molecular characterization, expression and methylation status analysis of bmp4 gene in skin tissue of liaoning cashmere goat during hair follicle cycle. Genetica 2016, 144, 457–467. [Google Scholar] [CrossRef]
- Huff, T.; Otto, A.M.; Muller, C.S.; Meier, M.; Hannappel, E. Thymosin beta4 is released from human blood platelets and attached by factor xiiia (transglutaminase) to fibrin and collagen. FASEB J. 2002, 16, 691–696. [Google Scholar] [CrossRef]
- Shi, B.; Ding, Q.; He, X.; Zhu, H.; Niu, Y.; Cai, B.; Cai, J.; Lei, A.; Kang, D.; Yan, H.; et al. Tbeta4-overexpression based on the piggybac transposon system in cashmere goats alters hair fiber characteristics. Transgenic Res. 2017, 26, 77–85. [Google Scholar] [CrossRef]
- Kumar, N.; Liao, T.D.; Romero, C.A.; Maheshwari, M.; Peterson, E.L.; Carretero, O.A. Thymosin beta4 deficiency exacerbates renal and cardiac injury in angiotensin-ii-induced hypertension. Hypertension 2018, 71, 1133–1142. [Google Scholar] [CrossRef]
- Philp, D.; Nguyen, M.; Scheremeta, B.; St-Surin, S.; Villa, A.M.; Orgel, A.; Kleinman, H.K.; Elkin, M. Thymosin beta4 increases hair growth by activation of hair follicle stem cells. FASEB J. 2004, 18, 385–387. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.Y.; Hou, F.; Zhang, Z.P.; Nuo, M.T.; Liang, H.; Cang, M.; Wang, Z.G.; Wang, X.; Xu, T.; Yan, L.Y.; et al. Role of thymosin beta 4 in hair growth. Mol. Genet. Genom. 2016, 291, 1639–1646. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Liang, H.; Hou, F.; Zhang, Z.; Nuo, M.; Guo, X.; Liu, D. Thymosin beta-4 induces mouse hair growth. PLoS ONE 2015, 10, e0130040. [Google Scholar] [CrossRef]
- Hajian, M.; Jafarpour, F.; Aghamiri, S.M.; Rouhollahi Varnosfaderani, S.; Nasr Esfahani, M.H. Effects of ovary storage temperature and embryo vitrification on somatic cell nuclear transfer outcomes in goats. Reprod. Fertil. Dev. 2019. [Google Scholar] [CrossRef]
- Burgstaller, J.P.; Brem, G. Aging of cloned animals: A mini-review. Gerontology 2017, 63, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Loi, P.; Iuso, D.; Czernik, M.; Ogura, A. A new, dynamic era for somatic cell nuclear transfer? Trends Biotechnol. 2016, 34, 791–797. [Google Scholar] [CrossRef]
- Yang, X.; Smith, S.L.; Tian, X.C.; Lewin, H.A.; Renard, J.P.; Wakayama, T. Nuclear reprogramming of cloned embryos and its implications for therapeutic cloning. Nat. Genet. 2007, 39, 295–302. [Google Scholar] [CrossRef]
- Matoba, S.; Zhang, Y. Somatic cell nuclear transfer reprogramming: Mechanisms and applications. Cell Stem Cell 2018, 23, 471–485. [Google Scholar] [CrossRef]
- Liu, Z.; Cai, Y.; Wang, Y.; Nie, Y.; Zhang, C.; Xu, Y.; Zhang, X.; Lu, Y.; Wang, Z.; Poo, M.; et al. Cloning of macaque monkeys by somatic cell nuclear transfer. Cell 2018, 172, 881–887. [Google Scholar] [CrossRef]
- Hao, F.; Yan, W.; Li, X.; Wang, H.; Wang, Y.; Hu, X.; Liu, X.; Liang, H.; Liu, D. Generation of cashmere goats carrying an edar gene mutant using crispr-cas9-mediated genome editing. Int. J. Biol. Sci. 2018, 14, 427–436. [Google Scholar] [CrossRef]
- Zhu, B.; Xu, T.; Yuan, J.; Guo, X.; Liu, D. Transcriptome sequencing reveals differences between primary and secondary hair follicle-derived dermal papilla cells of the cashmere goat (capra hircus). PLoS ONE 2013, 8, e76282. [Google Scholar] [CrossRef] [PubMed]
- Holm, P.; Booth, P.J.; Schmidt, M.H.; Greve, T.; Callesen, H. High bovine blastocyst development in a static in vitro production system using sofaa medium supplemented with sodium citrate and myo-inositol with or without serum-proteins. Theriogenology 1999, 52, 683–700. [Google Scholar] [CrossRef]
- Zhang, J.; Cui, M.L.; Nie, Y.W.; Dai, B.; Li, F.R.; Liu, D.J.; Liang, H.; Cang, M. Crispr/cas9-mediated specific integration of fat-1 at the goat mstn locus. FEBS J. 2018, 285, 2828–2839. [Google Scholar] [CrossRef] [PubMed]
- Siren, J.; Valimaki, N.; Makinen, V. Indexing graphs for path queries with applications in genome research. IEEE/ACM Trans. Comput. Biol. Bioinform. 2014, 11, 375–388. [Google Scholar] [CrossRef]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. Degseq: An r package for identifying differentially expressed genes from rna-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef]
- Wu, H.; Ren, Y.; Pan, W.; Dong, Z.; Cang, M.; Liu, D. The mammalian target of rapamycin signaling pathway regulates myocyte enhancer factor-2c phosphorylation levels through integrin-linked kinase in goat skeletal muscle satellite cells. Cell Biol. Int. 2015, 39, 1264–1273. [Google Scholar] [CrossRef]
- Nie, Y.; Liu, D. N-glycosylation is required for fdnc5 stabilization and irisin secretion. Biochem. J. 2017, 474, 3167–3177. [Google Scholar] [CrossRef]
- Philp, D.; St-Surin, S.; Cha, H.J.; Moon, H.S.; Kleinman, H.K.; Elkin, M. Thymosin beta 4 induces hair growth via stem cell migration and differentiation. Ann. N. Y. Acad. Sci. 2007, 1112, 95–103. [Google Scholar] [CrossRef]
- Lv, S.; Cheng, G.; Zhou, Y.; Xu, G. Thymosin beta4 induces angiogenesis through notch signaling in endothelial cells. Mol. Cell Biochem. 2013, 381, 283–290. [Google Scholar] [CrossRef]
- Jeon, B.J.; Yang, Y.; Kyung Shim, S.; Yang, H.M.; Cho, D.; Ik Bang, S. Thymosin beta-4 promotes mesenchymal stem cell proliferation via an interleukin-8-dependent mechanism. Exp. Cell Res. 2013, 319, 2526–2534. [Google Scholar] [CrossRef]
- Philp, D.; Goldstein, A.L.; Kleinman, H.K. Thymosin beta4 promotes angiogenesis, wound healing, and hair follicle development. Mech. Ageing Dev. 2004, 125, 113–115. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, V.; Panicker, A.G.; Indrakumar, S.; Chatterjee, K. Comparative study of keratin extraction from human hair. Int. J. Biol. Macromol. 2019, 133, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Li, W.; Deng, J.; Kan, J.; Guo, T.; Wang, B.; Hao, S. Recombinant human hair keratin nanoparticles accelerate dermal wound healing. ACS Appl. Mater. Interfaces 2019, 11, 18681–18690. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.L.; Lin, C.W.; Cheng, N.C.; Yang, K.C.; Yu, J. Modulation of keratin in adhesion, proliferation, adipogenic, and osteogenic differentiation of porcine adipose-derived stem cells. J. Biomed. Mater. Res. B Appl. Biomater. 2017, 105, 180–192. [Google Scholar] [CrossRef]
- Zhang, J.; Wu, J.; Zeng, W.; Yao, K.; Zu, H.; Zhao, Y. Function of thymosin beta-4 in ethanol-induced microglial activation. Cell Physiol. Biochem. 2016, 38, 2230–2238. [Google Scholar] [CrossRef]
- Chen, Y.; Tian, L.; Yang, F.; Tong, W.; Jia, R.; Zou, Y.; Yin, L.; Li, L.; He, C.; Liang, X.; et al. Tannic acid accelerates cutaneous wound healing in rats via activation of the erk 1/2 signaling pathways. Adv. Wound Care (New Rochelle) 2019, 8, 341–354. [Google Scholar] [CrossRef]
- Dai, B.; Zhang, M.; Yuan, J.L.; Ren, L.Q.; Han, X.Y.; Liu, D.J. Integrative analysis of methylation and transcriptional profiles to reveal the genetic stability of cashmere traits in the tbeta4 overexpression of cashmere goats. Animals (Basel) 2019, 9, 1002. [Google Scholar] [CrossRef]
- Saito, S.; Maeda, R.; Adachi, N. Dual loss of human polq and lig4 abolishes random integration. Nat. Commun. 2017, 8, 16112. [Google Scholar] [CrossRef]
- Iiizumi, S.; Kurosawa, A.; So, S.; Ishii, Y.; Chikaraishi, Y.; Ishii, A.; Koyama, H.; Adachi, N. Impact of non-homologous end-joining deficiency on random and targeted DNA integration: Implications for gene targeting. Nucleic Acids Res. 2008, 36, 6333–6342. [Google Scholar] [CrossRef]
- Zhang, G.; Guan, Q.; Chen, G.; Qian, F.; Liang, J. DNA methylation of the cdc2l1 gene promoter region decreases the expression of the cdk11p58 protein and reduces apoptosis in keloid fibroblasts. Arch. Dermatol. Res. 2018, 310, 107–115. [Google Scholar] [CrossRef]
- Capecchi, M.R. Gene targeting in mice: Functional analysis of the mammalian genome for the twenty-first century. Nat. Rev. Genet. 2005, 6, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Barrangou, R.; Doudna, J.A. Applications of crispr technologies in research and beyond. Nat. Biotechnol. 2016, 34, 933–941. [Google Scholar] [CrossRef] [PubMed]
- Whitworth, K.M.; Rowland, R.R.; Ewen, C.L.; Trible, B.R.; Kerrigan, M.A.; Cino-Ozuna, A.G.; Samuel, M.S.; Lightner, J.E.; McLaren, D.G.; Mileham, A.J.; et al. Gene-edited pigs are protected from porcine reproductive and respiratory syndrome virus. Nat. Biotechnol. 2016, 34, 20–22. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Lin, J.; Huang, J.; Zhang, H.; Zhang, R.; Zhang, X.; Cao, C.; Hambly, C.; Qin, G.; Yao, J.; et al. Reconstitution of ucp1 using crispr/cas9 in the white adipose tissue of pigs decreases fat deposition and improves thermogenic capacity. Proc. Natl. Acad. Sci. USA 2017, 114, E9474–E9482. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | SCNT Embryos (n) | Embryos Transferred (n) | Surrogates (n) | Pregnancies (n) | The Rate of Pregnancy (%) | The Number of Birth (n) | The Male ( ) Ratio (%) ) Ratio (%) | The Female ( ) Ratio (%) ) Ratio (%) | The Rate of Cloning Efficiency (%) |
|---|---|---|---|---|---|---|---|---|---|
| WT | 360 | 360 | 56 | 8 | 14.2 | 5 | 60.0 | 40.0 | 1.39 |
| Tβ4-OE | 1155 | 1137 | 241 | 19 | 8.9 | 15 | 46.7 | 53.3 | 1.32 |
| Type | Pathway Name | Count | p-Value | FDR 3 |
|---|---|---|---|---|
| DEG 1 | Arachidonic acid metabolism | 10 | 1.75846 × 10−5 | 0.002971802 |
| Linoleic acid metabolism | 7 | 1.17096 × 10−5 | 0.002971802 | |
| Nicotine addiction | 6 | 0.00036797 | 0.024874782 | |
| Steroid hormone biosynthesis | 8 | 0.000307903 | 0.024874782 | |
| Taste transduction | 7 | 0.000363475 | 0.024874782 | |
| Natural killer cell mediated cytotoxicity | 12 | 0.000649358 | 0.034851257 | |
| Neuroactive ligand-receptor interaction | 15 | 0.000721772 | 0.034851257 | |
| Retinol metabolism | 8 | 0.001021739 | 0.043168488 | |
| EIP 2 | Ribosome | 11 | 3.28496 × 10−8 | 3.57362 × 10−5 |
| Biosynthesis of amino acids | 7 | 1.12507 × 10−5 | 0.012238664 | |
| Carbon metabolism | 8 | 1.23556 × 10−5 | 0.013440547 | |
| Glycolysis/Gluconeogenesis | 6 | 8.69038 × 10−5 | 0.094499892 | |
| Biosynthesis of antibiotics | 9 | 0.000108387 | 0.117847957 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, B.; Liang, H.; Guo, D.-d.; Bi, Z.-w.; Yuan, J.-l.; Jin, Y.; Huan, L.; Guo, X.-d.; Cang, M.; Liu, D.-j. The Overexpression of Tβ4 in the Hair Follicle Tissue of Alpas Cashmere Goats Increases Cashmere Yield and Promotes Hair Follicle Development. Animals 2020, 10, 75. https://doi.org/10.3390/ani10010075
Dai B, Liang H, Guo D-d, Bi Z-w, Yuan J-l, Jin Y, Huan L, Guo X-d, Cang M, Liu D-j. The Overexpression of Tβ4 in the Hair Follicle Tissue of Alpas Cashmere Goats Increases Cashmere Yield and Promotes Hair Follicle Development. Animals. 2020; 10(1):75. https://doi.org/10.3390/ani10010075
Chicago/Turabian StyleDai, Bai, Hao Liang, Dong-dong Guo, Zhao-wei Bi, Jian-long Yuan, Yong Jin, Lei Huan, Xu-dong Guo, Ming Cang, and Dong-jun Liu. 2020. "The Overexpression of Tβ4 in the Hair Follicle Tissue of Alpas Cashmere Goats Increases Cashmere Yield and Promotes Hair Follicle Development" Animals 10, no. 1: 75. https://doi.org/10.3390/ani10010075
APA StyleDai, B., Liang, H., Guo, D.-d., Bi, Z.-w., Yuan, J.-l., Jin, Y., Huan, L., Guo, X.-d., Cang, M., & Liu, D.-j. (2020). The Overexpression of Tβ4 in the Hair Follicle Tissue of Alpas Cashmere Goats Increases Cashmere Yield and Promotes Hair Follicle Development. Animals, 10(1), 75. https://doi.org/10.3390/ani10010075