
Minimal Associations between Short-Term Dietary Intake and Salivary Microbiome Composition

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Dietary Intake
2.3. Microbiome Analyses
2.4. Statistical Analysis
3. Results
3.1. Study Population Characteristics
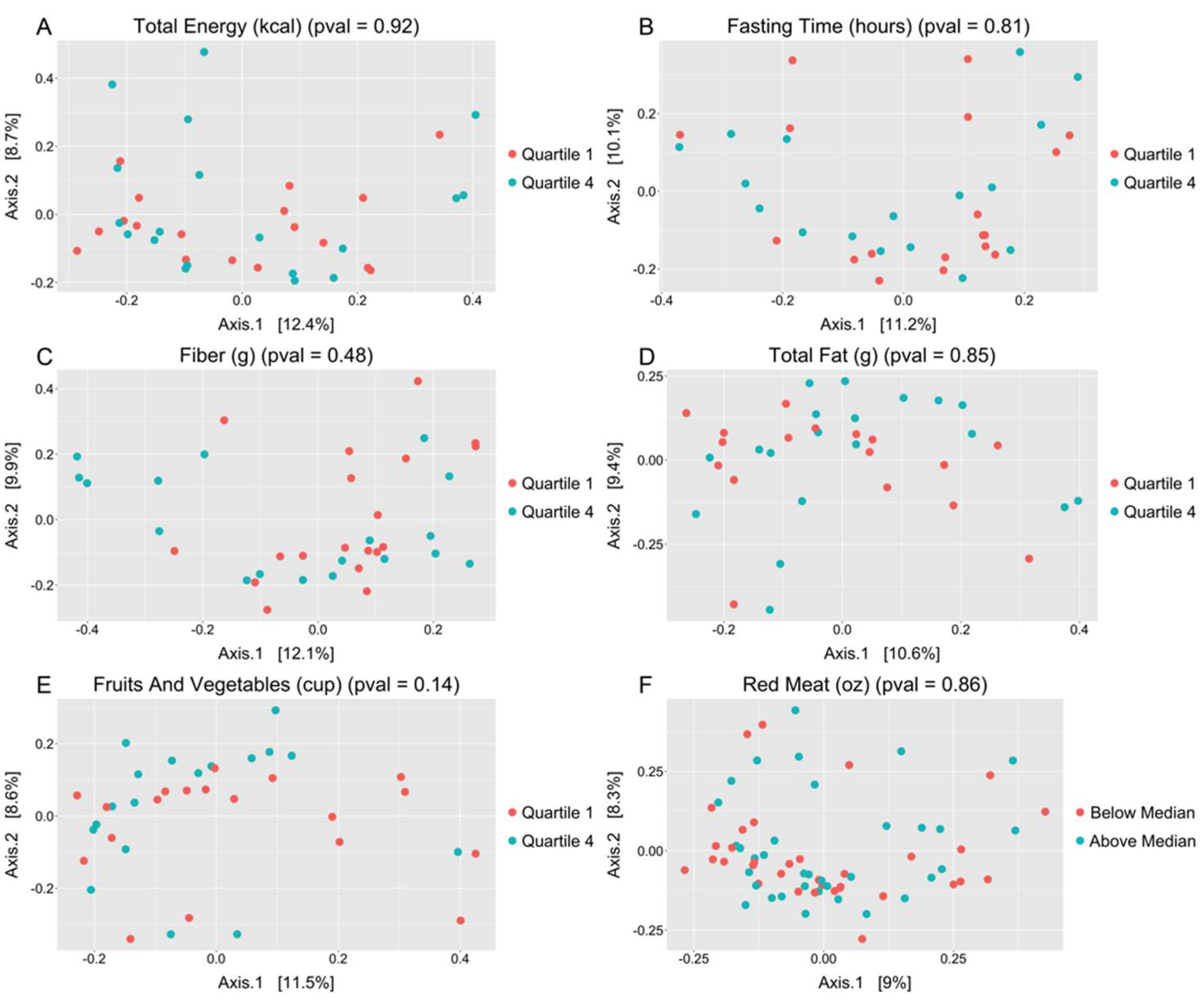
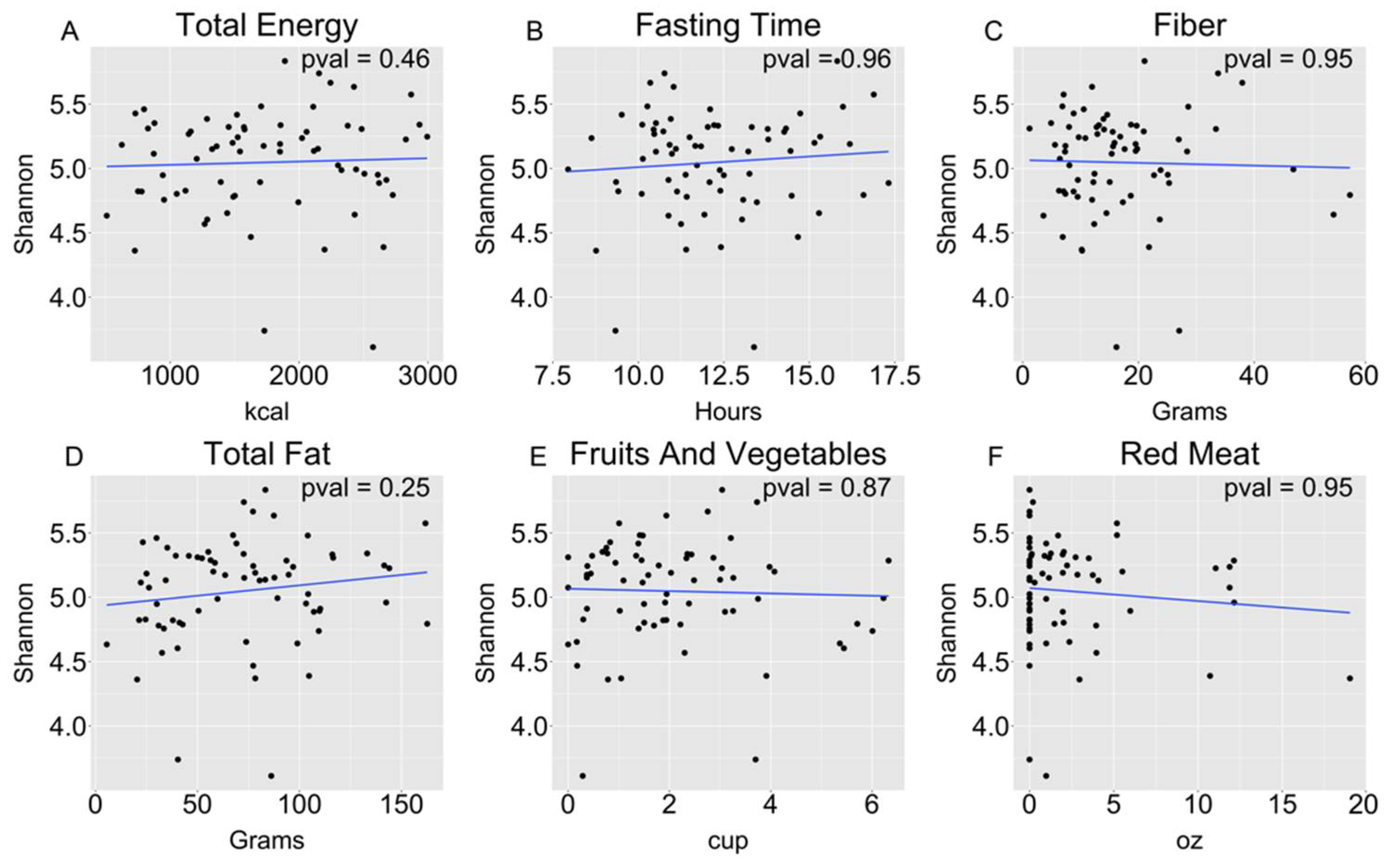
3.2. Salivary Microbiome and Dietary Intake
3.3. Salivary Microbiome and Clinical Features
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Segata, N.; Haake, S.K.; Mannon, P.; Lemon, K.P.; Waldron, L.; Gevers, D.; Huttenhower, C.; Izard, J. Composition of the adult digestive tract bacterial microbiome based on seven mouth surfaces, tonsils, throat and stool samples. Genome Biol. 2012, 13, R42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Francois, F.; Pei, Z. Molecular Pathways: Pathogenesis and Clinical Implications of Microbiome Alteration in Esophagitis and Barrett Esophagus: Figure 1. Clin. Cancer Res. 2012, 18, 2138–2144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annavajhala, M.K.; May, M.; Compres, G.; Freedberg, D.E.; Graham, R.; Stump, S.; Que, J.; Korem, T.; Uhlemann, A.-C.; Abrams, J.A. Relationship of the Esophageal Microbiome and Tissue Gene Expression and Links to the Oral Microbiome: A Randomized Clinical Trial. Clin. Transl. Gastroenterol. 2020, 11, e00235. [Google Scholar] [CrossRef] [PubMed]
- Snider, E.J.; Compres, G.; Freedberg, D.E.; Giddins, M.J.; Khiabanian, H.; Lightdale, C.J.; Nobel, Y.R.; Toussaint, N.; Uhlemann, A.-C.; Abrams, J.A. Barrett’s esophagus is associated with a distinct oral microbiome. Clin. Transl. Gastroenterol. 2018, 9, e135. [Google Scholar] [CrossRef]
- Peters, B.; Wu, J.; Pei, Z.; Yang, L.; Purdue, M.P.; Freedman, N.D.; Jacobs, E.J.; Gapstur, S.M.; Hayes, R.; Ahn, J. Oral Microbiome Composition Reflects Prospective Risk for Esophageal Cancers. Cancer Res. 2017, 77, 6777–6787. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Winckler, B.; Lu, M.; Cheng, H.; Yuan, Z.; Yang, Y.; Jin, L.; Ye, W. Oral Microbiota and Risk for Esophageal Squamous Cell Carcinoma in a High-Risk Area of China. PLoS ONE 2015, 10, e0143603. [Google Scholar] [CrossRef]
- Yu, G.; Gail, M.H.; Shi, J.; Klepac-Ceraj, V.; Paster, B.J.; Dye, B.A.; Wang, G.-Q.; Wei, W.-Q.; Fan, J.-H.; Qiao, Y.-L.; et al. Association between Upper Digestive Tract Microbiota and Cancer-Predisposing States in the Esophagus and Stomach. Cancer Epidemiol. Biomark. Prev. 2014, 23, 735–741. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Yuan, Z.; Lu, M.; Zhang, Y.; Jin, L.; Ye, W. Poor oral health is associated with an increased risk of esophageal squamous cell carcinoma—A population-based case-control study in China. Int. J. Cancer 2016, 140, 626–635. [Google Scholar] [CrossRef]
- Menya, D.; Maina, S.K.; Kibosia, C.; Kigen, N.; Oduor, M.; Some, F.; Chumba, D.; Ayuo, P.; Middleton, D.R.; Osano, O.; et al. Dental fluorosis and oral health in the African Esophageal Cancer Corridor: Findings from the Kenya ESCCAPE case–control study and a pan-African perspective. Int. J. Cancer 2018, 145, 99–109. [Google Scholar] [CrossRef] [Green Version]
- Vogtmann, E.; Hua, X.; Zhou, L.; Wan, Y.; Suman, S.; Zhu, B.; Dagnall, C.L.; Hutchinson, A.; Jones, K.; Hicks, B.D.; et al. Temporal Variability of Oral Microbiota over 10 Months and the Implications for Future Epidemiologic Studies. Cancer Epidemiol. Biomark. Prev. 2018, 27, 594–600. [Google Scholar] [CrossRef] [Green Version]
- Belstrøm, D.; Holmstrup, P.; Bardow, A.; Kokaras, A.; Fiehn, N.-E.; Paster, B.J. Temporal Stability of the Salivary Microbiota in Oral Health. PLoS ONE 2016, 11, e0147472. [Google Scholar] [CrossRef] [PubMed]
- Cameron, S.; Huws, S.; Hegarty, M.J.; Smith, D.; Mur, L.A.J. The human salivary microbiome exhibits temporal stability in bacterial diversity. FEMS Microbiol. Ecol. 2015, 91, fiv091. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Lauber, C.L.; Costello, E.K.; Berg-Lyons, D.; Gonzalez, A.; Stombaugh, J.; Knights, D.; Gajer, P.; Ravel, J.; Fierer, N.; et al. Moving pictures of the human microbiome. Genome Biol. 2011, 12, R50. [Google Scholar] [CrossRef] [Green Version]
- Cabral, D.; Wurster, J.; Flokas, M.E.; Alevizakos, M.; Zabat, M.; Korry, B.J.; Rowan-Nash, A.; Sano, W.; Andreatos, N.; Ducharme, R.B.; et al. The salivary microbiome is consistent between subjects and resistant to impacts of short-term hospitalization. Sci. Rep. 2017, 7, 11040. [Google Scholar] [CrossRef] [Green Version]
- Lim, Y.; Totsika, M.; Morrison, M.; Punyadeera, C. The saliva microbiome profiles are minimally affected by collection method or DNA extraction protocols. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef] [Green Version]
- Cotillard, A.; Kennedy, S.; Kong, L.C.; Prifti, E.; Pons, N.; Le Chatelier, E.; Almeida, M.; Quinquis, B.; Levenez, F.; Galleron, N.; et al. Dietary intervention impact on gut microbial gene richness. Nat. Cell Biol. 2013, 500, 585–588. [Google Scholar] [CrossRef]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.-Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking Long-Term Dietary Patterns with Gut Microbial Enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [Green Version]
- Zeng, H. Mechanisms linking dietary fiber, gut microbiota and colon cancer prevention. World J. Gastrointest. Oncol. 2014, 6, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Haro, C.; Montes-Borrego, M.; Rangel-Zuñiga, O.A.; Alcala-Diaz, J.F.; Delgado, F.G.; Pérez-Martínez, P.; Delgado-Lista, J.; Quintana-Navarro, G.M.; Tinahones, F.J.; Landa, B.B.; et al. Two Healthy Diets Modulate Gut Microbial Community Improving Insulin Sensitivity in a Human Obese Population. J. Clin. Endocrinol. Metab. 2016, 101, 233–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nat. Cell Biol. 2013, 505, 559–563. [Google Scholar] [CrossRef] [Green Version]
- Belstrøm, D.; Holmstrup, P.; Nielsen, C.H.; Kirkby, N.; Twetman, S.; Heitmann, B.L.; Klepac-Ceraj, V.; Paster, B.J.; Fiehn, N.-E. Bacterial profiles of saliva in relation to diet, lifestyle factors, and socioeconomic status. J. Oral Microbiol. 2014, 6. [Google Scholar] [CrossRef] [Green Version]
- Kato, I.; Vasquez, A.; Moyerbrailean, G.; Land, S.; Djuric, Z.; Sun, J.; Lin, H.-S.; Ram, J.L. Nutritional Correlates of Human Oral Microbiome. J. Am. Coll. Nutr. 2016, 36, 88–98. [Google Scholar] [CrossRef] [Green Version]
- Subar, A.F.; Kirkpatrick, S.I.; Mittl, B.; Zimmerman, T.P.; Thompson, F.E.; Bingley, C.; Willis, G.; Islam, N.G.; Baranowski, T.; McNutt, S.; et al. The Automated Self-Administered 24-Hour Dietary Recall (ASA24): A Resource for Researchers, Clinicians, and Educators from the National Cancer Institute. J. Acad. Nutr. Diet. 2012, 112, 1134–1137. [Google Scholar] [CrossRef] [Green Version]
- Hansen, T.H.; Kern, T.; Bak, E.G.; Kashani, A.; Allin, K.; Nielsen, T.; Hansen, T.; Pedersen, O. Impact of a vegan diet on the human salivary microbiota. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef]
- Takeshita, T.; Kageyama, S.; Furuta, M.; Tsuboi, H.; Takeuchi, K.; Shibata, Y.; Shimazaki, Y.; Akifusa, S.; Ninomiya, T.; Kiyohara, Y.; et al. Bacterial diversity in saliva and oral health-related conditions: The Hisayama Study. Sci. Rep. 2016, 6, 22164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Chi, X.; Zhang, Q.; Chen, F.; Deng, X. Characterization of the salivary microbiome in people with obesity. PeerJ 2018, 6, e4458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Filippis, F.; Vannini, L.; La Storia, A.; Laghi, L.; Piombino, P.; Stellato, G.; Serrazanetti, D.I.; Gozzi, G.; Turroni, S.; Ferrocino, I.; et al. The Same Microbiota and a Potentially Discriminant Metabolome in the Saliva of Omnivore, Ovo-Lacto-Vegetarian and Vegan Individuals. PLoS ONE 2014, 9, e112373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viljakainen, J.; Raju, S.; Viljakainen, H.; Figueiredo, R.; Roos, E.; Weiderpass, E.; Rounge, T.B. Meal Regularity Plays a Role in Shaping the Saliva Microbiota. Front. Microbiol. 2020, 11, 757. [Google Scholar] [CrossRef]
- Nearing, J.T.; DeClercq, V.; Van Limbergen, J.; Langille, M.G.I. Assessing the Variation within the Oral Microbiome of Healthy Adults. mSphere 2020, 5. [Google Scholar] [CrossRef]
- Börnigen, D.; Ren, B.; Pickard, R.; Li, J.; Ozer, E.; Hartmann, E.M.; Xiao, W.; Tickle, T.; Rider, J.; Gevers, D.; et al. Alterations in oral bacterial communities are associated with risk factors for oral and oropharyngeal cancer. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Metzker, M.L. Sequencing technologies—The next generation. Nat. Rev. Genet. 2010, 11, 31–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
| (n = 69) | |
|---|---|
| Age, median (IQR) | 61 (42–71) |
| BMI, median (IQR) | 26.6 (22.3–29.2) |
| Male sex | 45 (65%) |
| PPI use | 12 (17%) |
| Smoking | |
| Current | 2 (3%) |
| Former | 22 (32%) |
| Never | 45 (65%) |
| Oral health and hygiene | |
| Tooth loss status | |
| Has all or most of natural teeth | 44 (64%) |
| Has partial plates or implants | 20 (29%) |
| Has full upper and lower dentures or implants | 2 (3%) |
| Has full upper dentures or implants | 2 (3%) |
| Unknown | 1 (1%) |
| Tooth brushing | |
| Daily | 25 (36%) |
| Less than daily | 3 (4%) |
| More than daily | 41 (59%) |
| Brush on day of collection—yes | 59 (86%) |
| Mouthwash in the past month n (%) | |
| Daily | 19 (28%) |
| Less than daily | 19 (28%) |
| Does not use | 31 (44%) |
| Day of collection—yes | 19 (28%) |
| Total Calories (kcal) | |
|---|---|
| Median (interquartile range) | 1698 (1268–2302) |
| Range | 508.7–2993.51 |
| Total Fat (g) | |
| Median (interquartile range) | 72.7 (40.2–96.9) |
| Range | 5.5–162.6 |
| Fiber (g) | |
| Median (interquartile range) | 14.5 (9.5–20.9) |
| Range | 1.2–56.6 |
| Fruit/Veg (cups) | |
| Median (interquartile range) | 1.8 (0.8–3.0) |
| Range | 0.0–6.3 |
| Red Meat (oz) | |
| Median (interquartile range) | 0.99 (0–2.8) |
| Range | 0–19.0 |
| Fasting Time (hours) | |
| Median (interquartile range) | 12 (11–13) |
| Range | 8–17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.; Lee, M.; Baldwin-Hunter, B.; Solfisburg, Q.S.; Lightdale, C.J.; Korem, T.; Hur, C.; Abrams, J.A. Minimal Associations between Short-Term Dietary Intake and Salivary Microbiome Composition. Microorganisms 2021, 9, 1739. https://doi.org/10.3390/microorganisms9081739
Kim J, Lee M, Baldwin-Hunter B, Solfisburg QS, Lightdale CJ, Korem T, Hur C, Abrams JA. Minimal Associations between Short-Term Dietary Intake and Salivary Microbiome Composition. Microorganisms. 2021; 9(8):1739. https://doi.org/10.3390/microorganisms9081739
Chicago/Turabian StyleKim, Judith, Minyi Lee, Brittany Baldwin-Hunter, Quinn S. Solfisburg, Charles J. Lightdale, Tal Korem, Chin Hur, and Julian A. Abrams. 2021. "Minimal Associations between Short-Term Dietary Intake and Salivary Microbiome Composition" Microorganisms 9, no. 8: 1739. https://doi.org/10.3390/microorganisms9081739
APA StyleKim, J., Lee, M., Baldwin-Hunter, B., Solfisburg, Q. S., Lightdale, C. J., Korem, T., Hur, C., & Abrams, J. A. (2021). Minimal Associations between Short-Term Dietary Intake and Salivary Microbiome Composition. Microorganisms, 9(8), 1739. https://doi.org/10.3390/microorganisms9081739