
Genomic Insights into Drug Resistance Determinants in Cedecea neteri, A Rare Opportunistic Pathogen



Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Genomes
2.2. Sequence Database Search and Bioinformatics Tools
2.3. Comparative Genomic Analysis
3. Results and Discussion
3.1. Multiple Metallo-β-Lactamase-Encoding Genes in C. neteri

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Locus (cnt) | BLASTP Search Bacterium (% AA Sequence Identity) | Predicted Gene Product | Ambler Class | Predicted Subclass/Family Based on Sequence Fingerprints | Group-Specific Signatures/Conserved Residues |
|---|---|---|---|---|---|
| 00700 | kmi (96), cem (95) cen (95), clap (91) | Metallo-β-lactamase | B | B3/L1 * 198P-G-H-T-P-G203 | 129H-x-H-x-D-H134 D-97, L-112, G-203, H-224 |
| 03975 | cem (95), cen (95) kmi (95), clap (93) | Metallo-β-lactamase | B | NI | Lacks H-x-H-x-D-H motif D-85, D-87, H-113, H-186, H-220, G-228 |
| 10470 | cen (94), kmi (93) cem (93), clap (92) ear (76), enc (75) | AmpC (blaCMY/ACT) | C | CMY †/ACT 279R-Y-W-R-(v)-G-(s)-M-Y-Q288 | 85S-x-S-K88 171Y-A-N173 238D-A-E-A241 308S-D-N-K311 336K-T-G338 |
| 16535 | cem (94), cen (94) kmi (93), clap (93) | Metallo-β-lactamase | B | NI | 156H-x-H-x-D-H161 H-46, D-87, L-101, H-103, G-191, H-186, H-221 |
| 22070 | cen (90), cem (89) kmi (89), clap (89) ctu (69) | Metallo-β-lactamase | B | NI | 131H-x-H-x-D-H136 H-78, D-93, H-195, H-262, H-263 |
| 22350 | clap (93), opo (88) hav (87), kas (66), enc (65) | β-lactamase | Putative Class C | Novel variant | 144S-x-x-K147 |
3.2. A Novel β-Lactamase Gene in the SSMD04 Genome
3.3. CMY/ACT-Type AmpC β-Lactamase
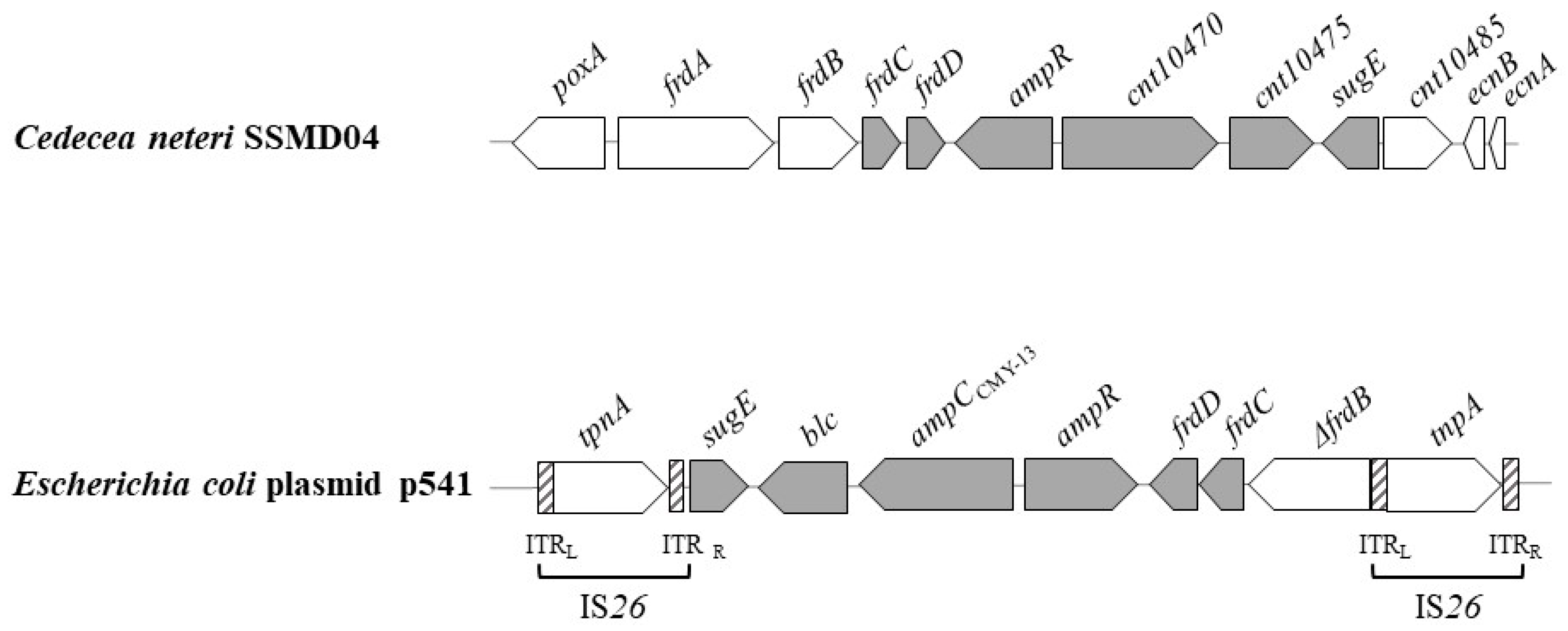
3.4. Genetic Environment of AmpC Gene in C. neteri SSMD04
3.5. Predicted Multidrug Efflux Pumps in C. neteri
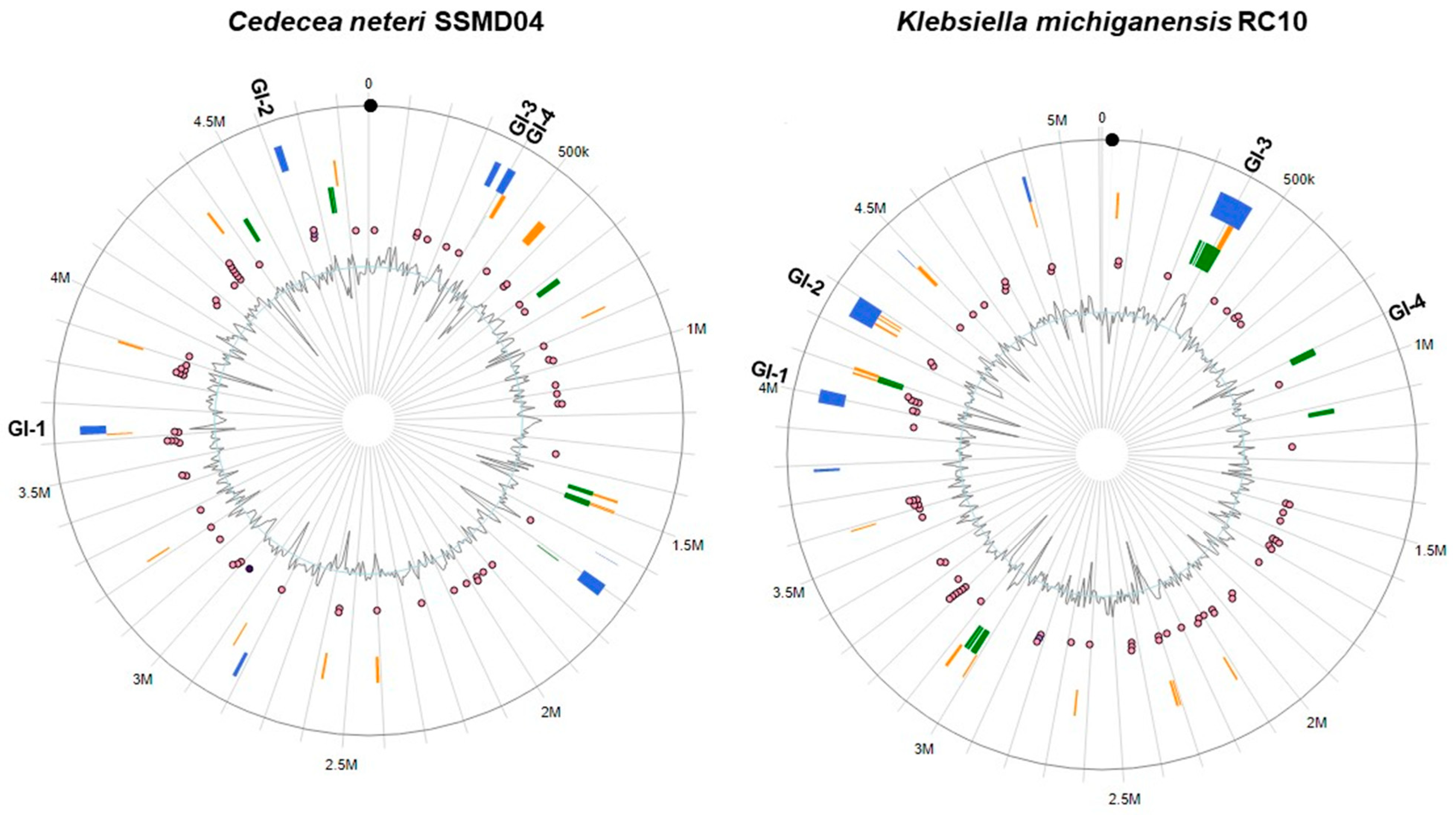
3.6. Comparison of Predicted Genomic Islands in C. neteri and K. michiganensis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grimont, P.; Grimont, F.; Farmer III, J.; Asbury, M. Cedecea davisae gen. nov., sp. nov. and Cedecea lapagei sp. nov., new Enterobacteriaceae from clinical specimens. Int. J. Syst. Bacteriol. 1981, 31, 317–326. [Google Scholar] [CrossRef] [Green Version]
- Dalamaga, M.; Vrioni, G. Cedecea. In Molecular Detection of Human Bacterial Pathogens; Liu, D., Ed.; CRC Press/Taylor and Francis Group: Boca Raton, FL, USA, 2011; pp. 817–825. [Google Scholar]
- Thompson, D.K.; Sharkady, S.M. Expanding spectrum of opportunistic Cedecea infections: Current clinical status and multidrug resistance. Int. J. Infect. Dis. 2020, 100, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Bae, B.H.C.; Sureka, S.B.; Ajamy, J.A. Enteric group 15 (Enterobacteriaceae) associated with pneumonia. J. Clin. Microbiol. 1981, 14, 596–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ismaael, T.G.; Zamora, E.M.; Khasawneh, F.A. Cedecea davisae’s role in a polymicrobial lung infection in a cystic fibrosis patient. Case Rep. Infect. Dis. 2012, 2012, 176864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, L.A.; Ibarra, B.S.; de la Garza, J.A.; Rada Fde, J.; Nuñez, A.I.; López, M.G. First reported case of pneumonia caused by Cedecea lapagei in America. Braz. J. Infect. Dis. 2013, 17, 626–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, S.K.; Lee, J.S.; Kim, E.C. First Korean case of Cedecea lapagei pneumonia in a patient with chronic obstructive pulmonary disease. Ann. Lan. Med. 2015, 35, 266–268. [Google Scholar] [CrossRef]
- Kury, C.M.H.; Yabrudi, A.A.; de Souza, T.B.; de Souza, E.C.; Silva Costa, L.T.E.; Soares, C.B.; Calixto, G.A.; Gramático, M.R. First reported case of ventilator-associated pneumonia and sepsis caused by Cedecea lapagei in a Brazilian neonatal intensive care unit. J. Pediatric Infect. Dis. Soc. 2017, 6, 209–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramaswamy, V.V.; Gummadapu, S.; Suryanarayana, N. Nosocomial pneumonia and sepsis caused by a rare organism Cedecea lapagei in an infant and a review of literature. BMJ Case Rep. 2019, 12, e229854. [Google Scholar] [CrossRef]
- Farmer III, J.J.; Sheth, N.K.; Hudzinski, J.A.; Rose, H.D.; Asbury, M.F. Bacteremia due to Cedecea neteri sp. nov. J. Clin. Microbiol. 1982, 16, 775–778. [Google Scholar] [CrossRef] [Green Version]
- Perkins, S.R.; Beckett, T.A.; Bump, C.M. Cedecea davisae bacteremia. J. Clin. Microbiol. 1986, 24, 675–676. [Google Scholar] [CrossRef] [Green Version]
- Aguilera, A.; Pascual, J.; Loza, E.; Lopez, J.; Garcia, G.; Liaño, F.; Quereda, C.; Ortuño, J. Bacteraemia with Cedecea neteri in a patient with systemic lupus erythematosus. Postgrad. Med. J. 1995, 71, 179–180. [Google Scholar] [CrossRef]
- Dalamaga, M.; Karmaniolas, K.; Arsenis, G.; Pantelaki, M.; Daskalopoulou, K.; Papadavid, E.; Migdalis, I. Cedecea lapagei bacteremia following cement-related chemical burn injury. Burns 2008, 34, 1205–1207. [Google Scholar] [CrossRef] [PubMed]
- Dalamaga, M.; Pantelaki, M.; Karmaniolas, K.; Matekovits, A.; Daskalopoulou, K. Leg ulcer and bacteremia due to Cedecea davisae. Eur. J. Dermatol. 2008, 18, 204–205. [Google Scholar] [CrossRef]
- Abate, G.; Qureshi, S.; Mazumder, S.A. Cedecea davisae bacteremia in a neutropenic patient with acute myeloid leukemia. J. Infect. 2011, 63, 83–85. [Google Scholar] [CrossRef]
- Akinosoglou, K.; Perperis, A.; Siagris, D.; Goutou, P.; Spiliopoulou, I.; Gogos, C.A.; Marangos, M. Bacteraemia due to Cedecea davisae in a patient with sigmoid colon cancer: A case report and brief review of the literature. Diagn. Microbiol. Infect. Dis. 2012, 74, 303–306. [Google Scholar] [CrossRef]
- Peretz, A.; Simsolo, C.; Farber, E.; Roth, A.; Brodsky, D.; Nakhoul, F. A rare bacteremia caused by Cedecea davisae in patient with chronic renal disease. Am. J. Case Rep. 2013, 14, 216–218. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, N.; Ali, S.M.; Khan, A.U. First reported New Delhi metallo-β-lactamase-1-producing Cedecea lapagei. Int. J. Antimicrob. Agents 2017, 49, 118–119. [Google Scholar] [CrossRef] [PubMed]
- Kanakadandi, V.S.; Sarao, M.S.; Cunningham, J.M. A rare case of Cedecea davisae bacteremia presenting as biliary sepsis. Cureus 2019, 11, e5298. [Google Scholar] [CrossRef] [Green Version]
- Mawardi, H.; Pavlakis, M.; Mandelbrot, D.; Woo, S.B. Sirolimus oral ulcer with Cedecea davisae superinfection. Transpl. Infect. Dis. 2010, 12, 446–450. [Google Scholar] [CrossRef] [PubMed]
- Biswal, I.; Hussain, N.A.; Grover, R.K. Cedecea lapagei in a patient with malignancy: Report of a rare case. J. Cancer Res. Ther. 2015, 11, 646. [Google Scholar] [CrossRef]
- Davis, O.; Wall, B.M. “Broom straw peritonitis” secondary to Cedecea lapagei in a liver transplant recipient. Perit. Dial. Int. 2006, 26, 512–513. [Google Scholar] [CrossRef]
- Ejaz, H.; Alzahrani, B.; Hamad, M.F.S.; Abosalif, K.O.A.; Junaid, K.; Abdalla, A.E.; Elamir, M.Y.M.; Aljaber, N.J.; Hamam, S.S.M.; Younas, S. Molecular analysis of the antibiotic resistant NDM-1 gene in clinical isolates of Enterobacteriaceae. Clin. Lab. 2020, 66, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Khalid, S.; Ahmad, N.; Ali, S.M.; Khan, A.U. Outbreak of efficiently transferred carbapenem-resistant blaNDM-producing gram-negative bacilli isolated from neonatal intensive care unit of an Indian hospital. Microb. Drug Resist. 2020, 26, 284–289. [Google Scholar] [CrossRef] [PubMed]
- Ammenouche, N.; Dupont, H.; Mammeri, H. Characterization of a novel AmpC β-lactamase produced by a carbapenem-resistant Cedecea davisae clinical isolate. Antimicrob. Agents Chemother. 2014, 58, 6942–6945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, H.; Masroor, T.; Parmar, S.A.; Panigrahi, D. Urinary tract infection by a rare pathogen Cedecea neteri in a pregnant female with polyhydramnios: Rare case report from UAE. BMC Infect. Dis. 2021, 21, 637. [Google Scholar] [CrossRef]
- Chan, K.G.; Tan, K.H.; Yin, W.F.; Tan, J.Y. Complete genome sequence of Cedecea neteri strain SSMD04, a bacterium isolated from pickled mackerel sashimi. Genome Announc. 2014, 2, e01339-14. [Google Scholar] [CrossRef] [Green Version]
- Chan, K.G.; Tan, W.S. Insights into Cedecea neteri strain M006 through complete genome sequence, a rare bacterium from aquatic environment. Stand. Genomic Sci. 2017, 12, 40. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Huerta-Cepas, J.; Serra, F.; Bork, P. ETE3: Reconstruction, analysis, and visualization of phylogenomic data. Mol. Biol. Evol. 2016, 33, 1635–1638. [Google Scholar] [CrossRef] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree: Computing large minimum evolution trees with profiles instead of a distance matrix. Mol. Biol. Evol. 2009, 26, 1641–1650. [Google Scholar] [CrossRef]
- Rodriguez-R, L.M.; Konstantinidis, K.T. The enveomics collection: A toolbox for specialized analyses of microbial genomes and metagenomes. PeerJ Preprints 2016, 4, e1900v1. [Google Scholar]
- Bertelli, C.; Laird, M.R.; Williams, K.P.; Simon Fraser University Research Computing Group; Lau, B.Y.; Hoad, G.; Winsor, G.L.; Brinkman, F.S.L. IslandViewer 4: Expanded prediction of genomic islands for larger-scale datasets. Nucleic Acids Res. 2017, 45, W30–W35. [Google Scholar] [CrossRef]
- Ginn, P.S.; Tart, S.B.; Sharkady, S.M.; Thompson, D.K. Urinary catheter colonization by multidrug-resistant Cedecea neteri in patient with benign prostatic hyperplasia. Case Rep. Infect. Dis. 2018, 7520527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Queenan, A.M.; Bush, K. Carbapenemases: The versatile beta-lactamases. Clin. Microbiol. Rev. 2007, 20, 440–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mojica, M.F.; Bonomo, R.A.; Fast, W. B1-metallo-β-lactamases: Where do we stand? Curr. Drug Targets 2016, 17, 1029–1050. [Google Scholar] [CrossRef]
- Singh, R.; Saxena, A.; Singh, H. Identification of group specific motifs in beta-lactamase family of proteins. J. Biomed. Sci. 2009, 16, 109. [Google Scholar] [CrossRef] [Green Version]
- Carfi, A.; Pares, S.; Duee, E.; Galleni, M.; Duez, C.; Frere, J.M.; Dideberg, O. The 3-D structure of zinc metallo-beta-lactamase from Bacillus cereus reveals a new type of protein fold. EMBO J. 1995, 14, 4914–4921. [Google Scholar] [CrossRef] [Green Version]
- Srivastava, A.; Singhal, N.; Goel, M.; Virdi, J.S.; Kumar, M. Identification of family specific fingerprints in β-lactamase families. Sci. World J. 2014, 980572. [Google Scholar] [CrossRef] [Green Version]
- Gudeta, D.D.; Pollini, S.; Docquier, S.D.; Bortolaia, V.; Rossolini, G.M.; Guardabassi, L. Biochemical characterization of CPS-1, a subclass B3 metallo-β-lactamase from a Chryseobacterium piscium soil isolate. Antimicrob. Agents Chemother. 2015, 60, 1869–1873. [Google Scholar] [CrossRef] [Green Version]
- Gudeta, D.D.; Bortolaia, V.; Amos, G.; Wellington, E.M.H.; Brandt, K.K.; Poirel, L.; Nielsen, J.B.; Westh, H.; Guardabassi, L. The soil microbiota harbors a diversity of carbapenem-hydrolyzing β-lactamases of potential clinical relevance. Antimicrob. Agents Chemother. 2016, 60, 151–160. [Google Scholar] [CrossRef] [Green Version]
- Ridell, J.; Siitonen, A.; Paulin, L.; Mattila, L.; Korkeala, H.; Albert, M.J. Hafnia alvei in stool specimens from patients with diarrhea and healthy controls. J. Clin. Microbiol. 1994, 32, 2335–2337. [Google Scholar] [CrossRef] [Green Version]
- Barry, J.W.; Dominguez, E.A.; Boken, D.J.; Preheim, L.C. Hafnia alvei infection after liver transplantation. Clin. Infect. Dis. 1997, 24, 1263–1264. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, G.A. AmpC β-lactamases. Clin. Microbiol. Rev. 2009, 22, 161–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahid, M.; Sobia, F.; Singh, A.; Khan, H.M.; Hawkey, P.M.; Huq, A.; Khardori, N. AmpC β-lactamases and bacterial resistance: An updated mini review. Rev. Med. Microbiol. 2009, 20, 41–55. [Google Scholar] [CrossRef]
- Rodríguez-Martínez, J.M.; Poirel, L.; Nordmann, P. Extended-spectrum cephalosporinases in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2009, 53, 1766–1771. [Google Scholar] [CrossRef] [Green Version]
- Bebrone, C.; Moali, C.; Mahy, F.; Rival, S.; Docquier, J.D.; Rossolini, G.M.; Fastrez, J.; Pratt, R.F.; Frère, J.M.; Galleni, M. CENTA as a chromogenic substrate for studying β-lactamases. Antimicrob. Agents Chemother. 2001, 45, 1868–1871. [Google Scholar] [CrossRef] [Green Version]
- Miriagou, V.; Tzouvelekis, L.S.; Villa, L.; Lebessi, E.; Vatopoulos, A.C.; Carattoli, A.; Tzelepi, E. CMY-13, a novel inducible cephalosporinase encoded by an Escherichia coli plasmid. Antimicrob. Agents Chemother. 2004, 48, 3172–3174. [Google Scholar] [CrossRef] [Green Version]
- Caille, O.; Zincke, D.; Merighi, M.; Balasubramanian, D.; Kumari, H.; Kong, K.F.; Silva-Herzog, E.; Narasimhan, G.; Schneper, L.; Lory, S.; et al. Structural and functional characterization of Pseudomonas aeruginosa global regulator AmpR. J. Bacteriol. 2014, 196, 3890–3902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, Y.J.; Saier, M.H. Overexpression of the Escherichia coli sugE gene confers resistance to a narrow range of quaternary ammonium compounds. J. Bacteriol. 2002, 184, 2543–2545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, C.; Joris, B.; Jamin, M.; Klarsov, K.; Van Beeumen, J.; Mengin-Lecreulx, D.; van Heijenoort, J.; Park, J.T.; Normark, S.; Frère, J.M. AmpD, essential for both beta-lactamase regulation and cell wall recycling, is a novel cytosolic N-acetylmuramyl-L-alanine amidase. Mol. Microbiol. 1995, 15, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Dik, D.A.; Fisher, J.F.; Mobashery, S. Cell-wall recycling of the gram-negative bacteria and the nexus to antibiotic resistance. Chem. Rev. 2018, 118, 5952–5984. [Google Scholar] [CrossRef]
- Honoré, N.; Nicolas, M.H.; Cole, S.T. Regulation of enterobacterial cephalosporinase production: The role of a membrane bound sensory transducer. Mol. Microbiol. 1989, 3, 1121–1130. [Google Scholar] [CrossRef]
- Lindquist, S.; Galleni, M.; Lindberg, F.; Normark, S. Signalling proteins in enterobacterial AmpC β-lactamase regulation. Mol. Microbol. 1989, 3, 1091–1102. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Z.; Nikaido, H. Efflux-mediated drug resistance in bacteria: An update. Drugs 2009, 69, 1555–1623. [Google Scholar] [CrossRef]
- Blair, J.M.A.; Richmond, G.E.; Piddock, L.J.V. Multidrug efflux pumps in gram-negative bacteria and their role in antibiotic resistance. Future Microbiol. 2014, 9, 1165–1177. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, H.; Ning, J.; Sajid, A.; Cheng, G.; Yuan, Z.; Hao, H. The nature and epidemiology of OqxAB, a multidrug efflux pump. Antimicrob. Resist. Infect. Control 2019, 8, 44. [Google Scholar] [CrossRef] [Green Version]
- Nikaido, H.; Takatsuka, Y. Mechanisms of RND multidrug efflux pumps. Biochim. Biophys. Acta. 2009, 1794, 769–781. [Google Scholar] [CrossRef] [Green Version]
- Fralick, J.A. Evidence that TolC is required for functioning of the Mar/AcrAB efflux pump of Escherichia coli. J. Bacteriol. 1996, 178, 5803–5805. [Google Scholar] [CrossRef] [Green Version]
- Nishino, K.; Yamada, J.; Hirakawa, H.; Hirata, T.; Yamaguchi, A. Roles of TolC-dependent multidrug transporters of Escherichia coli in resistance to β-lactams. Antimicrob. Agents Chemother. 2003, 47, 3030–3033. [Google Scholar] [CrossRef] [Green Version]
- Anes, J.; McCusker, M.P.; Fanning, S.; Martins, M. The ins and outs of RND efflux pumps in Escherichia coli. Front. Microbiol. 2015, 6, 587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hobbs, E.C.; Yin, X.; Paul, B.J.; Astarita, J.L.; Storz, G. Conserved small protein associates with the multidrug efflux pump AcrB and differentially affects antibiotic resistance. Proc. Natl. Acad. Sci. USA 2012, 109, 16696–16701. [Google Scholar] [CrossRef] [Green Version]
- Rosenberg, E.Y.; Ma, D.; Nikaido, H. AcrD of Escherichia coli is an aminoglycoside efflux pump. J. Bacteriol. 2000, 182, 1754–1756. [Google Scholar] [CrossRef] [Green Version]
- Nishino, K.; Yamaguchi, A. Analysis of a complete library of putative drug transporter genes in Escherichia coli. J. Bacteriol. 2001, 183, 5803–5812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baranova, N.; Nikaido, H. The BaeSR two-component regulatory system activates transcription of the yegMNOB (mdtABCD) transporter gene cluster in Escherichia coli and increases its resistance to novobiocin and deoxycholate. J. Bacteriol. 2002, 184, 4168–4176. [Google Scholar] [CrossRef] [Green Version]
- Nagakubo, S.; Nishino, K.; Hirata, T.; Yamaguchi, A. The putative response regulator BaeR stimulates multidrug resistance of Escherichia coli via a novel multidrug exporter system, MdtABC. J. Bacteriol. 2002, 184, 4161–4167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lomovskaya, O.; Lewis, K. emr, an Escherichia coli locus for multidrug resistance. Proc. Natl. Acad. Sci. USA 1992, 89, 8938–8942. [Google Scholar] [CrossRef] [Green Version]
- Lomovskaya, O.; Lewis, K.; Matin, A. EmrR is a negative regulator of the Escherichia coli multidrug resistance pump EmrAB. J. Bacteriol. 1995, 177, 2328–2334. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.F.; Lin, Y.Y.; Lan, C.Y. Contribution of EmrAB efflux pumps to colistin resistance in Acinetobacter baumannii. J. Microbiol. 2017, 55, 130–136. [Google Scholar] [CrossRef]
- Seiffert, S.N.; Wüthrich, D.; Gerth, Y.; Egli, A.; Kohler, P.; Nolte, O. First clinical case of KPC-3-producing Klebsiella michiganensis in Europe. New Microbes New Infect. 2019, 29, 100516. [Google Scholar] [CrossRef]
- Pimenta, A.L.; Racher, K.; Jamieson, L.; Blight, M.A.; Holland, I.B. Mutations in HlyD, part of the type 1 translocator for hemolysin secretion, affect the folding of the secreted toxin. J. Bacteriol. 2005, 187, 7471–7480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanonenberg, K.; Spitz, O.; Erenburg, I.N.; Beer, T.; Schmitt, L. Type I secretion system—it takes three and a substrate. FEMS Microbiol. Lett. 2018, 365, fny094. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.; Holland, I.B.; Schmitt, L. The type 1 secretion pathway – the hemolysin system and beyond. Biochim. Biophys. Acta. 2014, 1843, 1629–1641. [Google Scholar] [CrossRef] [Green Version]
- Atac, N.; Kurt-Azap, O.; Dolapci, I.; Yesilkaya, A.; Ergonul, O.; Gonen, M.; Can, F. The role of AcrAB-TolC efflux pumps on quinolone resistance of E. coli ST131. Curr. Microbiol. 2018, 75, 1661–1666. [Google Scholar] [CrossRef] [Green Version]
- Rosenberg, E.Y.; Bertenthal, D.; Nilles, M.; Bertrand, K.P.; Nikaido, H. Bile salts and fatty acids induce the expression of Escherichia coli AcrAB multidrug efflux pump through their interaction with Rob regulatory protein. Mol. Microbiol. 2003, 48, 1609–1619. [Google Scholar] [CrossRef] [PubMed]
- Thanassi, D.G.; Cheng, L.W.; Nikaido, H. Active efflux of bile salts by Escherichia coli. J. Bacteriol. 1997, 179, 2512–2518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilson, L.; Mahanty, H.K.; Kolter, R. Genetic analysis of an MDR-like export system: The secretion of colicin V. EMBO J. 1990, 9, 3875–3884. [Google Scholar] [CrossRef]
- Vakharia, H.; German, G.J.; Misra, R. Isolation and characterization of Escherichia coli tolC mutants defective in screening enzymatically active alpha-hemolysin. J. Bacteriol. 2001, 183, 6908–6916. [Google Scholar] [CrossRef] [Green Version]
- Wandersman, C.; Delepelaire, P. TolC, an Escherichia coli outer membrane protein required for hemolysin secretion. Proc. Natl. Acad. Sci. USA 1990, 87, 4776–4780. [Google Scholar] [CrossRef] [Green Version]
- Zgurskaya, H.I.; Krishnamoorthy, G.; Ntreh, A.; Lu, S. Mechanism and function of the outer membrane channel TolC in multidrug resistance and physiology of Enterobacteria. Front. Microbiol. 2011, 2, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yousefian, N.; Ornik-Cha, A.; Poussard, S.; Decossas, M.; Berbon, M.; Daury, L.; Taveau, J.C.; Dupuy, J.W.; Dordević-Marquardt, S.; Lambert, O.; et al. Structural characterization of the EmrAB-TolC efflux complex from E. coli. Biochim. Biophys. Acta Biomembr. 2021, 1863, 183488. [Google Scholar] [CrossRef] [PubMed]
- Worley, M.J.; Ching, K.H.; Heffron, F. Salmonella SsrB activates a global regulon of horizontally acquired genes. Mol. Microbiol. 2000, 36, 749–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Hou, X.; Sun, Y.; Zhang, G.; Hu, X.; Xie, Y.; Mo, X.; Ding, X.; Xia, L.; Hu, S. Screening a fosmid library of Xenorhabdus stockiae HN_xs01 reveals SrfABC toxin that exhibits both cytotoxicity and injectable insecticidal activity. J. Invertebr. Pathol. 2019, 167, 107247. [Google Scholar] [CrossRef]
- Sun, Y.; Zhang, G.; Hou, X.; Xiao, S.; Yang, X.; Xie, Y.; Huang, X.; Wang, F.; Mo, X.; Ding, X.; et al. SrfABC toxin from Xenorhabdus stockiae induces cytotoxicity and apoptosis in HeLa cells. Toxins 2019, 11, 685. [Google Scholar] [CrossRef] [Green Version]


| Strain | Chromosome Size (bp) | Protein-Coding Genes | Source | Reference |
|---|---|---|---|---|
| Cedecea neteri SSMD04 | 4,876,443 | 4318 | Pickled mackerel sashimi | [27] |
| Cedecea neteri M006 | 4,965,436 | 4423 | Malaysian waterfall | [28] |
| Cedecea neteri ND14a | 4,659,311 | 4141 | Malaysian waterfall | Unpublished |
| Klebsiella michiganensis RC10 | 5,107,633 | 4536 | Rice field | Unpublished |
| C. neteri SSMD04 | K. michiganensis RC10 | ||||
|---|---|---|---|---|---|
| Gene Locus | GenBank Accession No. | Gene Product | Gene Locus | GenBank Accession No. | Gene Product |
| JT31_RS16805 | WP_038479661.1 | Transposase | VW41_RS18405 | WP_045783700.1 | Outer membrane protein assembly factor BamE |
| JT31_RS16810 | None | Integrase | VW41_RS18410 | WP_045783701.1 | RnfH family protein |
| JT31_RS16815 | WP_038479664.1 | Secretion protein HlyD | VW41_RS18415 | WP_045783702.1 | Ubiquinone-binding protein |
| JT31_RS16820 | WP_038479667.1 | ATP-binding protein | VW41_RS18420 | WP_008458185.1 | SsrA-binding protein |
| JT31_RS16825 | WP_038479668.1 | Type I secretion protein TolC | VW41_RS18425 | WP_045783703.1 | Large repetitive protein |
| JT31_RS16830 | WP_038479669.1 | Large repetitive protein | VW41_RS18430 | WP_045783704.1 | Type I secretion protein TolC |
| JT31_RS16835 | WP_038479670.1 | SsrA-binding protein | VW41_RS18435 | WP_045783705.1 | ATP-binding protein |
| JT31_RS16840 | WP_038479681.1 | Ubiquinone-binding protein | VW41_RS18440 | WP_045783706.1 | HlyD family type I secretion periplasmic adaptor subunit |
| JT31_RS16845 | WP_038479684.1 | RnfH family protein | VW41_RS18445 | WP_045783707.1 | Integrase |
| JT31_RS16850 | WP_038479687.1 | Outer membrane protein assembly factor BamE | VW41_RS18450 | WP_045783708.1 | ATP-dependent DNA helicase |
| JT31_RS23250 | WP_071842976.1 | Hypothetical protein | VW41_RS18455 | WP_052699081.1 | Chromosome segregation protein SMC |
| JT31_RS16855 | WP_038479690.1 | DNA repair protein RecN | VW41_RS18460 | WP_045783709.1 | Hypothetical protein |
| VW41_RS18465 | WP_045783710.1 | Hypothetical protein | |||
| VW41_RS18470 | WP_045783711.1 | Hypothetical protein | |||
| VW41_RS18475 | WP_045783712.1 | Hypothetical protein | |||
| VW41_RS24485 | None | Relaxase | |||
| VW41_RS18480 | WP_045783713.1 | Transposase | |||
| VW41_RS18485 | WP_071844405.1 | Transposase | |||
| VW41_RS18490 | WP_045783715.1 | Hypothetical protein | |||
| VW41_RS18495 | WP_045783716.1 | Hypothetical protein | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thompson, D.K.; Sharkady, S.M. Genomic Insights into Drug Resistance Determinants in Cedecea neteri, A Rare Opportunistic Pathogen. Microorganisms 2021, 9, 1741. https://doi.org/10.3390/microorganisms9081741
Thompson DK, Sharkady SM. Genomic Insights into Drug Resistance Determinants in Cedecea neteri, A Rare Opportunistic Pathogen. Microorganisms. 2021; 9(8):1741. https://doi.org/10.3390/microorganisms9081741
Chicago/Turabian StyleThompson, Dorothea K., and Stephen M. Sharkady. 2021. "Genomic Insights into Drug Resistance Determinants in Cedecea neteri, A Rare Opportunistic Pathogen" Microorganisms 9, no. 8: 1741. https://doi.org/10.3390/microorganisms9081741
APA StyleThompson, D. K., & Sharkady, S. M. (2021). Genomic Insights into Drug Resistance Determinants in Cedecea neteri, A Rare Opportunistic Pathogen. Microorganisms, 9(8), 1741. https://doi.org/10.3390/microorganisms9081741