
Pathogenicity and Metabolites of Endoparasitic Nematophagous Fungus Drechmeria coniospora YMF1.01759 against Nematodes

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Normal Culture
2.2. General Experimental Instruments
2.3. Assaying of Nematicidal Activity
2.3.1. Pathogenicity of D. coniospora YMF1.01759 against C. elegans
2.3.2. Nematicidal Activity of Extracts of D. coniospora YMF1.01759 on Different Media
2.3.3. Nematicidal Activity of Compounds
2.3.4. Inhibition of Egg Hatching Activity of M. incognita
2.4. Extraction and Isolation of Metabolites from D. coniospora YMF1.01759
2.5. Statistical Analysis
3. Results
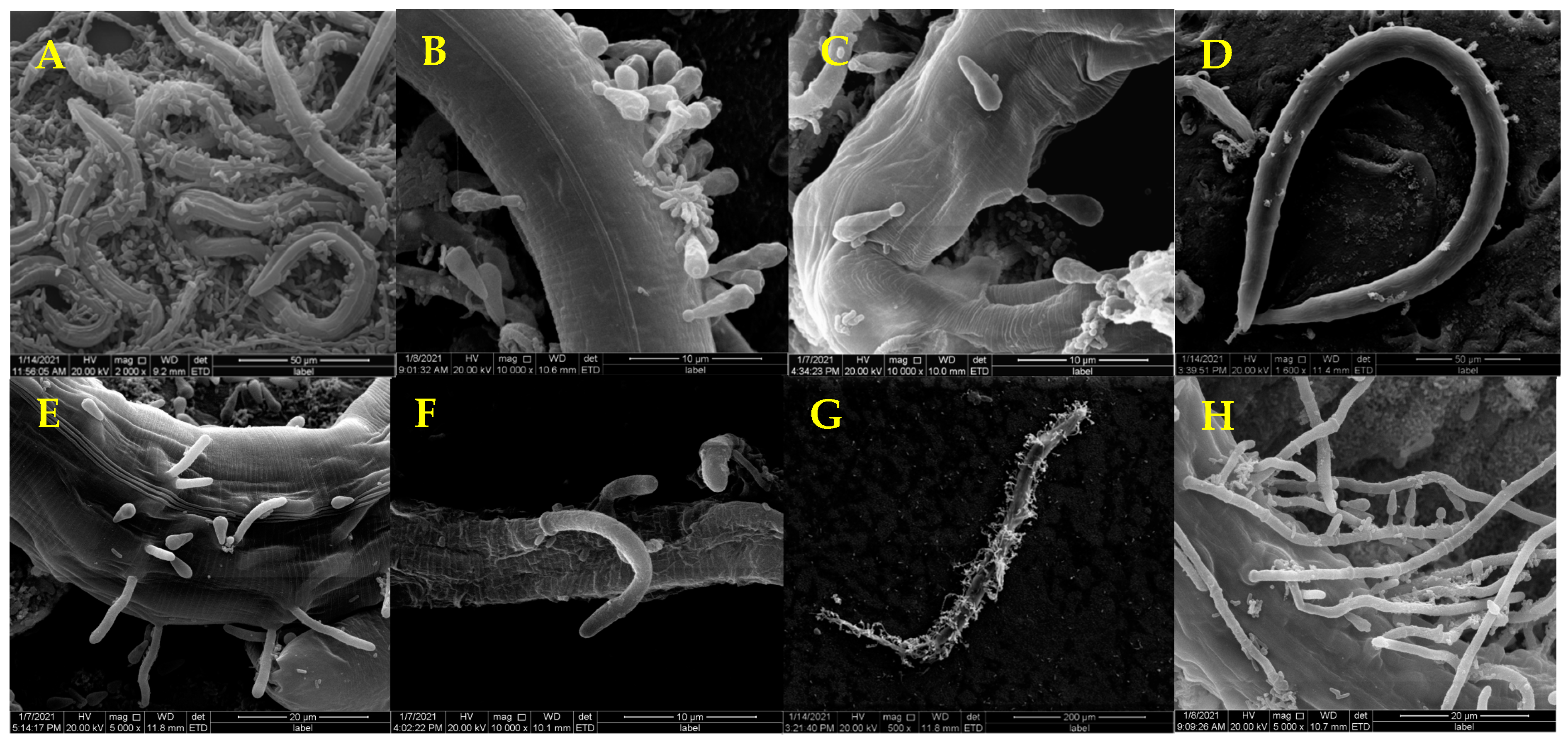
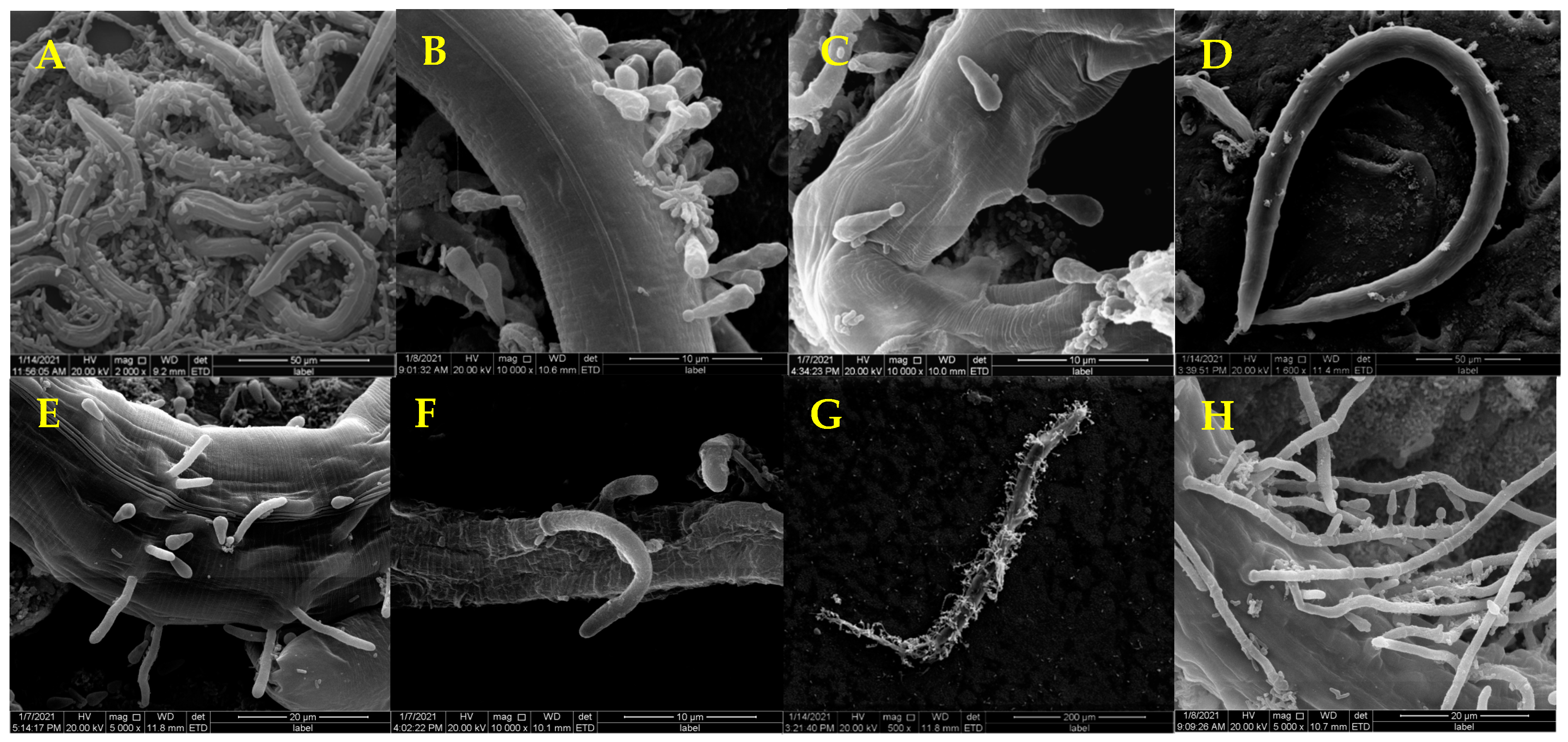
3.1. Pathogenicity of D. coniospora YMF1.01759 against Nematodes
3.2. Nematicidal Activity of Medium Screening Extracts
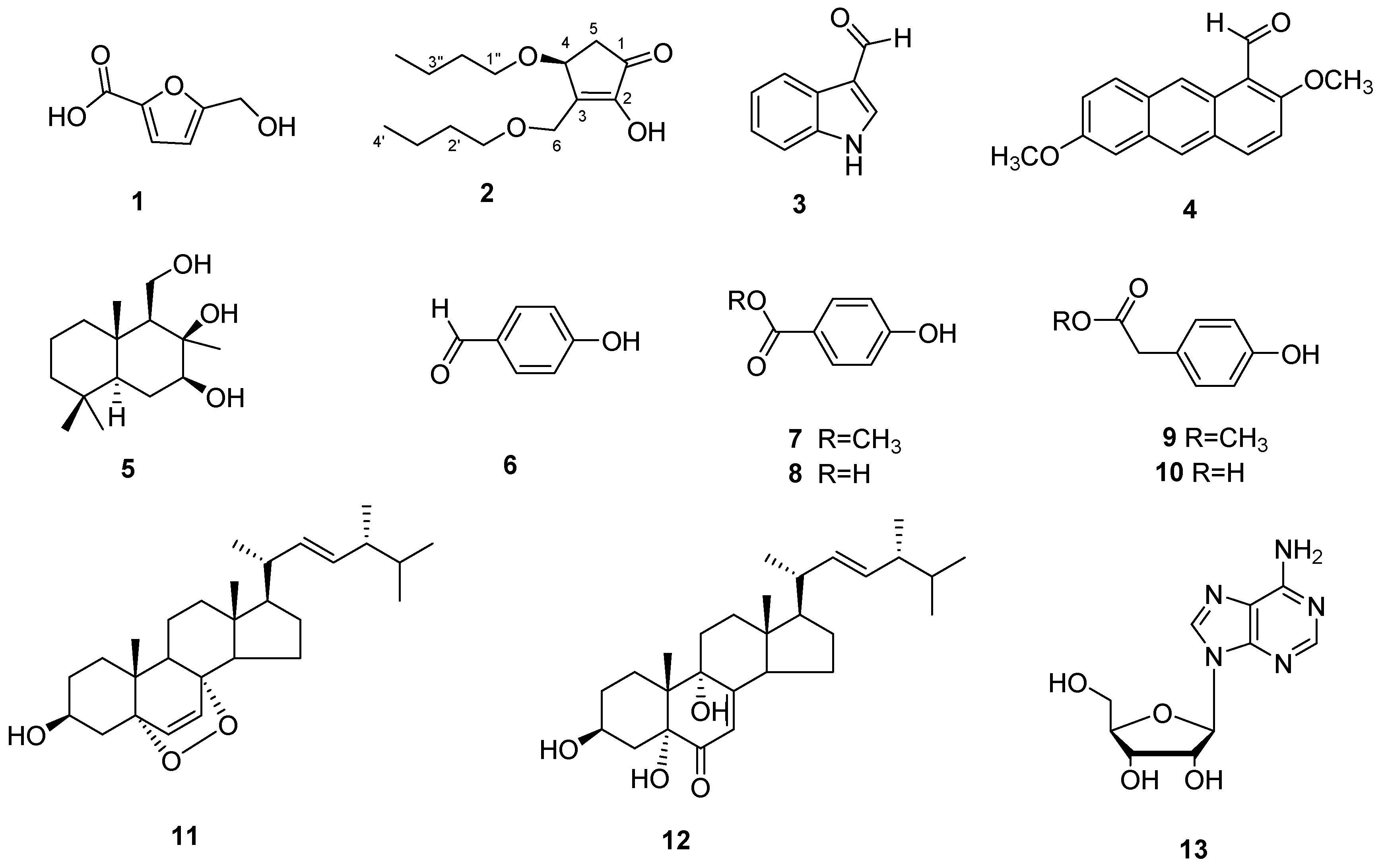
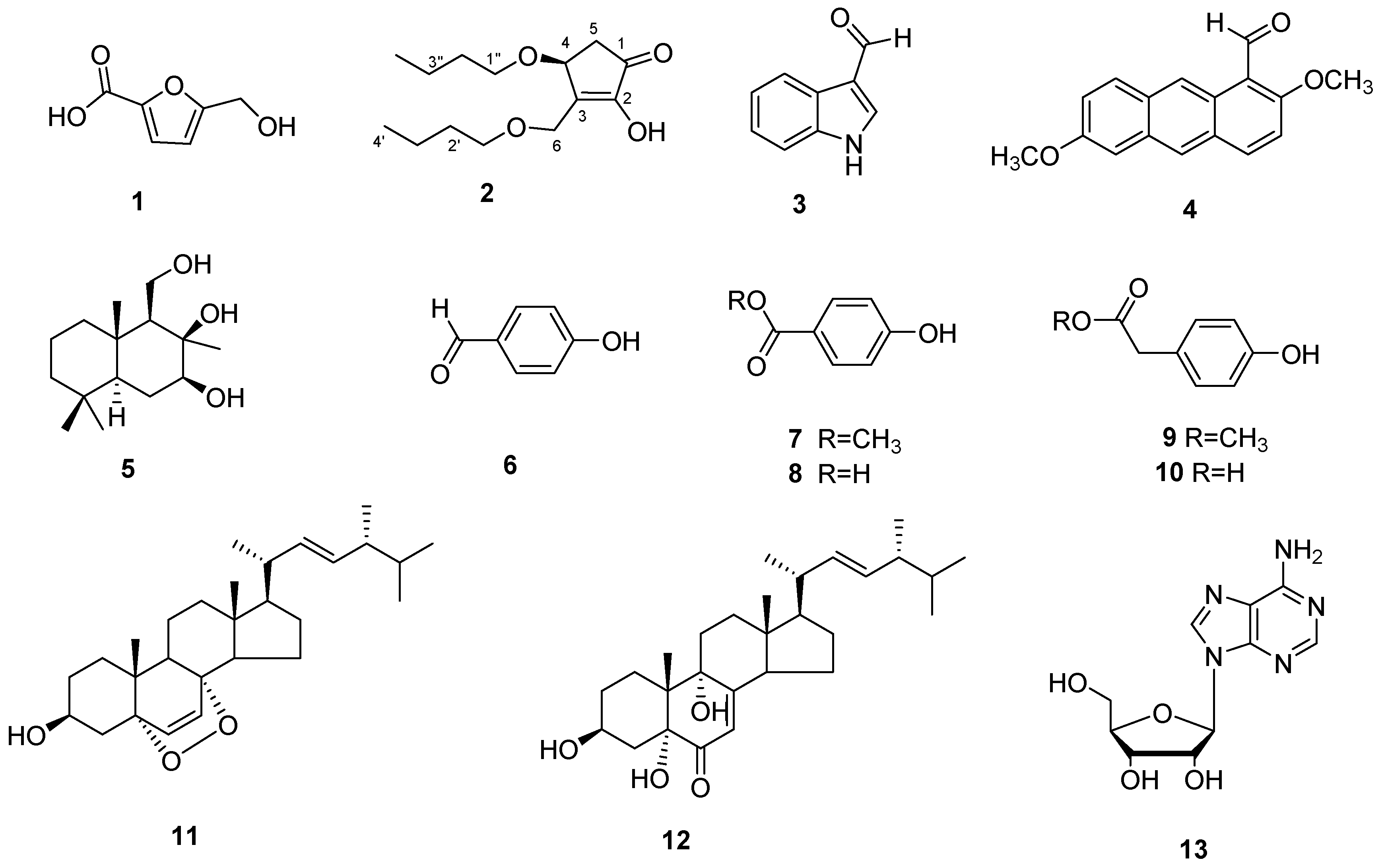
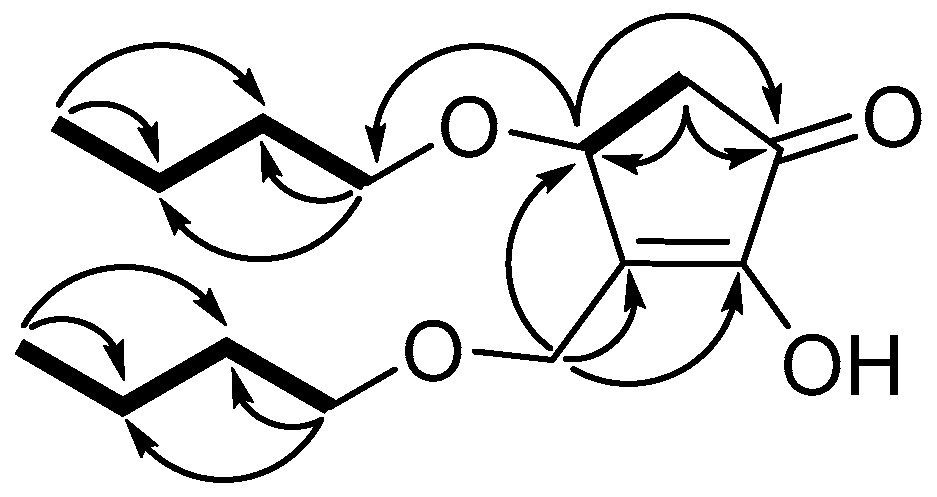
3.3. Structure Identification
3.4. Nematicidal Activity of Compounds and Their Inhibition of Egg Hatching
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Elling, A.A. Major emerging problems with minor Meloidogyne species. Phytopathology 2013, 103, 1092–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eloh, K.; Demurtas, M.; Mura, M.G.; Deplano, A.; Onnis, V.; Sasanelli, N.; Maxia, A.; Caboni, P. Potent nematicidal activity of maleimide derivatives on Meloidogyne incognita. J. Agric. Food Chem. 2016, 64, 4876–4881. [Google Scholar] [CrossRef]
- Hu, Y.; Zhang, W.P.; Zhang, P.; Ruan, W.B.; Zhu, X. Nematicidal activity of chaetoglobosin A poduced by Chaetomium globosum NK102 against Meloidogyne incognita. J. Agric. Food Chem. 2013, 61, 41–46. [Google Scholar] [CrossRef]
- Waldo, B.; Soto-Adames, F.; Crow, W. Nematicide effects on arthropods in bermudagrass. Fla. Entomol. 2020, 103, 458–464. [Google Scholar]
- Pino-Otín, M.R.; Val, J.; Ballestero, D.; Navarro, E.; Sánchez, E.; Mainar, A.M. Impact of Artemisia absinthium hydrolate extracts with nematicidal activity on non-target soil organisms of different trophic levels. Ecotoxicol. Environ. Saf. 2019, 180, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Copping, L.G.; Duke, S.O. Natural products that have been used commercially as crop protection agents. Pest Manag. Sci. 2007, 63, 524–554. [Google Scholar] [CrossRef]
- Dijksterhuis, J.; Harder, W.; Wyss, U.; Veenhuis, M. Colonization and digestion of nematodes by the endoparasitic nematophagous fungus Drechmeria coniospora. Mycol. Res. 1991, 95, 873–878. [Google Scholar] [CrossRef]
- Tunlid, A.; Jansson, H.B.; Nordbringhertz-Hertz, B. Fungal attachment to nematodes. Mycol. Res. 1992, 96, 401–412. [Google Scholar] [CrossRef]
- Liu, X.Z.; Xiang, M.C.; Che, Y.S. The living strategy of nematophagous fungi. Mycoscience 2009, 50, 20–25. [Google Scholar] [CrossRef]
- Jansson, H.B.; Jeyaprakash, A.; Zuckerman, B.M. Differential adhesion and infection of nematodes by the Endoparasitic fungus Meria coniospora (Deuteromycetes). Appl. Environ. Microb. 1985, 49, 552–555. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Zhou, Z.; Guo, Q.; Fokkens, L.; Miskei, M.; Pócsi, I.; Zhang, W.; Chen, M.; Wang, L.; Sun, Y.; et al. Insights into adaptations to a near-obligate nematode endoparasitic lifestyle from the finished genome of Drechmeria coniospora. Sci. Rep. 2016, 6, 23122. [Google Scholar] [CrossRef] [Green Version]
- Jansson, H.B. Adhesion of conidia of Drechmeria coniospora to Caenorhabditis elegans Wild type and mutants. J. Nematol. 1994, 26, 430–435. [Google Scholar]
- Khambay, B.P.S.; Bourne, J.M.; Cameron, S.; Kerry, B.R.; Zaki, M.J. Communication to the Editor—A nematicidal metabolite from Verticillium chlamydosporium. Pest Manag. Sci. 2000, 56, 1098–1099. [Google Scholar] [CrossRef]
- Niu, X.M.; Wang, Y.L.; Chu, Y.S.; Xue, H.X.; Li, N.; Wei, L.X.; Mo, M.H.; Zhang, K.Q. Nematodetoxic aurovertin-type metabolites from a root-knot nematode parasitic fungus Pochonia chlamydosporia. J. Agric. Food Chem. 2010, 58, 828–834. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.C.; Wang, Y.L.; Zhang, T.Y.; Chen, Z.J.; Yang, T.M.; Wu, Y.Y.; Sun, C.P.; Ma, X.C.; Zhang, Y.X. Indole diterpenoids from the endophytic fungus Drechmeria sp. as natural antimicrobial agents. Phytochemistry 2018, 148, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.C.; Luan, Z.L.; Liang, J.H.; Cheng, Z.B.; Sun, C.P.; Wang, Y.L.; Zhang, M.Y.; Zhang, T.Y.; Wang, Y.; Yang, T.M.; et al. Drechmerin H, a novel 1(2), 2(18)-diseco indole diterpenoid from the fungus Drechmeria sp. as a natural agonist of human pregnane X receptor. Bioorg. Chem. 2018, 79, 250–256. [Google Scholar] [CrossRef]
- Liang, J.H.; Huo, X.K.; Cheng, Z.B.; Sun, C.P.; Zhao, J.C.; Kang, X.H.; Zhang, T.Y.; Chen, Z.J.; Yang, T.M.; Wu, Y.Y.; et al. An indole diterpenoid isolated from the fungus Drechmeria sp. and its antimicrobial activity. Nat. Prod. Res. 2019, 33, 2770–2776. [Google Scholar] [CrossRef]
- Lebrigand, K.; He, L.D.; Thakur, N.; Arguel, M.J.; Polanowska, J.; Henrissat, B.; Record, E.; Magdelenat, G.; Barbe, V.; Raffaele, S.; et al. Comparative genomic analysis of Drechmeria coniospora reveals core and specific genetic requirements for fungal endoparasitism of nematodes. PLoS Genet. 2016, 12, e1006017. [Google Scholar] [CrossRef] [Green Version]
- Jang, J.Y.; Le, D.Q.; Choi, Y.H.; Choi, G.J.; Jang, K.S.; Cha, B.; Luu, N.H.; Kim, J.C. Nematicidal activities of 4-quinolone alkaloids isolated from the aerial part of Triumfetta grandidens against Meloidogyne incognita. J. Agric. Food Chem. 2014, 63, 68–74. [Google Scholar] [CrossRef]
- Huang, D.; Yu, C.; Shao, Z.; Cai, M.; Li, G.; Zheng, L.; Yu, Z.; Zhang, J. Identification and characterization of nematicidal volatile organic compounds from deep sea Virgibacillus dokdonensis MCCC 1A00493. Molecules 2020, 25, 744. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.Q. Genus and species of endoparasitic fungi of Meloidogyne incognita newly recorded in China. Acta Mycol. Sin. 1994, 13, 75–76. [Google Scholar]
- Boogert, P.; Dijksterhuis, J.; Velvis, H.; Veenhuis, M. Adhesive knob formation by conidia of the nematophagous fungus Drechmeria coniospora. Antonie Leeuwenhoek 1992, 61, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Jansson, H.B.; Friman, E. Infection-related surface proteins on conidia of the nematophagous fungus Drechmeria coniospora. Mycol. Res. 1999, 103, 249–256. [Google Scholar] [CrossRef]
- Mitsukura, K.; Sato, Y.; Yoshida, T.; Nagasawa, T. Oxidation of heterocyclic and aromatic aldehydes to the corresponding carboxylic acids by Acetobacter and Serratia strains. Biotechnol. Lett. 2004, 26, 1643–1648. [Google Scholar] [CrossRef]
- Soorukram, D.; Knochel, P. Formal enantioselective synthesis of (+)-estrone. Org. Lett. 2007, 9, 1021–1023. [Google Scholar] [CrossRef]
- Nakajima, E.; Nakano, H.; Yamada, K.; Shigemori, H.; Hasegawa, K. Isolation and identification of lateral bud growth inhibitor, indole-3-aldehyde, involved in apical dominance of pea seedlings. Phytochemistry 2002, 61, 863–865. [Google Scholar] [CrossRef]
- Bilger, C.; Demerseman, P.; Royer, R. Synthesis of new mutagenic compounds, nitro derivatives of anthrafurans. J. Heterocycl. Chem. 1985, 22, 735–739. [Google Scholar] [CrossRef]
- Panasenko, A.A.; Gorincioi, E.C.; Aricu, A.N.; Barcari, E.A.; Deleanu, K.; Vlad, P.F. H−1 and C−13 NMR spectra of some drimanic sesquiterpenoids. Russ. Chem. Bull. 2004, 53, 2700–2705. [Google Scholar] [CrossRef]
- Yue, J.M.; Chen, S.N.; Lin, Z.W.; Sun, H.D. Sterols from the fungus Lactarium volemus. Phytochemistry 2001, 56, 801–806. [Google Scholar] [CrossRef]
- Cai, H.H.; Liu, X.M.; Chen, Z.Y.; Liao, S.T.; Zou, Y.X. Isolation, purification and identification of nine chemical compounds from Flammulina velutipes fruiting bodies. Food Chem. 2013, 141, 2873–2879. [Google Scholar] [CrossRef]
- Kimura, Y.; Tania, S.; Hayashia, A.; Ohtania, K.; Fujiokab, S.; Kawanoa, T.; Shimada, A. Nematicidal activity of 5-hydroxymethyl-2-furoic acid against plant-parasitic nematodes. Z. Nat. C 2007, 62, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Liu, Y.J.; Fang, L.; Li, X.; Tan, N.H.; Zhang, K.Q. Coprinus comatus damages nematode cuticles mechanically with spiny balls and produces potent toxins to immobilize nematodes. Appl. Environ. Microb. 2007, 73, 3916–3923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, M.; Wada, K.; Munakata, K. New nematicidal metabolites from a fungus, Irpex lacteus. Agric. Biol. Chem. 1981, 45, 1527–1529. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | 1 H | 13 C | HMBC |
|---|---|---|---|
| 1 | - | 202.7, s | - |
| 2 | - | 153.6, s | - |
| 3 | - | 151.2, s | - |
| 4 | 4.65 (1 H, brd, J = 5.6 Hz) | 73.2, d | C-1, C-1″, C-3, C-6 |
| 5 | 2.24 (1 H, brd, J =18.4 Hz) | 41.8, t | C-1, C-3, C-4 |
| 2.70 (1 H, dd, J =18.4, 5.7 Hz) | C-1, C-3 | ||
| 6 | 4.28 (1 H, d, J = 13.3 Hz) | 55.6, t | C-2, C-3, C-4 |
| 4.55 (1 H, d, J = 13.3 Hz) | C-2, C-3, C-4 | ||
| 1′ | 4.20 (2 H, m) | 71.5, t | - |
| 2′ | 1.58 (1 H, m) | 33.2, t | - |
| 1.60 (1 H, m) | - | ||
| 3′ | 1.44 (2 H, m) | 20.4, t | C-4′ |
| 4′ | 0.93 (3 H, t, J = 7.4 Hz) | 14.1, q | C-2′, C-3′ |
| 1″ | 3.55 (2 H, m) | 70.7, t | C-2″ |
| 2″ | 1.44 (1 H, m) | 33.1, t | C-1″, C-4″ |
| 1.58 (1 H, m) | - | ||
| 3″ | 0.96 (1 H, m) | 19.9, t | - |
| 1.44 (1 H, m) | C-1″, C-4″ | ||
| 4″ | 0.94 (3 H, t, J = 7.4 Hz) | 14.2, q | C-2″, C-3″ |
| Concentration (µg mL−1) | 24 h | 48 h | 72 h |
|---|---|---|---|
| 400 | 90.10 ± 3.64 a | 99.51 ± 0.84 a | 100 a |
| 200 | 76.48 ± 3.86 a | 95.55 ± 1.7 a | 97.50 ± 0.91 a |
| 100 | 48.99 ± 2.05 a | 81.50 ± 2.46 a | 83.34 ± 1.67 a |
| 50 | 5.06 ± 0.87 a | 16.70 ± 0.82 a | 22.67 ± 2.37 a |
| 25 | 3.10 ± 0.49 a | 4.56 ± 0.59 a | 9.42 ± 0.69 a |
| Control (3% methanol) | 0.99 ± 0.08 | 2.03 ± 0.11 | 4.02 ± 0.14 |
| 400 µg mL−1 | 200 µg mL−1 | 100 µg mL−1 | 50 µg mL−1 | 25 µg mL−1 | |
|---|---|---|---|---|---|
| 24 h−48 h | −7.3 a | −19.5 a | −33.5 a | −3.8 b | −1.5 b |
| 48 h−72 h | −0.4 | −1.9 | −1.3 | −14.4 a | −4.2 c |
| Concentration (µg mL−1) | Hatched Worms per Egg Mass ± SD | ||
|---|---|---|---|
| 1 Day | 2 Days | 3 Days | |
| 200 | 5.83 ± 1.17 a | 8.17 ± 1.47 a | 11.17 ± 2.04 a |
| 100 | 12.00 ± 2.00 a | 18.50 ± 3.08 a | 24.00 ± 3.90 a |
| 50 | 13.50 ± 1.87 a | 35.17 ± 3.97 a | 44.00 ± 4.60 a |
| Control (3% methanol) | 37.33 ± 3.44 | 53.67 ± 3.33 | 63.33 ± 3.78 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wan, J.; Dai, Z.; Zhang, K.; Li, G.; Zhao, P. Pathogenicity and Metabolites of Endoparasitic Nematophagous Fungus Drechmeria coniospora YMF1.01759 against Nematodes. Microorganisms 2021, 9, 1735. https://doi.org/10.3390/microorganisms9081735
Wan J, Dai Z, Zhang K, Li G, Zhao P. Pathogenicity and Metabolites of Endoparasitic Nematophagous Fungus Drechmeria coniospora YMF1.01759 against Nematodes. Microorganisms. 2021; 9(8):1735. https://doi.org/10.3390/microorganisms9081735
Chicago/Turabian StyleWan, Juan, Zebao Dai, Keqin Zhang, Guohong Li, and Peiji Zhao. 2021. "Pathogenicity and Metabolites of Endoparasitic Nematophagous Fungus Drechmeria coniospora YMF1.01759 against Nematodes" Microorganisms 9, no. 8: 1735. https://doi.org/10.3390/microorganisms9081735
APA StyleWan, J., Dai, Z., Zhang, K., Li, G., & Zhao, P. (2021). Pathogenicity and Metabolites of Endoparasitic Nematophagous Fungus Drechmeria coniospora YMF1.01759 against Nematodes. Microorganisms, 9(8), 1735. https://doi.org/10.3390/microorganisms9081735