Morphometric Evaluation of Spermatogenic Cells and Seminiferous Tubules and Exploration of Luteinizing Hormone Beta Polypeptide in Testis of Datong Yak

 and
and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Exploration through Histology
2.3. Extraction of RNA and Synthesis of cDNA
2.4. PCR Amplification and Primer Design
2.5. Cloning
2.6. Analysis of LHB Using Quantitative Real-Time PCR
2.7. Analysis of LHB Using Semi-Quantitative PCR
2.8. Bioinformatics Analysis
2.9. Western Blot Analysis of LHB Protein
2.10. Statistical Analyses
3. Results
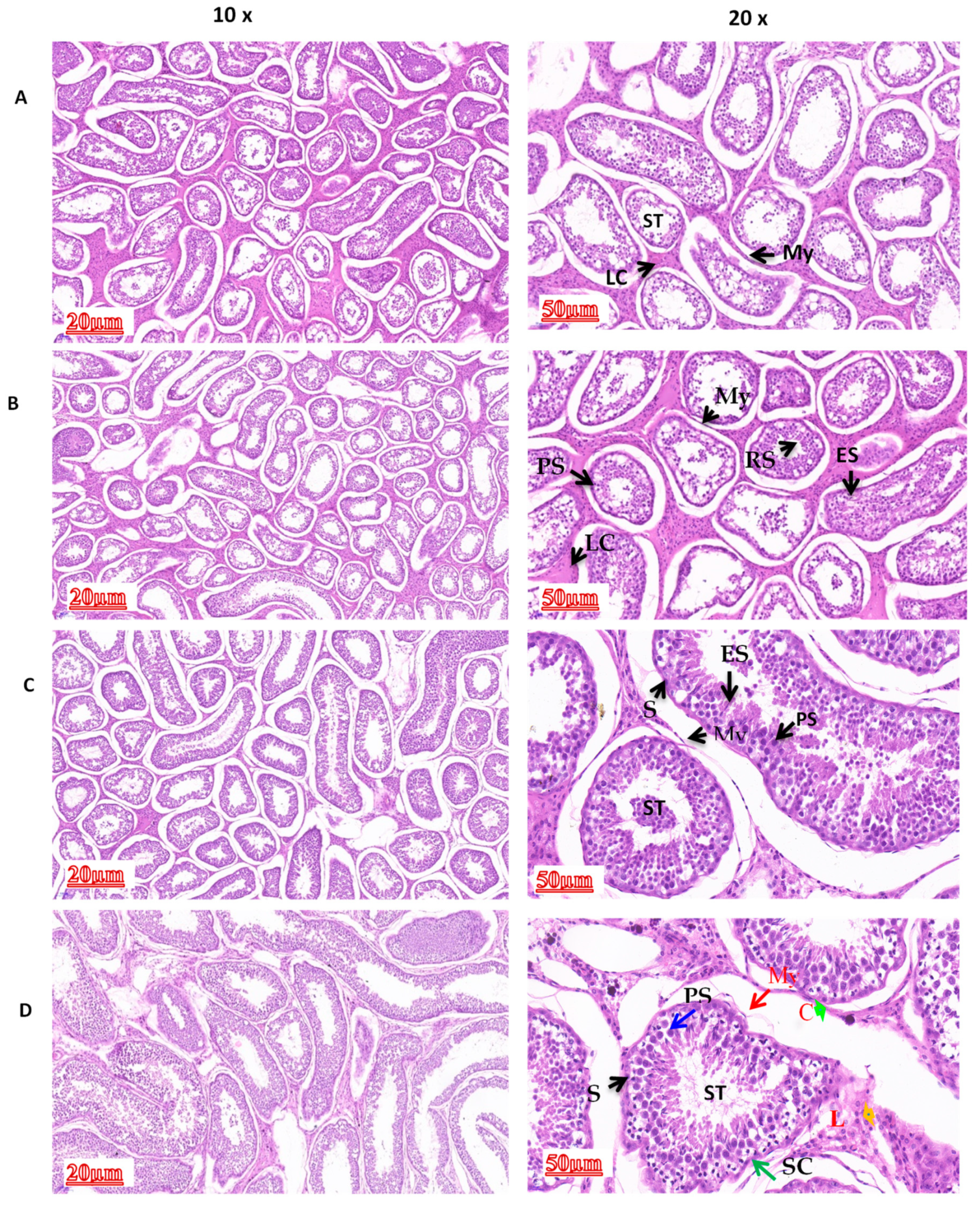
3.1. Comparison of Morphological Differences in Different Yak Developmental Stages
3.2. Diameters of Spermatogenic Cells and Their Nuclei in Yak Testis
3.3. Diameter of the Seminiferous Tubule and Number of Cells in the Testicles
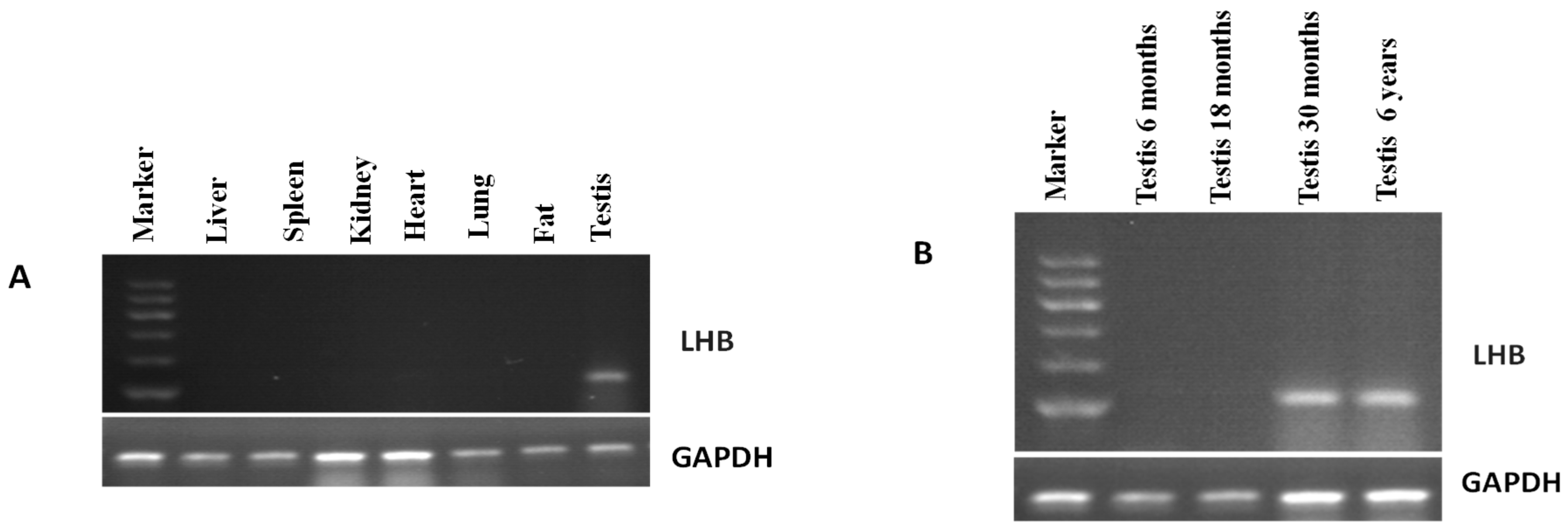
3.4. Expression Pattern of LHB Determined Using Semi-Quantitative PCR
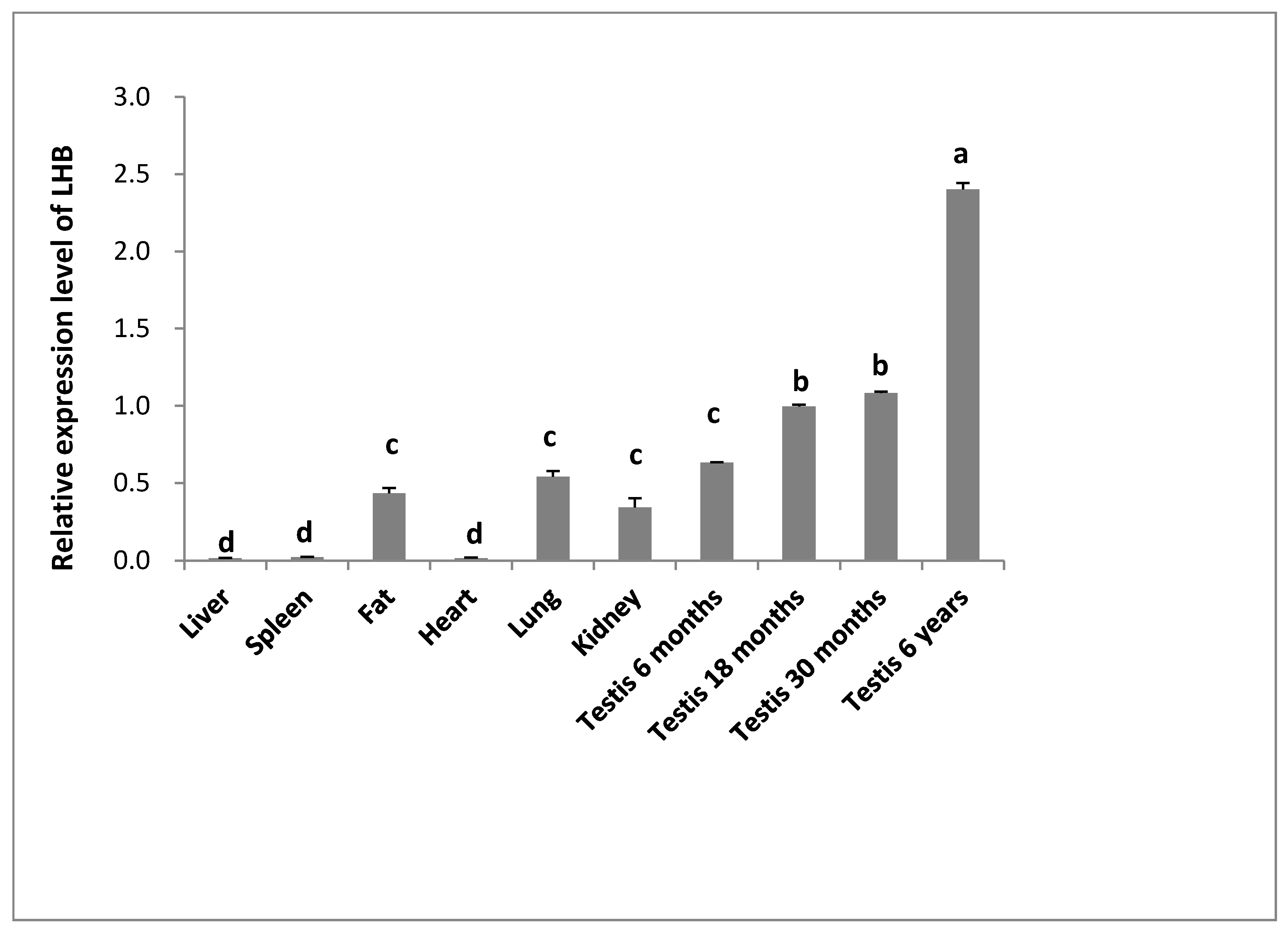
3.5. Expression Pattern of LHB Determined Using Quantitative Real-Time PCR
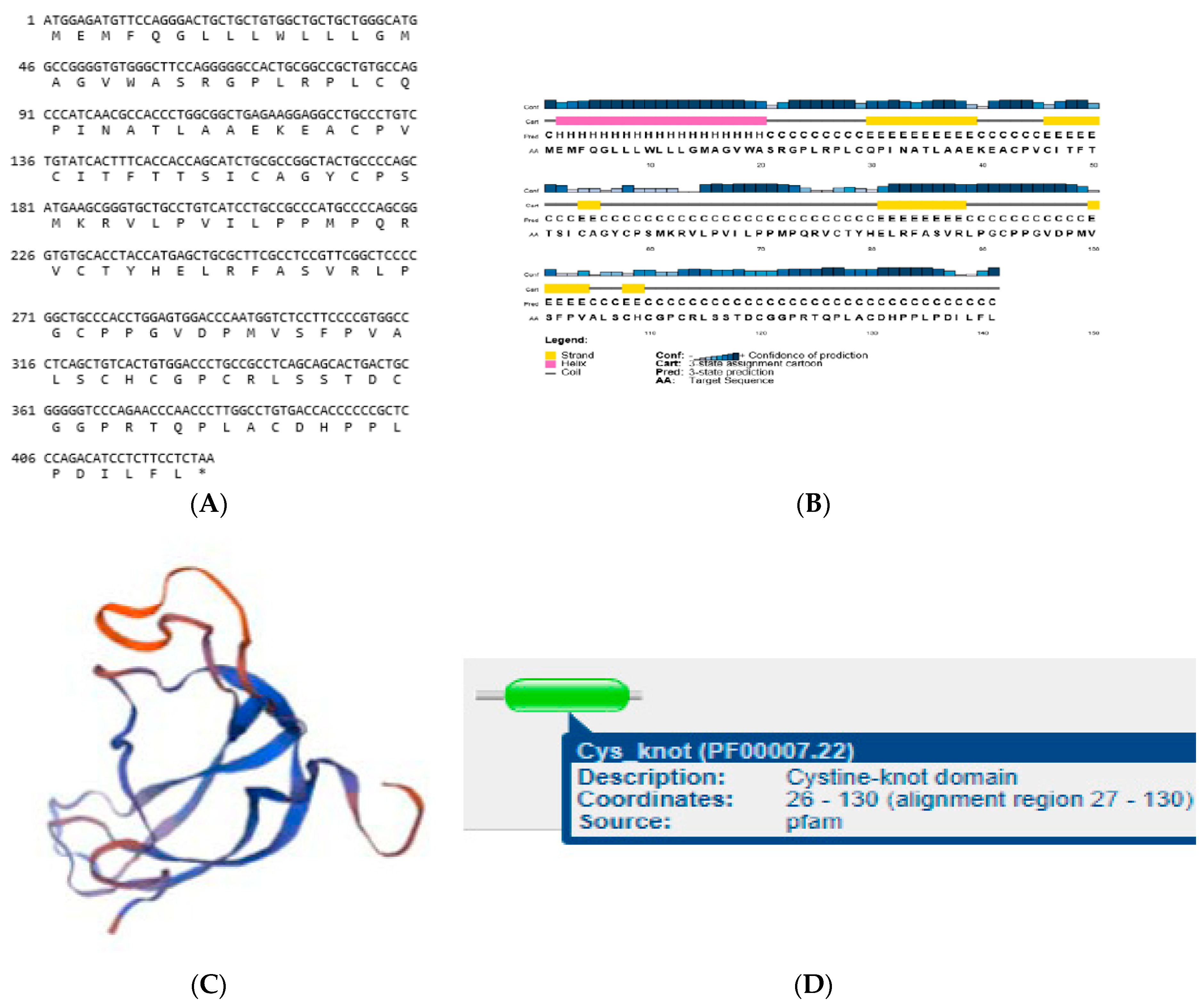
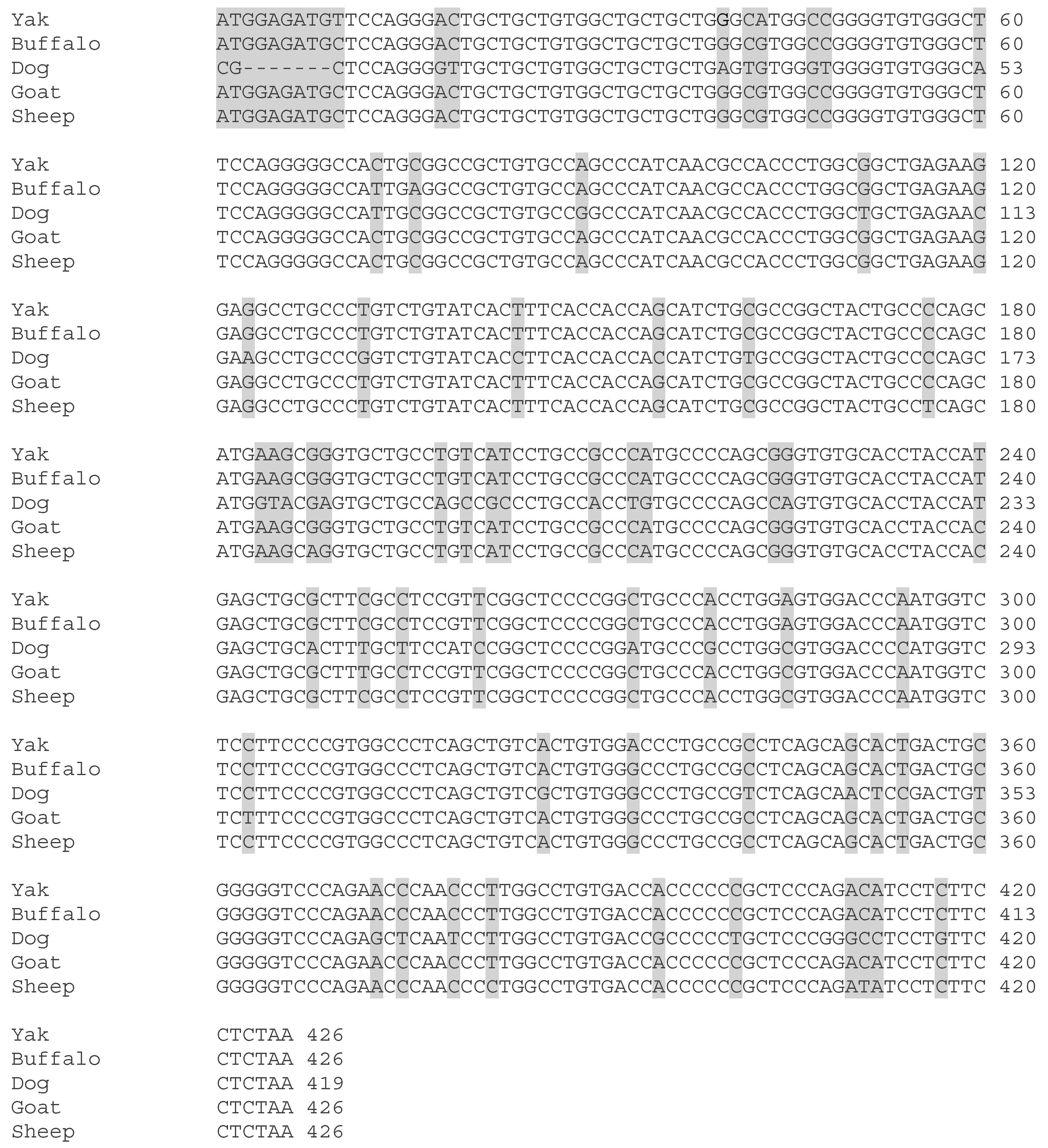
3.6. Structures of LHB
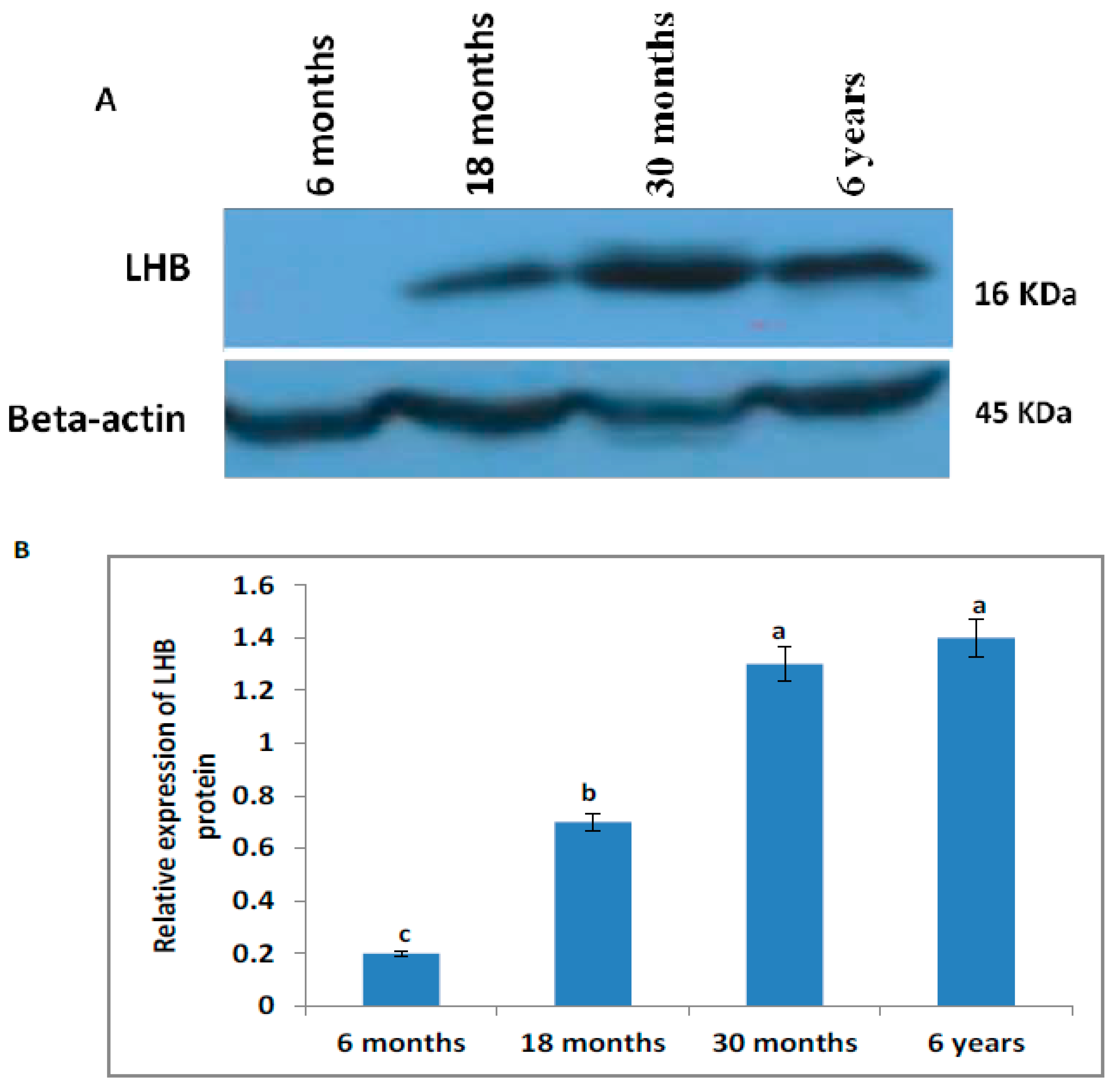
3.7. Western Blot Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Davis, J.R.; Langford, G.A.; Kirby, P.J. The testicular capsule. In The Testis. Development, Anatomy and Physiology; Johnson, A.D., Gomes, R., Vandemark, N.L., Eds.; Academic Press: London, UK, 1970; pp. 281–337. [Google Scholar]
- Hargrove, J.L.; MacIndoe, J.H.; Ellis, L.C. Testicular contractile cells and sperm transport. Fertil. Steril. 1977, 28, 1146–1157. [Google Scholar] [CrossRef]
- Banks, F.C.; Knight, G.E.; Calvert, R.C.; Turmaine, M.; Thompson, C.S.; Mikhailidis, D.P.; Burnstock, G. Smooth muscle and purinergic contraction of the human, rabbit, rat, and mouse testicular capsule. Biol. Reprod. 2006, 74, 473–480. [Google Scholar] [CrossRef]
- Maekawa, M.; Kamimura, K.; Nagano, T. Peritubular myoid cells in the testis: Their structure and function. Arch. Histol. Cytol. 1996, 59, 1–13. [Google Scholar] [CrossRef]
- Leal, M.C.; Becker-Silva, S.C.; Chiarini-Garcia, H.; Franca, L.R. Sertoli cell efficiency and daily sperm production in goats (Capra hircus). Anim. Reprod. 2004, 1, 122128. [Google Scholar]
- Almeida, F.F.L.; Marcelo, C.L.; Franca, L.R. Testis morphometry, duration of spermatogenesis and spermatogenic efficiency in the wild boar (Sus scrofa scrofa). Biol. Reprod. 2006, 75, 792–799. [Google Scholar] [CrossRef]
- Potorac, I.; Rivero-Müller, A.; Trehan, A.; Kiełbus, M.; Jozwiak, K.; Pralong, F.; Hafidi, A.; Thiry, A.; Ménagé, J.J.; Huhtaniemi, I.; et al. A vital region for human glycoprotein hormone trafficking revealed by an LHB mutation. J. Endocrinal. 2016, 231, 197–207. [Google Scholar] [CrossRef]
- Sairam, M.R.; Li, C.H. Human pituitary lutropin. Isolation, properties, and the complete amino acid sequence of the beta-subunit. Biochim. Biophys. Acta 1975, 412, 70–81. [Google Scholar] [CrossRef]
- Valdes-Socin, H.; Daly, A.F.; Beckers, A. Luteinizing Hormone Deficiency: Historical Views and Future Perspectives. Austin Androl. 2017, 2, 10–15. [Google Scholar]
- Lee, S.; Kim, N.K.; Kim, H.J.; Lee, S.H.; Jeong, H.J.; Cha, K.Y. Genetic analysis of three polymorphic sites of the luteinizing hormone beta-subunit gene in infertile Korean men with non-obstructive azoospermia. Fertil. Steril. 2003, 79, 517–521. [Google Scholar] [CrossRef]
- Lejeune, H.; Hebert, R.; Saez, J.M. Origin, proliferation, and differentiation of Leydig cells. J. Mol. Endocrinol. 1998, 20, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Berthezéne, F.; Forest, M.G.; Grimaud, J.A.; Claustrat, B.; Mornex, R. Leydig-cell agenesis: A cause of male pseudohermaphroditism. N. Engl. J. Med. 1976, 295, 969–972. [Google Scholar] [CrossRef]
- Yang, X.; Ochin, H.; Shu, L.; Liu, J.; Shen, J.; Liu, J.; Lin, C.; Cui, Y. Homozygous nonsense mutation Trp28X in the LHB gene causes male hypogonadism. J. Assist. Reprod. Genet. 2018, 35, 913–919. [Google Scholar] [CrossRef]
- Manna, P.R.; Joshi, L.; Reinhold, V.N.; Aubert, M.L.; Suganuma, N.; Pettersson, K.; Huhtaniemi, I.T. Synthesis of purification and structural and functional characterization of a recombinant form of a common genetic variant of the human luteinizing hormone. Hum. Mol. Genet. 2002, 11, 301–315. [Google Scholar] [CrossRef]
- Themmen, A.P.N.; Huhtaniemi, I.T. Mutations of gonadotropins and gonadotropin receptors: Elucidating the physiology and pathophysiology of pituitary-gonadal function. Endocr. Rev. 2000, 21, 551–583. [Google Scholar] [CrossRef] [PubMed]
- Achermann, J.C.; Jameson, J.L. Fertility and infertility: Genetic contributions from the hypothalamic-pituitary-gonadal axis. Mol. Endocrinol. 1999, 13, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Wiener, G.; Jianlin, H.; Ruijun, L. The Yak, 2nd ed.; FAO Regional Office for Asia and the Pacific Food and Agriculture Organization of the United Nations: Bangkok, Thailand, 2003; pp. 88–102. [Google Scholar]
- Ma, X.; Dong, Y.; Matzuk, M.M.; Kumar, T.R. Targeted disruption of luteinizing hormone beta-subunit leads to hypogonadism, defects in gonadal steroidogenesis, and infertility. Proc. Natl. Acad. Sci. USA 2004, 101, 17294–17299. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Wang, X.; Zhang, H.; Chen, Z.; Zhao, X.; Ma, Y. Histomorphological Comparisons and Expression Patterns of BOLL Gene in Sheep Testes at Different Development Stages. Animals 2019, 9, 105. [Google Scholar] [CrossRef]
- Qudratullah, K.; Xuezhi, D.; Anum, A.A.; Min, C.; Xiaoyun, W.; Pengjia, B.; Ping, Y. Expression Analysis of IZUMO1 Gene during Testicular Development of Datong Yak (Bos Grunniens). Animals 2019, 9, 292. [Google Scholar]
- Niu, W.; Qazi, I.H.; Li, S.; Zhao, X.; Yin, H.; Wang, Y.; Zhu, Q.; Han, H.; Zhou, G.; Du, X. Expression of FOXL2 and RSPO1 in Hen Ovarian Follicles and Implication of Exogenous Leptin in Modulating Their mRNA Expression in In Vitro Cultured Granulosa Cells. Animals 2019, 9, 1083. [Google Scholar] [CrossRef]
- Hou, Y.; Zhou, X.; Liu, J.; Yuan, J.; Cheng, H.; Zhou, R. Nuclear factor-Y (NF-Y) regulates transcription of mouse Dmrt7 gene by binding to tandem CCAAT boxes in its proximal promoter. Int. J. Biol. Sci. 2010, 6, 655. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C (T) method. Nat. Protoc. 2008, 3, 1101. [Google Scholar] [CrossRef] [PubMed]
- Matzuk, M.M.; Lamb, D.J. Genetic dissection of mammalian fertility pathways. Nat. Cell Biol. 2002, 4, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Weinbauer, G.F.; Nieschlag, E. Hormonal control of spermatogenesis. In Molecular Biology of the Male Reproductive System; De Kretser, D.M., Ed.; Academic Press: San Diego, CA, USA, 1993; pp. 99–142. [Google Scholar]
- O’Donnell, L.; Robertson, K.M.; Jones, M.E.; Simpson, E.R. Estrogen and Spermatogenesis. Endocr. Rev. 2001, 22, 289–318. [Google Scholar] [CrossRef] [PubMed]
- Sherman, G.B.; Wolfe, M.W.; Farmerie, T.A.; Clay, C.M.; Threadgill, D.S.; Sharp, D.C.; Nilson, J.H. A single gene encodes the beta-subunits of equine luteinizing hormone and chorionicgonadotropin. Mol. Endocrinol. 1992, 6, 9519. [Google Scholar]
- Talmadge, K.; Vamvakopoulos, N.C.; Fiddes, J.C. Evolution of the genes for the beta subunits of human chorionic gonadotropin and luteinizing hormone. Nature 1984, 307, 37–40. [Google Scholar] [CrossRef]
- Valdes-Socin, H.; Salvi, R.; Thiry, A.; Daly, A.F.; Pralong, F.P.; Gaillard, R.; Beckers, A. Testicular effects of isolated luteinizing hormone deficiency and reversal by long-term human chorionic gonadotropin treatment. J. Clin. Endocr. Metab. 2009, 94, 3–4. [Google Scholar] [CrossRef]
- Valdes-Socin, H.; Salvi, R.; Daly, A.F.; Gaillard, R.C.; Quatresooz, P.; Tebeu, P.M.; Pralong, F.P.; Beckers, A. Hypogonadism in a patient with a mutation in the luteinizing hormone beta-subunit gene. N. Engl. J. Med. 2004, 351, 2619–2625. [Google Scholar] [CrossRef]
- Weiss, J.; Axelrod, L.; Whitcomb, R.W.; Harris, P.E.; Crowley, W.F.; Jameson, J.L. Hypogonadism caused by a single amino acid substitution in the beta subunit of luteinizing hormone. N. Engl. J. Med. 1992, 326, 179–183. [Google Scholar] [CrossRef]
- Valdes-Socin, H.; Beckers, A. Pasqualini’s syndrome: Hypoandrogenism with spermatogenesis. Medicina 2015, 75, 53–58. [Google Scholar]
- Cahoreau, C.; Klett, D.; Combarnous, Y. Structure-function relationships of glycoprotein hormones and their subunits’ ancestors. Front. Endocrinol. 2015, 6, 26. [Google Scholar] [CrossRef]
- Lapthorn, A.J.; Harris, D.C.; Littlejohn, A.; Lustbader, J.W.; Canfield, R.E.; Machin, K.J.; Morgan, F.J.; Isaacs, N.W. Crystal structure of human chorionic gonadotropin. Nature 1994, 369, 455–461. [Google Scholar] [CrossRef]
- Xing, Y.; Lin, W.; Jiang, M.; Myers, R.V.; Cao, D.; Bernard, M.P.; Moyle, W.R. Alternatively Folded Chorionic gonadotropin Analogs implications for hormone folding and biological activity. J. Biol. Chem. 2001, 276, 46953–46960. [Google Scholar] [CrossRef] [PubMed]
- Huhtaniemi, I.T.; Themmen, A.P. Mutations in human gonadotropin and gonadotropin receptor genes. Endocrine 2005, 26, 207–217. [Google Scholar] [CrossRef]
- Kamal, S.; Devi, J. Morphometrical changes of the seminiferous tubules and Leydig cells in Assam goats (Caprahircus) from birth to 10 months. J. Appl. Anim. Res. 2017, 45, 268–274. [Google Scholar]
- Wrobel, K.H.; Dostal, S.; Schimmel, M. Postnatal development of the tubular lamina propria and the intertubular tissue in the bovine testis. Cell Tissue Res. 1988, 252, 639–653. [Google Scholar] [CrossRef]
- Baishya, G.; Ahmed, S.; Bhattacharya, M. A correlative study on biometry and histomorphometry of Male gonad and thyroid gland (0–90 days) in Assam goat (Capra hircus). Indian Vet. J. 1986, 63, 928–932. [Google Scholar]
- Ahmad, N.; Umair, S.; Shahab, M.; Arslan, M. Testicular development and establishment of spermatogenesis in Nili-Ravi buffalo bulls. Theriogenology 2010, 73, 20–25. [Google Scholar] [CrossRef]
- Akosman, M.S.; Lenger, O.F.; Demirel, H.H. Morphological, stereological and histometrical assessment of the testicular parameters between Holstein and Simmental bulls. Int. J. Morphol. 2013, 31, 1076–1080. [Google Scholar] [CrossRef]
- Mohammed, A.H.S.; Kadium, D.A.H.; Ebed, A.K. Some morphometric and histological description of the seminiferous, striaghted and rete testis tubules in the testis of indigenous male goats (two years old). Kufa J. Vet. Med Sci. 2011, 2, 19–29. [Google Scholar]
- Ibrahim, N.S.; Al-sahaf, M.M.H.; Alwan, A.F. Reproductive activity of mature Iraqi bull buffaloes: Testes dimensions and histological picture. Int. J. Anim. Vet. Adv. 2013, 5, 34–37. [Google Scholar] [CrossRef]
- Nishimura, S.; Okano, K.; Yasukouchi, K.; Gotoh, T.; Tabata, S.; Iwamoto, H. Testis developments and puberty in the male Tokara (Japanese native) goat. Anim. Reprod. Sci. 2000, 64, 127–131. [Google Scholar] [CrossRef]
- Franca, L.R.; Russell, L.D. The testis of domestic animals in: Male reproduction. In A Multidisciplinary Overview; Regadera, J., Martinez-Garcia, F., Eds.; Churchill Livingstone: Madrid, Spain, 1998; pp. 197–219. [Google Scholar]
- Mustafa, S.; Wei, Q.; Ennab, W.; Lv, Z.; Nazar, K.; Siyal, F.A.; Rodeni, S.; Kavita, N.M.X.; Shi, F. Resveratrol Ameliorates Testicular Histopathology of Mice Exposed to Restraint Stress. Animals 2019, 9, 743. [Google Scholar] [CrossRef] [PubMed]
- Paulo, A.T.A.; Deiler, S.C.; Fábio, J.C.F.; Carlos, A.C.F.; Marcelo, D.S.; Juliana, C.B.S. Testicular Histomorphometric Evaluation of Zebu Bull Breeds. Braz. Arch. Biol. Technol. 2014, 57, 900–907. [Google Scholar]
- Karmore, S.K.; Bhamburkar, V.R.; Banubakode, B.; Dalvi, R.S.; Waghaye, Y. Age wise changes in the histological and histochemical status of the interstitial cells in goat (Capra hircus). Indian J. Vet. Anat. 2001, 13, 49–52. [Google Scholar]
- Sun, L.; Mipam, T.D.; Zhao, F.; Liu, W. Comparative testis proteome of cattleyak from different developmental stages. Animal 2017, 11, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Yan, P.; Xiang, L.; Guo, X.; Bao, P.J.; Jin, S.; Wu, X.Y. The low expression of Dmrt7 is associated with spermatogenic arrest in cattle-yak. Mol. Biol. Rep. 2014, 41, 7255–7263. [Google Scholar] [CrossRef]
- Achard, C.; Courtillot, C.; Lahuna, O.; Méduri, G.; Soufir, J.C.; Lière, P.; Bachelot, A.; Benyounes, H.; Schumacher, M.; Kuttenn, F.; et al. Normal Spermatogenesis in a Man with Mutant Luteinizing Hormone. N. Engl. J. Med. 2009, 361, 1856–1863. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession No | Gene | Primer Sequence (5’- >3’) | Product Length (bp) | Annealing Temperature (°C) |
|---|---|---|---|---|
| XM_005895365.2 | LHB (For cloning) | F: TGGAGTGGACCCAATGGTCT | 426 | 60.48 |
| R:TAGGATGGGCATGGGAGGTT | 60.33 | |||
| XM_019980229.1 | LHB (For gene expression) | F:ACCAGTGCAGTGAGACGAAG | 131 | 59.97 |
| R:GATTAAAGCCTGGGAAGGGC | 58.60 | |||
| NM_001034034.2 | GAPDH | F: ATGAAAGGGCCATCACCATC | 204 | 55.85 |
| R: GTGGTTCACGCCCATCACA | 60.00 | |||
| L08165.1 | β-actin | F:TGGGTATGGAGTCCTGTGGT | 160 | 60.00 |
| R: AGGGCTGTGATCTCCTTCTG | 60.40 |
| Germ Cells | 6 Months | 18 Months | 30 Months | 6 Years |
|---|---|---|---|---|
| Spermatogonium | 4.23 ± 0.56 a | 4.60 ± 0.38 a | 5.90 ± 0.70 b | 6.52 ± 0.79 b |
| Spermatogonium nuclei | 2.30 ± 1.20 a | 2.70 ± 1.80 a | 3.85 ± 1.20 b | 4.80 ± 1.80 b |
| Primary spermatocyte | 4.55 ± 0.41a | 5.13 ± 0.50 a | 6.20 ± 0.90 b | 7.10 ± 0.89 b |
| Primary spermatocyte nuclei | 2.10 ± 0.98 a | 2.63 ± 1.76 a | 3.70 ± 0.98 b | 5.63 ± 1.76 b |
| Round spermatid | 4.90 ± 1.10 a | 5.22 ± 2.43 a | 6.10 ± 1.10 b | 8.22 ± 2.43 b |
| Round spermatid nuclei | 4.01 ± 1.21 a | 6.89 ± 1.72 ab | 7.01 ± 1.21 b | 6.89 ± 1.72 b |
| Sertoli cells | 2.30 ± 0.76 a | 3.17 ± 0.52 b | 4.70 ± 0.49 c | 5.20 ± 0.96 c |
| Leydig cells | 4.10 ± 0.66 a | 5.10 ± 1.76 b | 6.20 ± 1.65 c | 8.30 ± 1.52 d |
| Germ Cells | 6 Months | 18 Months | 30 Months | 6 Years |
|---|---|---|---|---|
| Tubular diameter (μm) | 190.0 ± 0.41 a | 220.82 ± 0.67 b | 235.69 ± 0.50 c | 245.69 ± 0.59 d |
| Epithelial height (μm) | 45.70 ± 0.45 a | 60.33 ± 0.75 b | 61.99 ± 0.67 b | 72.99 ± 0.52 c |
| Luminal diameter (µm2) | 50.23 ± 0.67 a | 60.94 ± 0.92 b | 72.84 ± 0.66 c | 88.94 ± 0.82 d |
| Luminal area (µm2) | 37.40 ± 9.38 a | 49.26 ± 8.47 b | 56.29 ± 6.70 c | 68.33 ± 8.244 d |
| Leydig cell area (µm2) | 90.19 ± 10.60 a | 114.73 ± 20.6 b | 126.99 ± 26.10 c | 149.76 ± 34.28 d |
| Width of tunica Albuginea (µm) | 12.09 ± 0.20 a | 13.05 ± 1.09 a | 14.30 ± 1.1 a | 15.41 ± 0.80 a |
| Cross sectional area (µm2) | 46,675.0 ± 2689 a | 47,695.0 ± 2910 a | 48,200 ± 3025 a | 50,812 ± 3711 a |
| ST Volume density (%) | 68.21 ± 0.15 a | 70.84 ± 0.73 a | 75.25 ± 0.50 a | 78.84 ± 0.73 b |
| Leydig cells (%) | 8.00 ± 3.56 a | 10.00 ± 1.76 b | 26.00 ± 2.65 a | 30.00 ± 1.52 a |
| Sertoli cells (%) | 16.20 ± 2.56 a | 20.0 ± 3.55 b | 26.00 ± 2.46 c | 41.00 ± 4.56 d |
| Spermatogonium (%) | 85.33 ± 17.60 a | 160 ± 25.49 b | 180 ± 15.80 c | 200 ± 18.79 d |
| Spermatocyte (%) | 9.00 ± 1.86 a | 14.60 ± 1.50 b | 17.25 ± 2.80 c | 20.50 ± 1.90 d |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalwar, Q.; Chu, M.; Ahmad, A.A.; Ding, X.; Wu, X.; Bao, P.; Yan, P. Morphometric Evaluation of Spermatogenic Cells and Seminiferous Tubules and Exploration of Luteinizing Hormone Beta Polypeptide in Testis of Datong Yak. Animals 2020, 10, 66. https://doi.org/10.3390/ani10010066
Kalwar Q, Chu M, Ahmad AA, Ding X, Wu X, Bao P, Yan P. Morphometric Evaluation of Spermatogenic Cells and Seminiferous Tubules and Exploration of Luteinizing Hormone Beta Polypeptide in Testis of Datong Yak. Animals. 2020; 10(1):66. https://doi.org/10.3390/ani10010066
Chicago/Turabian StyleKalwar, Qudratullah, Min Chu, Anum Ali Ahmad, Xuezhi Ding, Xiaoyun Wu, Pengjia Bao, and Ping Yan. 2020. "Morphometric Evaluation of Spermatogenic Cells and Seminiferous Tubules and Exploration of Luteinizing Hormone Beta Polypeptide in Testis of Datong Yak" Animals 10, no. 1: 66. https://doi.org/10.3390/ani10010066
APA StyleKalwar, Q., Chu, M., Ahmad, A. A., Ding, X., Wu, X., Bao, P., & Yan, P. (2020). Morphometric Evaluation of Spermatogenic Cells and Seminiferous Tubules and Exploration of Luteinizing Hormone Beta Polypeptide in Testis of Datong Yak. Animals, 10(1), 66. https://doi.org/10.3390/ani10010066