
The Terrestrial Plastisphere: Diversity and Polymer-Colonizing Potential of Plastic-Associated Microbial Communities in Soil

 , , , , and
, , , , and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Microbial Community Analysis
2.2.1. DNA Extraction
2.2.2. Paired End Illumina Sequencing and Bioinformatics
2.2.3. Statistics and Data Visualization
2.2.4. Thermogravimetric Analysis for Plastic Content Quantification
2.3. Colonization Experiment
2.3.1. Plastic Material and UV-Weathering
2.3.2. Medium and Preparation of Microbial Inoculum
2.3.3. Light Imaging and Field Emission Scanning Electron Microscopy of Microbial Colonization
3. Results
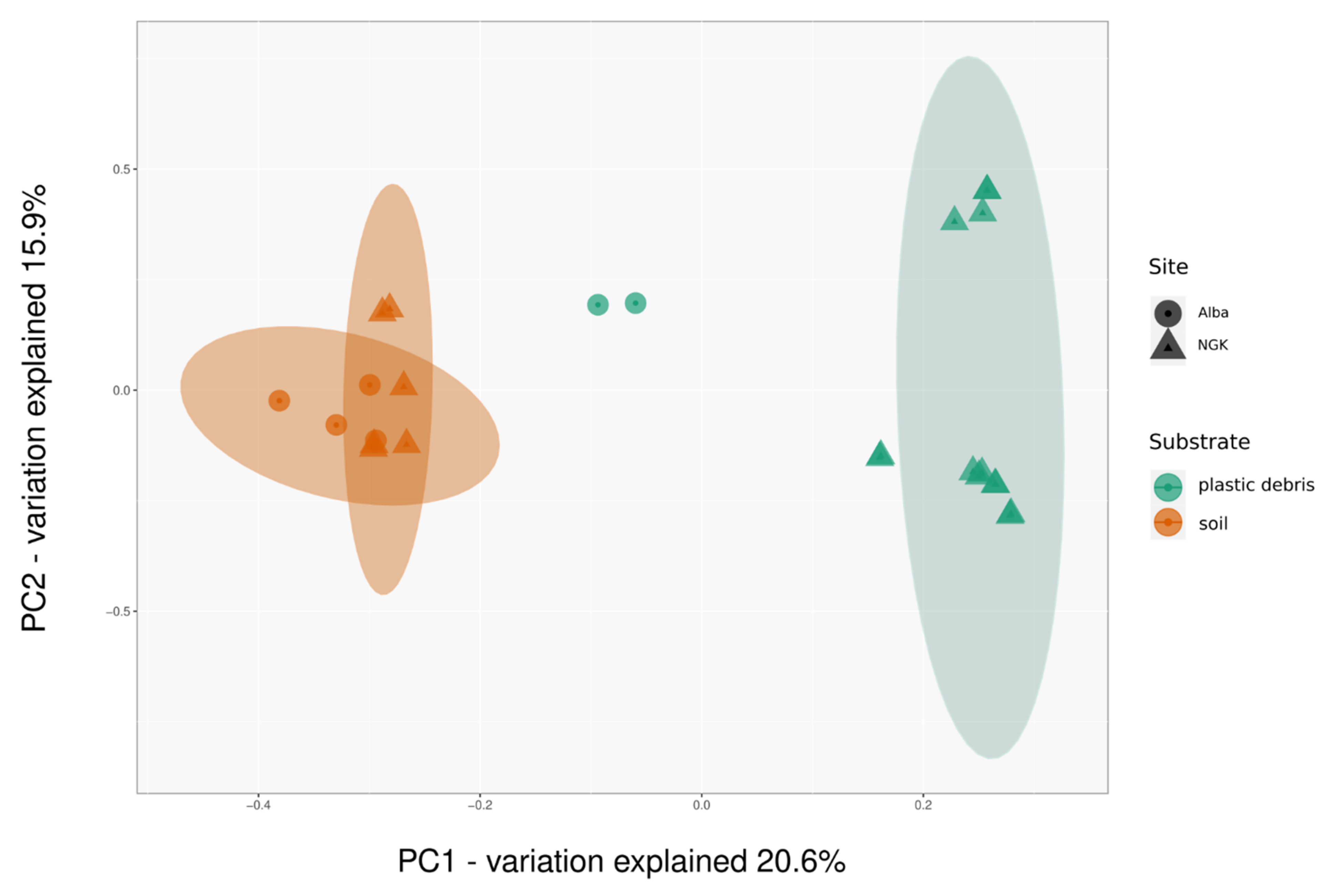
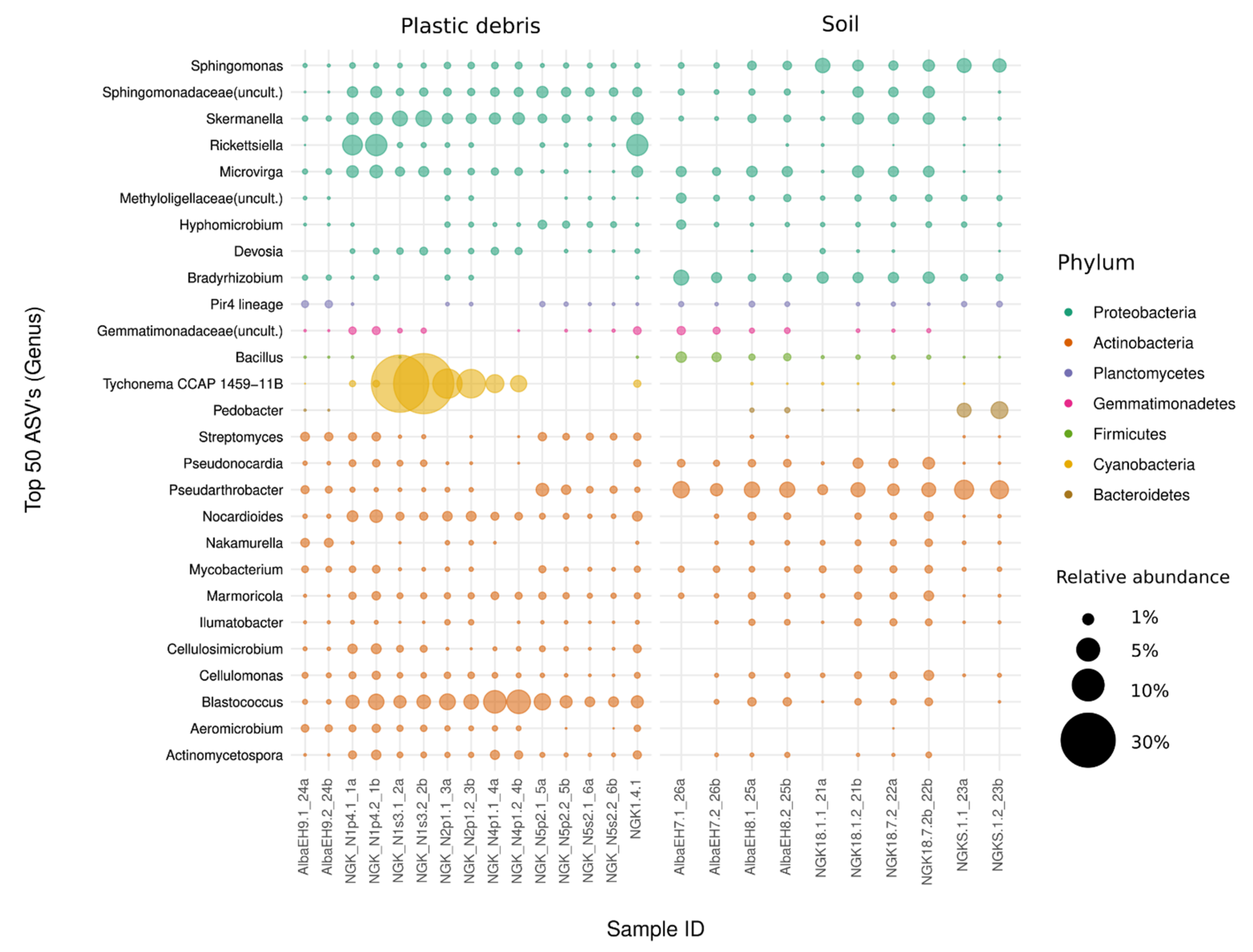
3.1. Diversity and Community Composition
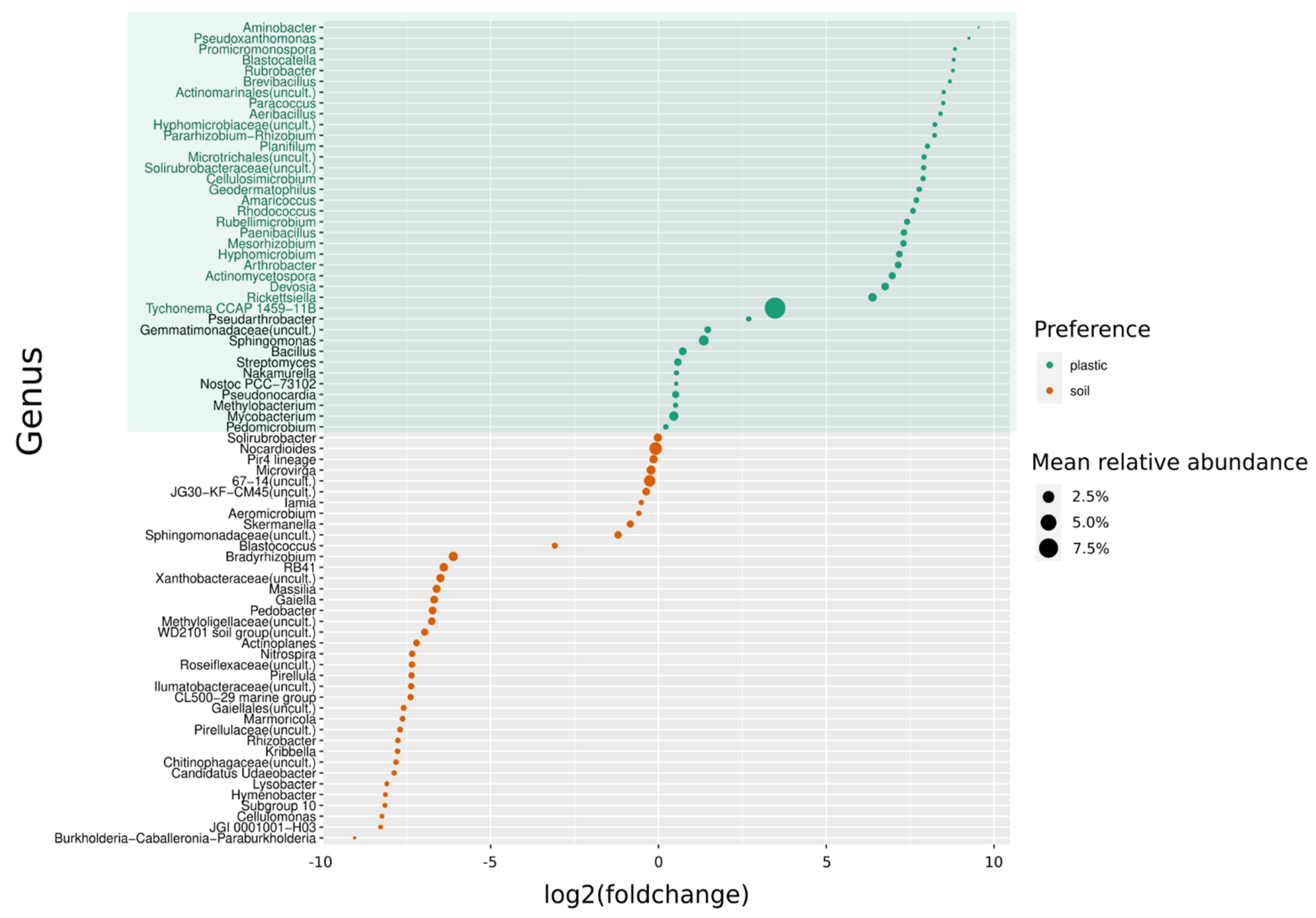
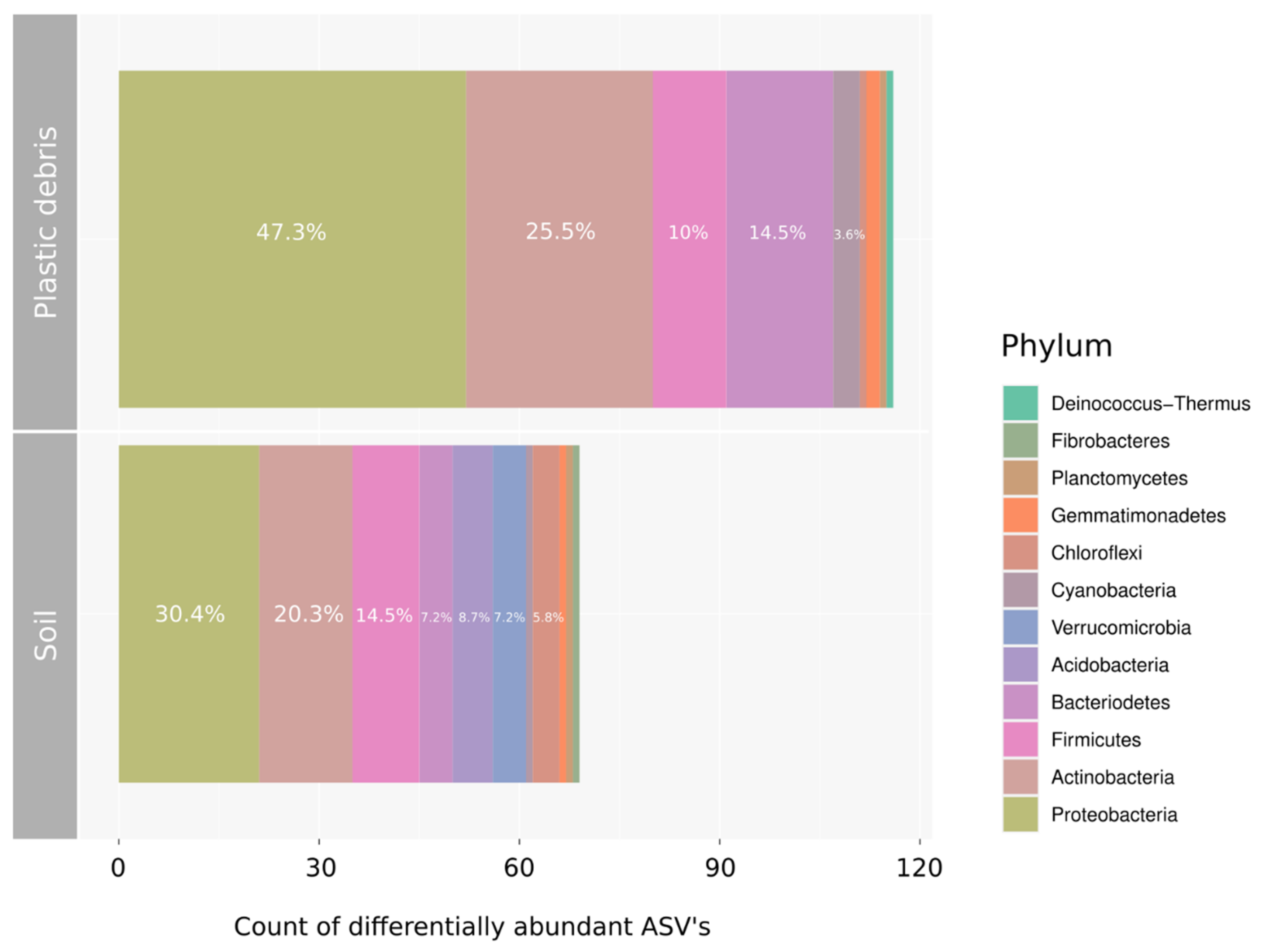
3.2. Community Assembly
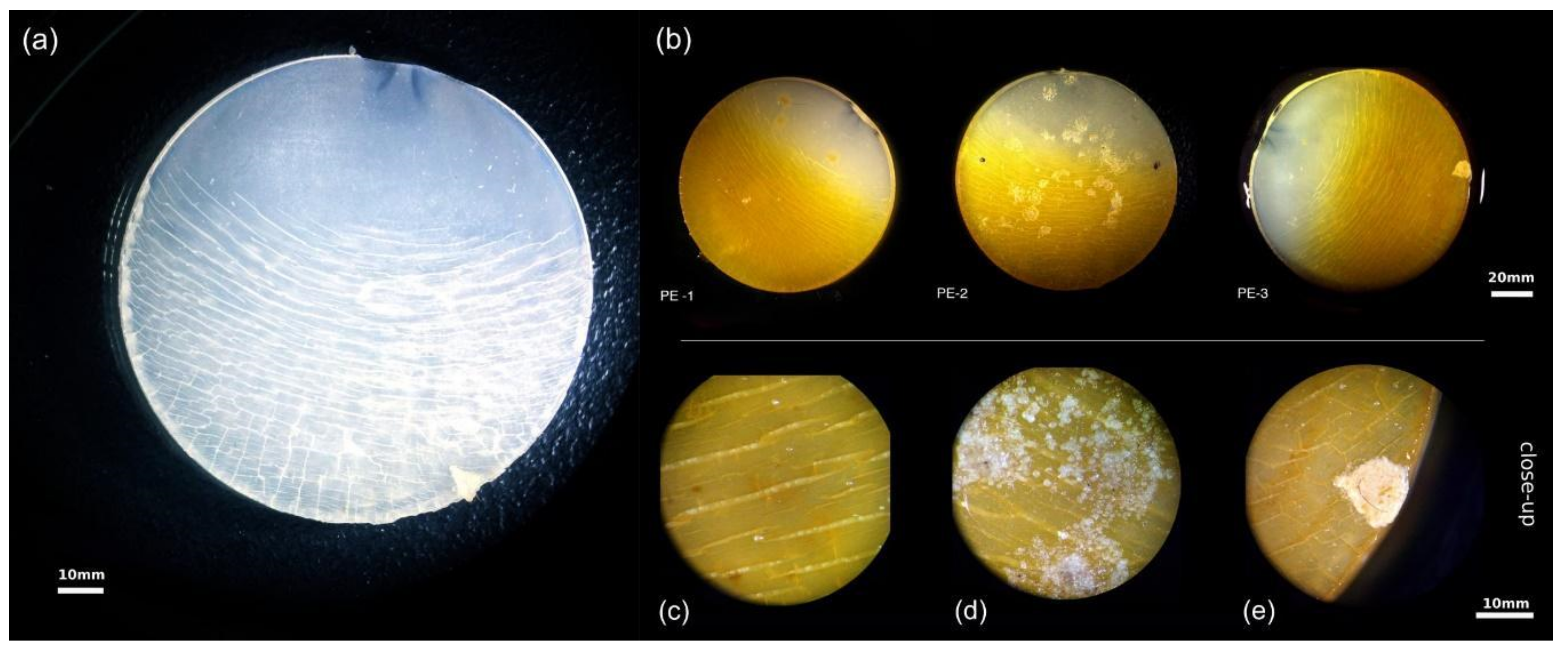
3.3. Surface Deterioration of Polyethylene Facilitated by UV-Weathering and Microbial Attachment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Machado, A.A.S.D.S.; Lau, C.W.; Till, J.; Kloas, W.; Lehmann, A.; Becker, R.; Rillig, M. Impacts of Microplastics on the Soil Biophysical Environment. Environ. Sci. Technol. 2018, 52, 9656–9665. [Google Scholar] [CrossRef] [Green Version]
- Haegerbaeumer, A.; Mueller, M.-T.; Fueser, H.; Traunspurger, W. Impacts of Micro- and Nano-Sized Plastic Particles on Benthic Invertebrates: A Literature Review and Gap Analysis. Front. Environ. Sci. 2019, 7. [Google Scholar] [CrossRef] [Green Version]
- Zettler, E.; Mincer, T.; Amaral-Zettler, L.A. Life in the “Plastisphere”: Microbial Communities on Plastic Marine Debris. Environ. Sci. Technol. 2013, 47, 7137–7146. [Google Scholar] [CrossRef]
- De Tender, C.; Devriese, L.I.; Haegeman, A.; Maes, S.; Ruttink, T.; Dawyndt, P. Bacterial Community Profiling of Plastic Litter in the Belgian Part of the North Sea. Environ. Sci. Technol. 2015, 49, 9629–9638. [Google Scholar] [CrossRef]
- McCormick, A.; Hoellein, T.J.; Mason, S.A.; Schluep, J.; Kelly, J. Microplastic is an Abundant and Distinct Microbial Habitat in an Urban River. Environ. Sci. Technol. 2014, 48, 11863–11871. [Google Scholar] [CrossRef]
- Amaral-Zettler, L.A.; Zettler, E.; Slikas, B.; Boyd, G.D.; Melvin, D.W.; Morrall, C.E.; Proskurowski, G.; Mincer, T.J. The biogeography of the Plastisphere: Implications for policy. Front. Ecol. Environ. 2015, 13, 541–546. [Google Scholar] [CrossRef]
- Horton, A.A.; Walton, A.; Spurgeon, D.J.; Lahive, E.; Svendsen, C. Microplastics in freshwater and terrestrial environments: Evaluating the current understanding to identify the knowledge gaps and future research priorities. Sci. Total Environ. 2017, 586, 127–141. [Google Scholar] [CrossRef] [Green Version]
- Machado, A.A.D.S.; Kloas, W.; Zarfl, C.; Hempel, S.; Rillig, M. Microplastics as an emerging threat to terrestrial ecosystems. Glob. Chang. Biol. 2018, 24, 1405–1416. [Google Scholar] [CrossRef] [Green Version]
- Browne, M.A.; Crump, P.; Niven, S.J.; Teuten, E.; Tonkin, A.; Galloway, T.; Thompson, R. Accumulation of Microplastic on Shorelines Woldwide: Sources and Sinks. Environ. Sci. Technol. 2011, 45, 9175–9179. [Google Scholar] [CrossRef]
- Rillig, M.C. Microplastic in Terrestrial Ecosystems and the Soil? Environ. Sci. Technol. 2012, 46, 6453–6454. [Google Scholar] [CrossRef]
- Rillig, M.C.; Lehmann, A. Microplastic in terrestrial ecosystems. Science 2020, 368, 1430–1431. [Google Scholar] [CrossRef] [PubMed]
- Labbate, M.; Seymour, J.R.; Lauro, F.; Brown, M.V. Anthropogenic Impacts on the Microbial Ecology and Function of Aquatic Environments. Front. Microbiol. 2016, 7, 1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.-F.; Wang, M.-X.; Zuo, X.-H.; Yao, Y.-H. Comparative Investigation of Bacterial, Fungal, and Archaeal Community Structures in Soils in a Typical Oilfield in Jianghan, China. Arch. Environ. Contam. Toxicol. 2016, 72, 65–77. [Google Scholar] [CrossRef]
- Sun, M.Y.; Dafforn, K.; Johnston, E.; Brown, M.V. Core sediment bacteria drive community response to anthropogenic contamination over multiple environmental gradients. Environ. Microbiol. 2013, 15, 2517–2531. [Google Scholar] [CrossRef]
- Zhang, M.; Zhao, Y.; Qin, X.; Jia, W.; Chai, L.; Huang, M.; Huang, Y. Microplastics from mulching film is a distinct habitat for bacteria in farmland soil. Sci. Total Environ. 2019, 688, 470–478. [Google Scholar] [CrossRef]
- Huang, Y.; Zhao, Y.; Wang, J.; Zhang, M.; Jia, W.; Qin, X. LDPE microplastic films alter microbial community composition and enzymatic activities in soil. Environ. Pollut. 2019, 254, 112983. [Google Scholar] [CrossRef]
- Puglisi, E.; Romaniello, F.; Galletti, S.; Boccaleri, E.; Frache, A.; Cocconcelli, P.S. Selective bacterial colonization processes on polyethylene waste samples in an abandoned landfill site. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Esan, E.; Abbey, L.; Yurgel, S. Exploring the long-term effect of plastic on compost microbiome. PLoS ONE 2019, 14, e0214376. [Google Scholar] [CrossRef] [Green Version]
- Rüthi, J.; Bölsterli, D.; Pardi-Comensoli, L.; Brunner, I.; Frey, B. The “Plastisphere” of biodegradable plastics is characterized by specific microbial taxa of Alpine and Arctic soils. Front. Environ. Sci. 2020, 8, 562263. [Google Scholar] [CrossRef]
- Bandopadhyay, S.; González, J.E.L.Y.; Henderson, K.B.; Anunciado, M.B.; Hayes, D.G.; Debruyn, J.M. Soil Microbial Communities Associated With Biodegradable Plastic Mulch Films. Front. Microbiol. 2020, 11, 587074. [Google Scholar] [CrossRef]
- Azeko, S.; Odusanya, O.; Malatesta, K.; Anuku, N.; Soboyejo, W. Bacterial Remediation of Polyethylene by Serratia marcescens sub sp. marcescens and its Supernatant. Adv. Mater. Res. 2015, 1132, 238–251. [Google Scholar] [CrossRef]
- Jumaah, O.S. Screening of Plastic Degrading Bacteria from Dumped Soil Area. IOSR J. Environ. Sci. Toxicol. Food Technol. 2017, 11, 93–98. [Google Scholar] [CrossRef]
- Odusanya, S.; Nkwogu, J.; Alu, N.; Udo, G.E.; Ajao, J.; Osinkolu, G.; Uzomah, A. Preliminary Studies on Microbial Degradation of Plastics Used in Packaging Potable Water in Nigeria. Niger. Food J. 2013, 31, 63–72. [Google Scholar] [CrossRef] [Green Version]
- Shao, H.; Chen, M.; Fei, X.; Zhang, R.; Zhong, Y.; Ni, W.; Tao, X.; He, X.; Zhang, E.; Yong, B.; et al. Complete Genome Sequence and Characterization of a Polyethylene Biodegradation Strain, Streptomyces Albogriseolus LBX-2. Microorganisms 2019, 7, 379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, S.; Hiraga, K.; Takehana, T.; Taniguchi, I.; Yamaji, H.; Maeda, Y.; Toyohara, K.; Miyamoto, K.; Kimura, Y.; Oda, K. A Bacterium That Degrades and Assimilates Poly(Ethylene Terephthalate). Science 2016, 351, 6278. [Google Scholar] [CrossRef]
- Thoha, T.B.; Thomas, B.T.; Olanrewaju-Kehinde DS, K.; Popoola, O.D.; James, E.S. Degradation of Plastic and Polythene Materials by Some Selected Microorganisms Isolated from Soil. World Appl. Sci. J. 2015, 33, 1888–1891. [Google Scholar] [CrossRef]
- Skariyachan, S.; Manjunatha, V.; Sultana, S.; Jois, C.; Bai, V.; Vasist, K.S. Novel bacterial consortia isolated from plastic garbage processing areas demonstrated enhanced degradation for low density polyethylene. Environ. Sci. Pollut. Res. 2016, 23, 18307–18319. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, D.; Ghosh, R.; Sengupta, T.K. Biofilm-Mediated Enhanced Crude Oil Degradation by Newly Isolated Pseudomonas Species. ISRN Biotechnol. 2013, 2013, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mumtaz, T.; Khan, M.; Hassan, M.A. Study of environmental biodegradation of LDPE films in soil using optical and scanning electron microscopy. Micron 2010, 41, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Schlundt, C.; Welch, J.L.M.; Knochel, A.M.; Zettler, E.R.; Amaral-Zettler, L.A. Spatial structure in the “Plastisphere”: Molecular resources for imaging microscopic communities on plastic marine debris. Mol. Ecol. Resour. 2020, 20, 620–634. [Google Scholar] [CrossRef]
- Zumstein, M.T.; Schintlmeister, A.; Nelson, T.F.; Baumgartner, R.; Woebken, D.; Wagner, M.; Kohler, H.-P.E.; McNeill, K.; Sander, M. Biodegradation of synthetic polymers in soils: Tracking carbon into CO2 and microbial biomass. Sci. Adv. 2018, 4, eaas9024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package, R Package Version 2.5-6 (2019). Available online: https://CRAN.R-project.org/package=vegan (accessed on 11 May 2020).
- Villanueva, R.A.M.; Chen, Z.J.; Wickham, H. Ggplot2: Elegant Graphics for Data Analysis Using the Grammar of Graphics; Springer-Verlag: New York, NY, USA, 2016. [Google Scholar] [CrossRef]
- Burd, D. Plastic Not Fantastic. Project Reports of the Canada Wide Science Fair. 2008, pp. 1–5. Available online: https://www.academia.edu/download/46356573/08BurdReport.pdf (accessed on 11 May 2020).
- Morris, C.E.; Monier, J.-M.; Jacques, M.-A. A Technique to Quantify the Population Size and Composition of the Biofilm Component in Communities of Bacteria in the Phyllosphere. Appl. Environ. Microbiol. 1998, 64, 4789–4795. [Google Scholar] [CrossRef] [Green Version]
- Gilan, I.; Hadar, Y.; Sivan, A. Colonization, biofilm formation and biodegradation of polyethylene by a strain of Rhodococcus ruber. Appl. Microbiol. Biotechnol. 2004, 65, 97–104. [Google Scholar] [CrossRef]
- Abed, R.M.M.; Safi, N.M.D.; Köter, J.; de Beer, D.; El-Nahhal, Y.; Rullkötter, J.; Garcia-Pichel, F. Microbial Diversity of a Heavily Polluted Microbial Mat and Its Community Changes following Degradation of Petroleum Compounds. Appl. Environ. Microbiol. 2002, 68, 1674–1683. [Google Scholar] [CrossRef] [Green Version]
- Al-Hasan, R.H.; Al-Bader, D.A.; Sorkhoh, N.A.; Radwan, S.S. Evidence for nalkane consumption and oxidation by filamentous cyanobacteria from oil-contaminated coasts of the Arabian Gulf. Mar. Biol. 1998, 130, 521–527. [Google Scholar] [CrossRef]
- Roncero-Ramos, B.; Muñoz-Martín, M.; Cantón, Y.; Chamizo, S.; Rodríguez-Caballero, E.; Mateo, P. Land degradation effects on composition of pioneering soil communities: An alternative successional sequence for dryland cyanobacterial biocrusts. Soil Biol. Biochem. 2020, 146, 107824. [Google Scholar] [CrossRef]
- Abed, R.M.; Köster, J. The direct role of aerobic heterotrophic bacteria associated with cyanobacteria in the degradation of oil compounds. Int. Biodeterior. Biodegrad. 2005, 55, 29–37. [Google Scholar] [CrossRef]
- Miralles, I.; Lázaro, R.; Sánchez-Marañón, M.; Soriano, M.; Ortega, R. Biocrust cover and successional stages influence soil bacterial composition and diversity in semiarid ecosystems. Sci. Total Environ. 2020, 709, 134654. [Google Scholar] [CrossRef]
- Atlas, R.M.; Bartha, R. Degradation and mineralization of petroleum in sea water: Limitation by nitrogen and phosphorous. Biotechnol. Bioeng. 1972, 14, 309–318. [Google Scholar] [CrossRef]
- Head, I.M.; Jones, D.M.; Röling, W.F.M. Marine microorganisms make a meal of oil. Nat. Rev. Genet. 2006, 4, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Van Beilen, J.B.; Li, Z.; Duetz, W.A.; Smits, T.H.M.; Witholt, B. Diversity of Alkane Hydroxylase Systems in the Environment. Oil Gas Sci. Technol. 2003, 58, 427–440. [Google Scholar] [CrossRef]
- Danso, D.; Chow, J.; Streit, W.R. Plastics: Environmental and Biotechnological Perspectives on Microbial Degradation. Appl. Environ. Microbiol. 2019, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Restrepo-Flórez, J.-M.; Bassi, A.; Thompson, M.R. Microbial degradation and deterioration of polyethylene—A review. Int. Biodeterior. Biodegrad. 2014, 88, 83–90. [Google Scholar] [CrossRef]
- Weinstein, J.E.; Crocker, B.K.; Gray, A. From macroplastic to microplastic: Degradation of high-density polyethylene, polypropylene, and polystyrene in a salt marsh habitat. Environ. Toxicol. Chem. 2016, 35, 1632–1640. [Google Scholar] [CrossRef] [PubMed]
- Arutchelvi, J.; Sudhakar, M.; Arkatkar, A.; Doble, M.; Bhaduri, S.; Uppara, P.V. Biodegradation of Polyethylene and Polypropylene. Indian J. Biotechnol. 2008, 7, 9–22. [Google Scholar]
- Pramila, R. Biodegradation of low density polyethylene (LDPE) by fungi isolated from marine water—A SEM analysis. Afr. J. Microbiol. Res. 2011, 5, 131–136. [Google Scholar] [CrossRef]
- Das, M.P.; Kumar, S. An approach to low-density polyethylene biodegradation by Bacillus amyloliquefaciens. 3 Biotech 2015, 5, 81–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montazer, Z.; Najafi, M.B.H.; Levin, D.B. Challenges with Verifying Microbial Degradation of Polyethylene. Polymers 2020, 12, 123. [Google Scholar] [CrossRef] [Green Version]
- Zahra, S.; Abbas, S.S.; Mahsa, M.-T.; Mohsen, N. Biodegradation of low-density polyethylene (LDPE) by isolated fungi in solid waste medium. Waste Manag. 2010, 30, 396–401. [Google Scholar] [CrossRef]
- Bonhomme, S.; Cuer, A.; Delort, A.-M.; Lemaire, J.; Sancelme, M.; Scott, G. Environmental biodegradation of polyethylene. Polym. Degrad. Stab. 2003, 81, 441–452. [Google Scholar] [CrossRef]
- Santo, M.; Weitsman, R.; Sivan, A. The role of the copper-binding enzyme–laccase–in the biodegradation of polyethylene by the actinomycete Rhodococcus ruber. Int. Biodeterior. Biodegrad. 2013, 84, 204–210. [Google Scholar] [CrossRef]
- Yoon, M.G.; Jeon, H.J. Biodegradation of Polyethylene by a Soil Bacterium and AlkB Cloned Recombinant Cell. J. Bioremed. Biodegrad. 2012, 3. [Google Scholar] [CrossRef] [Green Version]
- Kirstein, I.V.; Wichels, A.; Gullans, E.; Krohne, G.; Gerdts, G. The Plastisphere—Uncovering tightly attached plastic “specific” microorganisms. PLoS ONE 2019, 14, e0215859. [Google Scholar] [CrossRef] [Green Version]
- Lucas, N.; Bienaime, C.; Belloy, C.; Queneudec, M.; Silvestre, F.; Nava-Saucedo, J.-E. Polymer biodegradation: Mechanisms and estimation techniques—A review. Chemosphere 2008, 73, 429–442. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MacLean, J.; Mayanna, S.; Benning, L.G.; Horn, F.; Bartholomäus, A.; Wiesner, Y.; Wagner, D.; Liebner, S. The Terrestrial Plastisphere: Diversity and Polymer-Colonizing Potential of Plastic-Associated Microbial Communities in Soil. Microorganisms 2021, 9, 1876. https://doi.org/10.3390/microorganisms9091876
MacLean J, Mayanna S, Benning LG, Horn F, Bartholomäus A, Wiesner Y, Wagner D, Liebner S. The Terrestrial Plastisphere: Diversity and Polymer-Colonizing Potential of Plastic-Associated Microbial Communities in Soil. Microorganisms. 2021; 9(9):1876. https://doi.org/10.3390/microorganisms9091876
Chicago/Turabian StyleMacLean, Joana, Sathish Mayanna, Liane G. Benning, Fabian Horn, Alexander Bartholomäus, Yosri Wiesner, Dirk Wagner, and Susanne Liebner. 2021. "The Terrestrial Plastisphere: Diversity and Polymer-Colonizing Potential of Plastic-Associated Microbial Communities in Soil" Microorganisms 9, no. 9: 1876. https://doi.org/10.3390/microorganisms9091876
APA StyleMacLean, J., Mayanna, S., Benning, L. G., Horn, F., Bartholomäus, A., Wiesner, Y., Wagner, D., & Liebner, S. (2021). The Terrestrial Plastisphere: Diversity and Polymer-Colonizing Potential of Plastic-Associated Microbial Communities in Soil. Microorganisms, 9(9), 1876. https://doi.org/10.3390/microorganisms9091876