
Molecular Characterization of Staphylococcus aureus Isolated from Raw Milk Samples of Dairy Cows in Manhiça District, Southern Mozambique

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
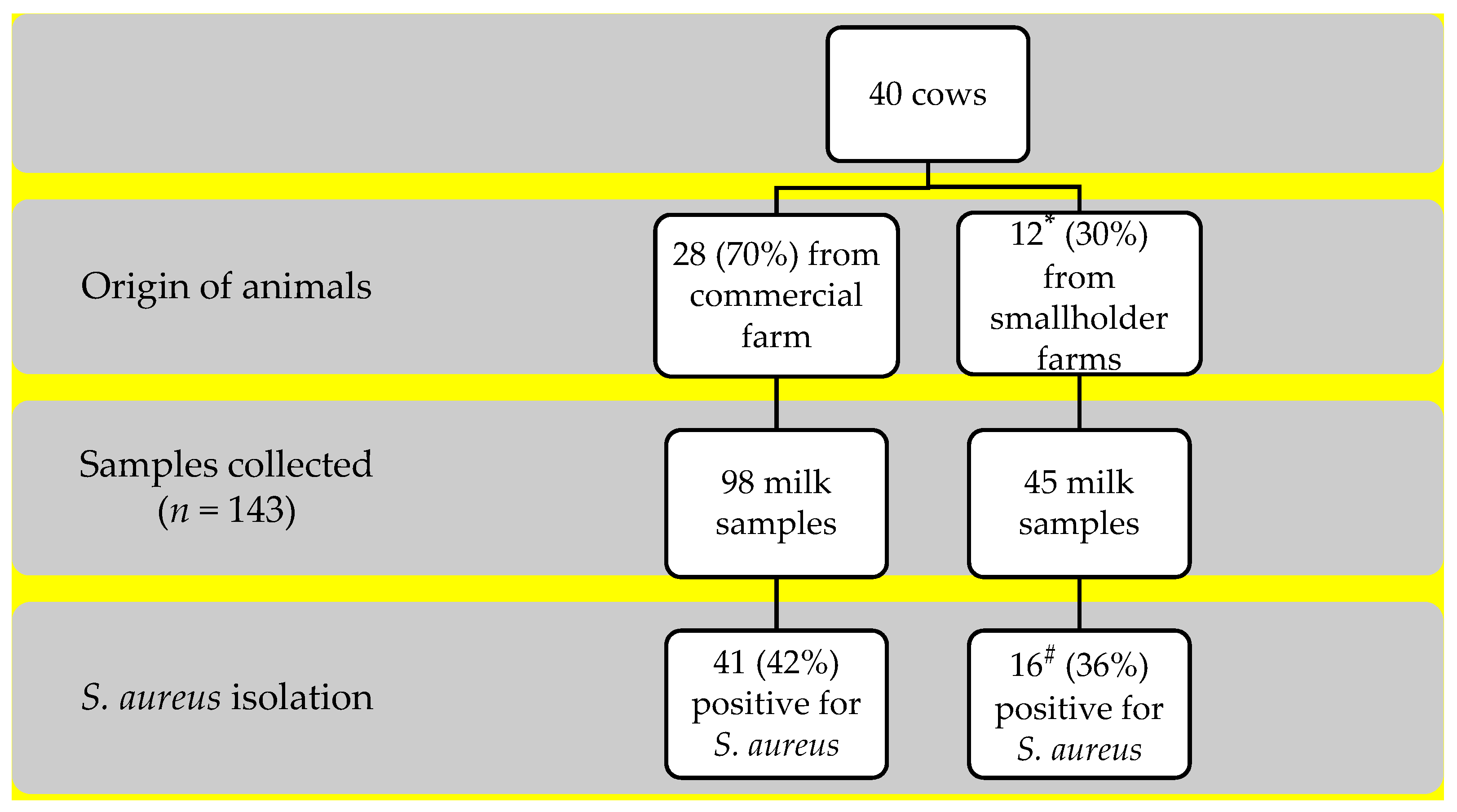
2.1. Study Design
2.2. Specimen Collection
2.3. Isolation and Identification of Staphylococcus aureus
2.4. Preparation of Bacterial DNA and Molecular Confirmation of S. aureus
2.5. Antimicrobial Susceptibility Test
2.6. Screening of Resistance Determinants
2.7. Detection of Virulence Genes
2.8. Staphylococcal Protein A (spa) Typing
2.9. Multilocus Sequence Typing (MLST)
2.10. Data Analysis
3. Results
3.1. Frequency of S. aureus Isolation
3.2. Antibiotic Susceptibility
3.3. Resistance Determinants
3.3.1. Resistance Determinants on Resistant Isolates
3.3.2. Resistance Determinants on Susceptible Isolates
3.4. Molecular Typing
3.5. PVL and Staphylococcal Enterotoxins (SEs) Gene Detection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hennekinne, J.-A.; De Buyser, M.-L.; Dragacci, S. Staphylococcus aureus and Its Food Poisoning Toxins: Characterization and Outbreak Investigation. FEMS Microbiol. Rev. 2012, 36, 815–836. [Google Scholar] [CrossRef] [PubMed]
- Claeys, W.L.; Cardoen, S.; Daube, G.; De Block, J.; Dewettinck, K.; Dierick, K.; De Zutter, L.; Huyghebaert, A.; Imberechts, H.; Thiange, P.; et al. Raw or Heated Cow Milk Consumption: Review of Risks and Benefits. Food Control. 2013, 31, 251–262. [Google Scholar] [CrossRef]
- Le Loir, Y.; Baron, F.; Gautier, M. Staphylococcus aureus and Food Poisoning. Genet. Mol. Res. 2003, 2, 63–76. [Google Scholar]
- Argudín, M.Á.; Mendoza, M.C.; Rodicio, M.R. Food Poisoning and Staphylococcus aureus Enterotoxins. Toxins 2010, 2, 1751–1773. [Google Scholar] [CrossRef]
- Lindsay, J.A.; Holden, M.T.G. Staphylococcus aureus: Superbug, Super Genome? Trends Microbiol. 2004, 12, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, J.R. Livestock-Associated Staphylococcus aureus: Origin, Evolution and Public Health Threat. Trends Microbiol. 2012, 20, 192–198. [Google Scholar] [CrossRef]
- Lundberg, Å.; Aspán, A.; Nyman, A.; Unnerstad, H.; Waller, K. Associations between Bacterial Genotype and Outcome of Bovine Clinical Staphylococcus aureus Mastitis. Acta Vet. Scand. 2014, 56, 2. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, P.; Papadopoulos, T.; Angelidis, A.S.; Kotzamanidis, C.; Zdragas, A.; Papa, A.; Filioussis, G.; Sergelidis, D. Prevalence, Antimicrobial Susceptibility and Characterization of Staphylococcus aureus and Methicillin-Resistant Staphylococcus aureus Isolated from Dairy Industries in North-Central and North-Eastern Greece. Int. J. Food Microbiol. 2019, 291, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Getahun, K.; Kelay, B.; Bekana, M.; Lobago, F. Bovine Mastitis and Antibiotic Resistance Patterns in Selalle Smallholder Dairy Farms, Central Ethiopia. Trop Anim. Health Prod. 2008, 40, 261–268. [Google Scholar] [CrossRef]
- Haftu, R.; Taddele, H.; Gugsa, G.; Kalayou, S. Prevalence, Bacterial Causes, and Antimicrobial Susceptibility Profile of Mastitis Isolates from Cows in Large-Scale Dairy Farms of Northern Ethiopia. Trop Anim. Health Prod. 2012, 44, 1765–1771. [Google Scholar] [CrossRef]
- Lindsay, J.A.; Holden, M.T.G. Understanding the Rise of the Superbug: Investigation of the Evolution and Genomic Variation of Staphylococcus aureus. Funct Integr. Genom. 2006, 6, 186–201. [Google Scholar] [CrossRef]
- Jackson, C.R.; Davis, J.A.; Barrett, J.B. Prevalence and Characterization of Methicillin-Resistant Staphylococcus aureus Isolates from Retail Meat and Humans in Georgia. J. Clin. Microbiol. 2013, 51, 1199–1207. [Google Scholar] [CrossRef] [PubMed]
- Zadoks, R.N.; Middleton, J.R.; McDougall, S.; Katholm, J.; Schukken, Y.H. Molecular Epidemiology of Mastitis Pathogens of Dairy Cattle and Comparative Relevance to Humans. J. Mammary Gland Biol. Neoplasia 2011, 16, 357–372. [Google Scholar] [CrossRef]
- Mekonnen, S.A.; Lam, T.J.G.M.; Hoekstra, J.; Rutten, V.P.M.G.; Tessema, T.S.; Broens, E.M.; Riesebos, A.E.; Spaninks, M.P.; Koop, G. Characterization of Staphylococcus aureus Isolated from Milk Samples of Dairy Cows in Small Holder Farms of North-Western Ethiopia. BMC Vet. Res. 2018, 14, 246. [Google Scholar] [CrossRef] [PubMed]
- Eyasu, T.; Tesfu, K.; Daniel, A.; Haile, A.; Thomas, S.; Pamela, R.F.A.; Wondwossen, G. Phenotypic and Genotypic Characterization of Staphylococcus aureus Isolates Recovered from Bovine Milk in Central Highlands of Ethiopia. Afr. J. Microbiol. Res. 2015, 9, 2209–2217. [Google Scholar] [CrossRef][Green Version]
- Schmidt, T.; Kock, M.M.; Ehlers, M.M. Molecular Characterization of Staphylococcus aureus Isolated from Bovine Mastitis and Close Human Contacts in South African Dairy Herds: Genetic Diversity and Inter-Species Host Transmission. Front. Microbiol. 2017, 8, 511. [Google Scholar] [CrossRef]
- Ruffing, U.; Alabi, A.; Kazimoto, T.; Vubil, D.C.; Akulenko, R.; Abdulla, S.; Alonso, P.; Bischoff, M.; Germann, A.; Grobusch, M.P.; et al. Community-Associated Staphylococcus aureus from Sub-Saharan Africa and Germany: A Cross-Sectional Geographic Correlation Study. Sci. Rep. 2017, 7, 154. [Google Scholar] [CrossRef] [PubMed]
- Vubil, D.; Garrine, M.; Ruffing, U.; Acácio, S.; Sigaúque, B.; Alonso, P.L.; von Müller, L.; Herrmann, M.; Mandomando, I. Molecular Characterization of Community Acquired Staphylococcus aureus Bacteremia in Young Children in Southern Mozambique, 2001–2009. Front. Microbiol. 2017, 8, 730. [Google Scholar] [CrossRef]
- Ceccarelli, D.; Mondlane, J.; Sale, M.; Salvia, A.M.; Folgosa, E.; Cappuccinelli, P.; Colombo, M.M. Sporadic Methicillin Resistance in Community Acquired Staphylococcus aureus in Mozambique. New Microbiol. 2005, 28, 327–336. [Google Scholar]
- Van der Meeren, B.T.; Millard, P.S.; Scacchetti, M.; Hermans, M.H.; Hilbink, M.; Concelho, T.B.; Ferro, J.J.; Wever, P.C. Emergence of Methicillin Resistance and Panton-Valentine Leukocidin Positivity in Hospital- and Community-Acquired Staphylococcus aureus Infections in Beira, Mozambique. Trop. Med. Int. Health 2014, 19, 169–176. [Google Scholar] [CrossRef]
- Mandomando, I.; Espasa, M.; Nhampossa, T.; Roca, A.; Sigaúque, B.; Menéndez, C.; Macete, E.; Machevo, S.; Alonso, P.L.; Quintò, L.; et al. Antimicrobial Drug Resistance Trends of Bacteremia Isolates in a Rural Hospital in Southern Mozambique. Am. J. Trop. Med. Hyg. 2010, 83, 152–157. [Google Scholar] [CrossRef]
- Sigaúque, B.; Roca, A.; Mandomando, I.; Morais, L.; Quintó, L.; Sacarlal, J.; Macete, E.; Nhamposa, T.; Machevo, S.; Aide, P.; et al. Community-Acquired Bacteremia among Children Admitted to a Rural Hospital in Mozambique. Pediatric Infect. Dis. J. 2009, 28, 108–113. [Google Scholar] [CrossRef]
- Alonso, P.; Saúte, F.; Aponte, J.; Gómez-Olivé, F.; Nhacolo, A.; Thomson, R.; Macete, E.; Abacassamo, F.; Ventura, P.; Bosch, X.; et al. Manhiça DSS, Mozambique. In Population and Health in Developing Countries Volume 1. Population, Health, and Survival at INDEPTH Sites, 1st ed.; INDEPTH, Ed.; International Development Research Centre (IDRC): Ottawa, ON, Canada, 2002; pp. 189–195. [Google Scholar]
- Alexopoulou, K.; Foka, A.; Petinaki, E.; Jelastopulu, E.; Dimitracopoulos, G.; Spiliopoulou, I. Comparison of Two Commercial Methods with PCR Restriction Fragment Length Polymorphism of the Tuf Gene in the Identification of Coagulase-Negative Staphylococci. Lett. Appl. Microbiol. 2006, 43, 450–454. [Google Scholar] [CrossRef]
- Brakstad, O.G.; Aasbakk, K.; Maeland, J.A. Detection of Staphylococcus aureus by Polymerase Chain Reaction Amplification of the Nuc Gene. J. Clin. Microbiol. 1992, 30, 1654–1660. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; Supplement M100S; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Lin, X.; Jiang, T.; Peng, Z.; Xu, J.; Yi, L.; Li, F.; Fanning, S.; Baloch, Z. Prevalence and Characterization of Staphylococcus aureus Cultured From Raw Milk Taken From Dairy Cows With Mastitis in Beijing, China. Front. Microbiol. 2018, 9, 1123. [Google Scholar] [CrossRef]
- Lina, G.; Piemont, Y.; Godail-Gamot, F.; Bes, M.; Peter, M.-O.; Gauduchon, V.; Vandenesch, F.; Etienne, J. Involvement of Panton-Valentine Leukocidin--Producing Staphylococcus aureus in Primary Skin Infections and Pneumonia. Clin. Infect. Dis. 1999, 29, 1128–1132. [Google Scholar] [CrossRef]
- Aires-de-Sousa, M.; Boye, K.; de Lencastre, H.; Deplano, A.; Enright, M.C.; Etienne, J.; Friedrich, A.; Harmsen, D.; Holmes, A.; Huijsdens, X.W.; et al. High Interlaboratory Reproducibility of DNA Sequence-Based Typing of Bacteria in a Multicenter Study. J. Clin. Microbiol. 2006, 44, 619–621. [Google Scholar] [CrossRef] [PubMed]
- Enright, M.C.; Day, N.P.; Davies, C.E.; Peacock, S.J.; Spratt, B.G. Multilocus Sequence Typing for Characterization of Methicillin-Resistant and Methicillin-Susceptible Clones of Staphylococcus aureus. J. Clin. Microbiol. 2000, 38, 1008–1015. [Google Scholar] [CrossRef]
- Hunter, P.R.; Gaston, M.A. Numerical Index of the Discriminatory Ability of Typing Systems: An Application of Simpson’s Index of Diversity. J. Clin. Microbiol. 1988, 26, 2465–2466. [Google Scholar] [CrossRef] [PubMed]
- Abera, M.; Habte, T.; Aragaw, K.; Asmare, K.; Sheferaw, D. Major Causes of Mastitis and Associated Risk Factors in Smallholder Dairy Farms in and around Hawassa, Southern Ethiopia. Trop Anim. Health Prod. 2012, 44, 1175–1179. [Google Scholar] [CrossRef] [PubMed]
- Katsande, S.; Matope, G.; Ndengu, M.; Pfukenyi, D.M. Prevalence of Mastitis in Dairy Cows from Smallholder Farms in Zimbabwe. Onderstepoort J. Vet. Res. 2013, 80, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Olde Riekerink, R.G.M.; Barkema, H.W.; Kelton, D.F.; Scholl, D.T. Incidence Rate of Clinical Mastitis on Canadian Dairy Farms. J. Dairy Sci. 2008, 91, 1366–1377. [Google Scholar] [CrossRef]
- Ericsson Unnerstad, H.; Lindberg, A.; Persson Waller, K.; Ekman, T.; Artursson, K.; Nilsson-Öst, M.; Bengtsson, B. Microbial Aetiology of Acute Clinical Mastitis and Agent-Specific Risk Factors. Vet. Microbiol. 2009, 137, 90–97. [Google Scholar] [CrossRef]
- Kadlec, K.; Entorf, M.; Peters, T. Occurrence and Characteristics of Livestock-Associated Methicillin-Resistant Staphylococcus aureus in Quarter Milk Samples From Dairy Cows in Germany. Front. Microbiol. 2019, 10, 1–6. [Google Scholar] [CrossRef]
- Yang, F.-L.; Li, X.-S.; He, B.-X.; Du, Y.-L.; Li, G.-H.; Yang, B.-B.; Qin-Hua, H. Bovine Mastitis in Subtropical Dairy Farms, 2005-2009. J. Anim. Vet. Adv. 2011, 10, 68–72. [Google Scholar] [CrossRef][Green Version]
- Sudhanthiramani, S.; Swetha, C.S.; Bharathy, S. Prevalence of Antibiotic Resistant Staphylococcus aureus from Raw Milk Samples Collected from the Local Vendors in the Region of Tirupathi, India. Vet. World 2015, 8, 478–481. [Google Scholar] [CrossRef]
- Garrine, M.; Costa, S.S.; Mandomando, I.; Couto, I. Antimicrobial Resistance and Molecular Typing of Staphylococcus aureus Causing Bacteraemia in Children Aged Less than 5 Years in Southern Mozambique, 2001–2019; Centro de Investigação Em Saúde de Manhiça (CISM): Maputo, Mozambique, 2021. [Google Scholar]
- Schmidt, T.; Kock, M.M.; Ehlers, M.M. Diversity and Antimicrobial Susceptibility Profiling of Staphylococci Isolated from Bovine Mastitis Cases and Close Human Contacts. J. Dairy Sci. 2015, 98, 6256–6269. [Google Scholar] [CrossRef] [PubMed]
- Martini, C.L.; Lange, C.C.; Brito, M.A.; Ribeiro, J.B.; Mendonça, L.C.; Vaz, E.K. Characterisation of Penicillin and Tetracycline Resistance in Staphylococcus aureus Isolated from Bovine Milk Samples in Minas Gerais, Brazil. J. Dairy Res. 2017, 84, 202–205. [Google Scholar] [CrossRef]
- Oniciuc, E.-A.; Nicolau, A.I.; Hernández, M.; Rodríguez-Lázaro, D. Presence of Methicillin-Resistant Staphylococcus aureus in the Food Chain. Trends Food Sci. Technol. 2017, 61, 49–59. [Google Scholar] [CrossRef]
- Capurro, A.; Aspán, A.; Ericsson Unnerstad, H.; Persson Waller, K.; Artursson, K. Identification of Potential Sources of Staphylococcus aureus in Herds with Mastitis Problems. J. Dairy Sci. 2010, 93, 180–191. [Google Scholar] [CrossRef] [PubMed]
- Sommerhäuser, J.; Kloppert, B.; Wolter, W.; Zschöck, M.; Sobiraj, A.; Failing, K. The Epidemiology of Staphylococcus aureus Infections from Subclinical Mastitis in Dairy Cows during a Control Programme. Vet. Microbiol. 2003, 96, 91–102. [Google Scholar] [CrossRef]
- Gurjar, A.; Gioia, G.; Schukken, Y.; Welcome, F.; Zadoks, R.; Moroni, P. Molecular Diagnostics Applied to Mastitis Problems on Dairy Farms. Vet. Clin. North. Am. Food Anim. Pract. 2012, 28, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Haveri, M.; Hovinen, M.; Roslöf, A.; Pyörälä, S. Molecular Types and Genetic Profiles of Staphylococcus aureus Strains Isolated from Bovine Intramammary Infections and Extramammary Sites. J. Clin. Microbiol. 2008, 46, 3728–3735. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zadoks, R.N.; van Leeuwen, W.B.; Kreft, D.; Fox, L.K.; Barkema, H.W.; Schukken, Y.H.; van Belkum, A. Comparison of Staphylococcus aureus Isolates from Bovine and Human Skin, Milking Equipment, and Bovine Milk by Phage Typing, Pulsed-Field Gel Electrophoresis, and Binary Typing. J. Clin. Microbiol. 2002, 40, 3894–3902. [Google Scholar] [CrossRef]
- Basanisi, M.G.; La Bella, G.; Nobili, G.; Franconieri, I.; La Salandra, G. Genotyping of Methicillin-Resistant Staphylococcus aureus (MRSA) Isolated from Milk and Dairy Products in South Italy. Food Microbiol. 2017, 62, 141–146. [Google Scholar] [CrossRef]
- Dan, M.; Yehui, W.; Qingling, M.; Jun, Q.; Xingxing, Z.; Shuai, M.; Kuojun, C.; Jinsheng, Z.; Zibing, C.; Zaichao, Z.; et al. Antimicrobial Resistance, Virulence Gene Profile and Molecular Typing of Staphylococcus aureus Isolates from Dairy Cows in Xinjiang Province, Northwest China. J. Glob. Antimicrob. Resist. 2019, 16, 98–104. [Google Scholar] [CrossRef]
- de Alves, M.F.N.F.; Penna, B.; Pereira, R.F.A.; Geraldo, R.B.; Folly, E.; Castro, H.C.; Aguiar-Alves, F. First Report of Meticillin-Resistant Staphylococcus aureus Harboring MecC Gene in Milk Samples from Cows with Mastitis in Southeastern Brazil. Braz. J. Microbiol. 2020, 51, 2175–2179. [Google Scholar] [CrossRef]
- Ruimy, R.; Maiga, A.; Armand-Lefevre, L.; Maiga, I.; Diallo, A.; Koumaré, A.K.; Ouattara, K.; Soumaré, S.; Gaillard, K.; Lucet, J.-C.; et al. The Carriage Population of Staphylococcus aureus from Mali Is Composed of a Combination of Pandemic Clones and the Divergent Panton-Valentine Leukocidin-Positive Genotype ST152. J. Bacteriol. (JB) 2008, 190, 3962–3968. [Google Scholar] [CrossRef]
- Koop, G.; Vrieling, M.; Storisteanu, D.M.L.; Lok, L.S.C.; Monie, T.; van Wigcheren, G.; Raisen, C.; Ba, X.; Gleadall, N.; Hadjirin, N.; et al. Identification of LukPQ, a Novel, Equid-Adapted Leukocidin of Staphylococcus aureus. Sci. Rep. 2017, 7, 40660. [Google Scholar] [CrossRef]


{kind=link}
| Antibiotics | Commercial Farm n = 41 (%) | Smallholder Farms n = 17 (%) | p | Total n = 58 (%) |
|---|---|---|---|---|
| Penicillin | 12 (29) | 13 (76) | 0.001 | 25 (43) |
| Tetracycline | 3 (7) | 6 (35) | 0.014 | 9 (16) |
| Erythromycin/Clindamycin 1 | 1 (2) | 1 (6) | 0.504 | 2 (3) |
| Co-trimoxazole | 0 | 1 (6) | NA | 1 (2) |
| N° Isolates Per Sector | ||||||
|---|---|---|---|---|---|---|
| Spa type (n) | MLST (n) | PVL Gene (n) | Smallholder Farms (n) | Commercial Farm (n) | Resistance Pattern (n) | Resistance Determinants (n) |
| t5388 (44) | ST1/CC1 (44) | 0 | 8 | 36 | PEN (13); PEN-TET (1); TET (1) | blaZ (1); tetK (2); blaZ-tetK (1), mecA-tetK (1) |
| t1299 (7) | ST152 (7) | 7 | 4 | 3 | PEN (2); TET (3); PEN-TET (1), PEN-TET-ERY-CD, (1) | blaZ (1); blaZ-ermC-tetK-tetL (1); blaZ-tetK-tetL (1); tetK-tetL (2); tetK-tetM-tetL (1) |
| t1476 (3) | ST8/CC8 (3) | 0 | 3 | 0 | PEN (1); PEN-TET (1); PEN-TET-SXT (1) | blaZ (1); blaZ-tetK-tetL (1); blaZ, tetK, dfrG (1) |
| t002 (2) | ST5/CC5 (2) | 0 | 0 | 2 | PEN (2) | blaZ (2) |
| t331 (1) | ST508/CC45 (1) | 0 | 1 | 0 | PEN | blaZ-mecA-tetK |
| t355 (1) | ST152 (1) | 1 | 1 | 0 | PEN-ERY-CD | blaZ-msrA-ermC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nhatsave, N.; Garrine, M.; Messa, A., Jr.; Massinga, A.J.; Cossa, A.; Vaz, R.; Ombi, A.; Zimba, T.F.; Alfredo, H.; Mandomando, I.; et al. Molecular Characterization of Staphylococcus aureus Isolated from Raw Milk Samples of Dairy Cows in Manhiça District, Southern Mozambique. Microorganisms 2021, 9, 1684. https://doi.org/10.3390/microorganisms9081684
Nhatsave N, Garrine M, Messa A Jr., Massinga AJ, Cossa A, Vaz R, Ombi A, Zimba TF, Alfredo H, Mandomando I, et al. Molecular Characterization of Staphylococcus aureus Isolated from Raw Milk Samples of Dairy Cows in Manhiça District, Southern Mozambique. Microorganisms. 2021; 9(8):1684. https://doi.org/10.3390/microorganisms9081684
Chicago/Turabian StyleNhatsave, Nilsa, Marcelino Garrine, Augusto Messa, Jr., Arsénia J. Massinga, Anélsio Cossa, Raúl Vaz, Angelina Ombi, Tomás F. Zimba, Hélder Alfredo, Inácio Mandomando, and et al. 2021. "Molecular Characterization of Staphylococcus aureus Isolated from Raw Milk Samples of Dairy Cows in Manhiça District, Southern Mozambique" Microorganisms 9, no. 8: 1684. https://doi.org/10.3390/microorganisms9081684
APA StyleNhatsave, N., Garrine, M., Messa, A., Jr., Massinga, A. J., Cossa, A., Vaz, R., Ombi, A., Zimba, T. F., Alfredo, H., Mandomando, I., & Tchamo, C. (2021). Molecular Characterization of Staphylococcus aureus Isolated from Raw Milk Samples of Dairy Cows in Manhiça District, Southern Mozambique. Microorganisms, 9(8), 1684. https://doi.org/10.3390/microorganisms9081684