
Assessment of Phycocyanin Extraction from Cyanidium caldarium by Spark Discharges, Compared to Freeze-Thaw Cycles, Sonication, and Pulsed Electric Fields

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Cultivation and Harvesting
2.2. Disintegration by Freeze-Thaw-Cycles
2.3. Disintegration by Sonication
2.4. Disintegration by Pulsed Electric Fields (PEF)
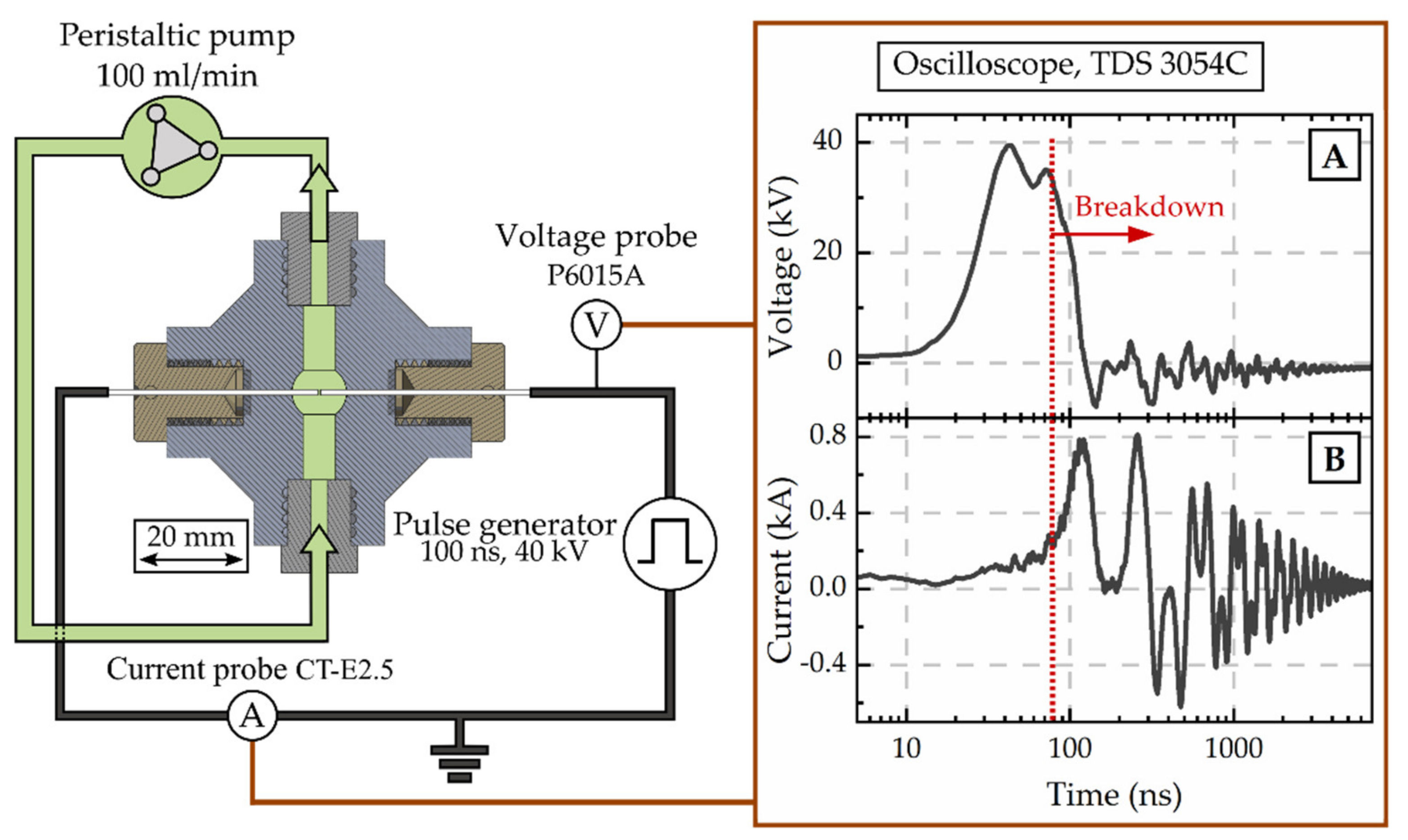
2.5. Disintegration by Spark Discharges
2.6. Photometric Determination of Phycocyanin Content and Purity of Raw Extract
2.7. Protein Precipitation and Desalination
2.8. Evaluation of the Purified Extract by Fluorescence-Spectroscopy
2.9. Evaluation of Purified Extract by Absorption Spectroscopy
2.10. NanoLC-MS/MS Data Acquisition and Protein Identification
3. Results
3.1. Freeze-Thaw Cycles
3.2. Sonication
3.2.1. Continuous Sonication
3.2.2. Intermittent Sonication
3.3. PEF Exposure
3.4. Spark Discharge Treatment
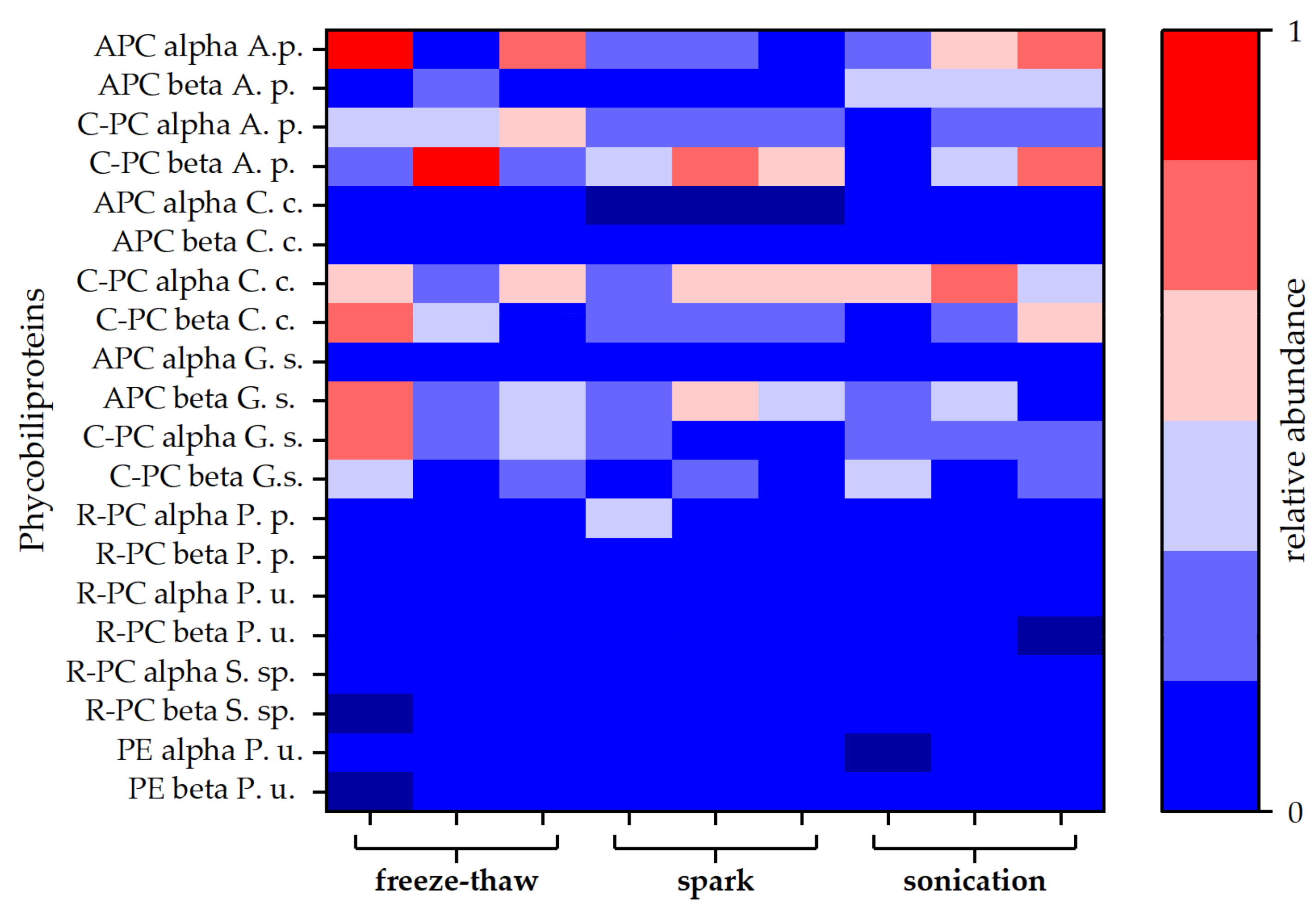
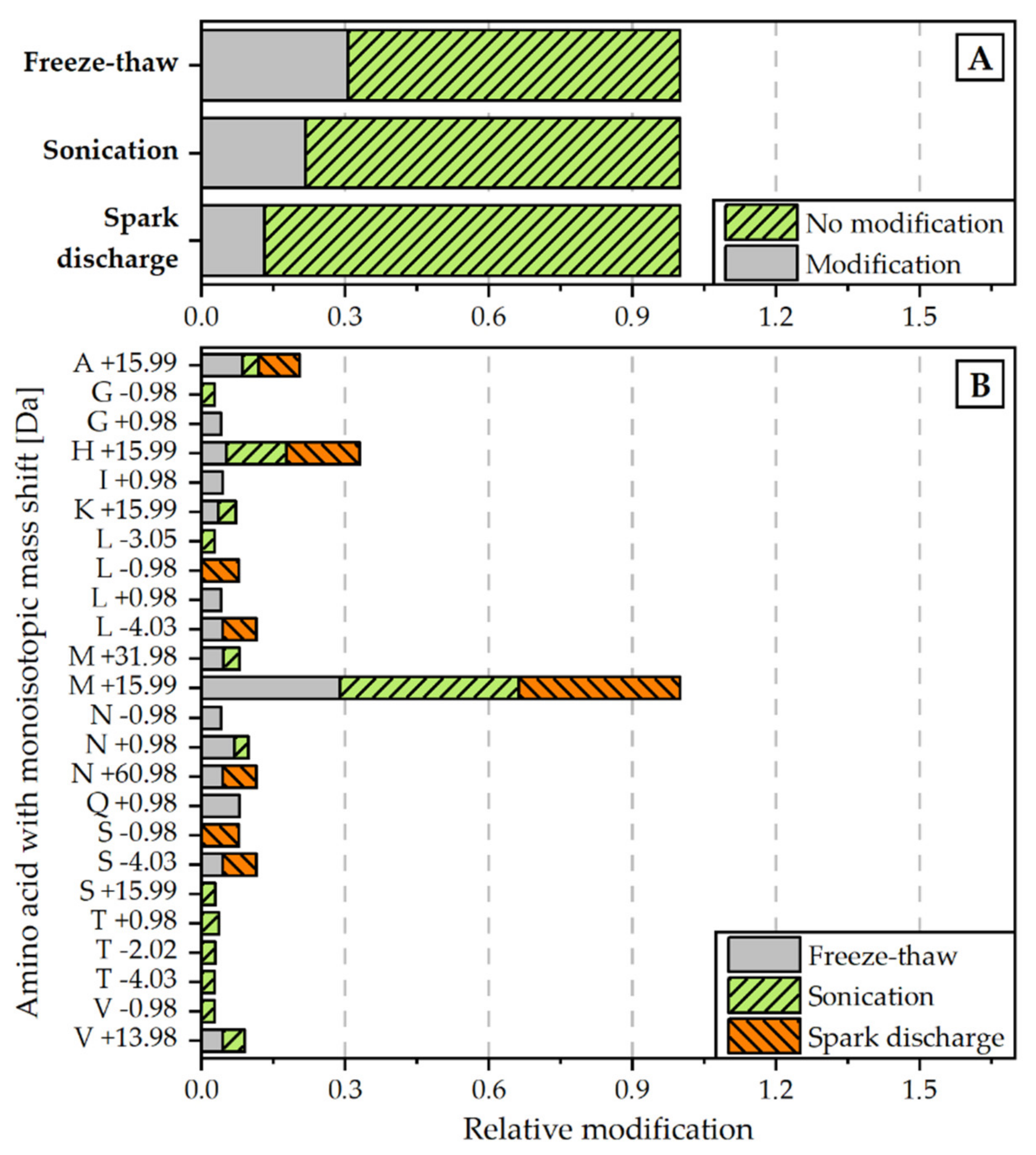
3.5. Protein Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kumar, D.; Dhar, D.W.; Pabbi, S.; Kumar, N.; Walia, S. Extraction and purification of C-phycocyanin from Spirulina platensis (CCC540). Indian J. Plant Physiol. 2014, 19, 184–188. [Google Scholar] [CrossRef]
- Feketea, G.; Tsabouri, S. Common food colorants and allergic reactions in children: Myth or reality? Food Chem. 2017, 230, 578–588. [Google Scholar] [CrossRef]
- Costa, D.; Mendonça, M.; Lopes, M.; Fernandes, A.L.; Nunes, S.; Müller, S. Patent blue V dye anaphylaxis: A case report and literature review. Rev. Bras. Anestesiol. 2020, 70, 662–666. [Google Scholar] [CrossRef]
- Ku, C.S.; Yang, Y.; Park, Y.; Lee, J. Health benefits of blue-green algae: Prevention of cardiovascular disease and nonalcoholic fatty liver disease. J. Med. Food 2013, 16, 103–111. [Google Scholar] [CrossRef]
- Remirez, D.; Ledón, N.; González, R. Role of histamine in the inhibitory effects of phycocyanin in experimental models of allergic inflammatory response. Mediat. Inflamm. 2002, 11, 109485. [Google Scholar] [CrossRef]
- Romay, C.; Gonzalez, R.; Ledon, N.; Remirez, D.; Rimbau, V. C-phycocyanin: A biliprotein with antioxidant, anti-inflammatory and neuroprotective effects. Curr. Protein Pept. Sci. 2003, 4, 207–216. [Google Scholar] [CrossRef]
- Jiang, L.; Wang, Y.; Yin, Q.; Liu, G.; Liu, H.; Huang, Y.; Li, B. Phycocyanin: A Potential Drug for Cancer Treatment. J. Cancer 2017, 8, 3416–3429. [Google Scholar] [CrossRef]
- Eriksen, N.T. Production of phycocyanin—A pigment with applications in biology, biotechnology, foods and medicine. Appl. Microbiol. Biotechnol. 2008, 80, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Brock, T.D. The Genus Cyanidium. In Thermophilic Microorganisms and Life at High Temperatures; Brock, T.D., Ed.; Springer: Berlin/Heidelberg, Germany, 1978. [Google Scholar]
- Hirooka, S.; Tomita, R.; Fujiwara, T.; Ohnuma, M.; Kuroiwa, H.; Kuroiwa, T.; Miyagishima, S.Y. Efficient open cultivation of cyanidialean red algae in acidified seawater. Sci. Rep. 2020, 10, 13794. [Google Scholar] [CrossRef]
- Hu, Q.; Marquardt, J.; Iwasaki, I.; Miyashita, H.; Kurano, N.; Mörschel, E.; Miyachi, S. Molecular structure, localization and function of biliproteins in the chlorophyll a/d containing oxygenic photosynthetic prokaryote Acaryochloris marina. Biochim. Biophys. Acta (BBA) Bioenerg. 1999, 1412, 250–261. [Google Scholar] [CrossRef]
- Ghosh, T.; Paliwal, C.; Maurya, R.; Mishra, S. Microalgal Rainbow Colours for Nutraceutical and Pharmaceutical Applications. In Plant Biology and Biotechnology: Volume I: Plant Diversity, Organization, Function and Improvement; Bahadur, B., Venkat Rajam, M., Sahijram, L., Krishnamurthy, K.V., Eds.; Springer: New Delhi, India, 2015; pp. 777–791. [Google Scholar]
- Singh, N.K.; Sonani, R.R.; Rastogi, R.P.; Madamwar, D. The phycobilisomes: An early requisite for efficient photosynthesis in cyanobacteria. EXCLI J. 2015, 14, 268–289. [Google Scholar] [CrossRef]
- Abalde, J.; Betancourt, L.; Torres, E.; Cid, A.; Barwell, C. Purification and characterization of phycocyanin from the marine cyanobacterium Synechococcus sp. IO9201. Plant Sci. 1998, 136, 109–120. [Google Scholar] [CrossRef]
- Padyana, A.K.; Bhat, V.B.; Madyastha, K.M.; Rajashankar, K.R.; Ramakumar, S. Crystal Structure of a Light-Harvesting Protein C-Phycocyanin from Spirulina platensis. Biochem. Biophys. Res. Commun. 2001, 282, 893–898. [Google Scholar] [CrossRef]
- Martelli, G.; Folli, C.; Visai, L.; Daglia, M.; Ferrari, D. Thermal stability improvement of blue colorant C-Phycocyanin from Spirulina platensis for food industry applications. Process Biochem. 2014, 49, 154–159. [Google Scholar] [CrossRef]
- Mercer, F.V.; Bogorad, L.; Mullens, R. Studies with Cyanidium caldarium. I. The fine structure and systematic position of the organism. J. Cell Biol. 1962, 13, 393–403. [Google Scholar] [CrossRef]
- Safaei, M.; Maleki, H.; Soleimanpour, H.; Norouzy, A.; Zahiri, H.S.; Vali, H.; Noghabi, K.A. Development of a novel method for the purification of C-phycocyanin pigment from a local cyanobacterial strain Limnothrix sp. NS01 and evaluation of its anticancer properties. Sci. Rep. 2019, 9, 9474. [Google Scholar] [CrossRef]
- Chittapun, S.; Jonjaroen, V.; Khumrangsee, K.; Charoenrat, T. C-phycocyanin extraction from two freshwater cyanobacteria by freeze thaw and pulsed electric field techniques to improve extraction efficiency and purity. Algal Res. 2020, 46, 101789. [Google Scholar] [CrossRef]
- Doke, J.M. An Improved and Efficient Method for the Extraction of Phycocyanin from Spirulina sp. Int. J. Food Eng. 2005, 1. [Google Scholar] [CrossRef]
- Schmidt, R.A.; Wiebe, M.G.; Eriksen, N.T. Heterotrophic high cell-density fed-batch cultures of the phycocyanin-producing red alga Galdieria sulphuraria. Biotechnol. Bioeng. 2005, 90, 77–84. [Google Scholar] [CrossRef]
- Martinez, J.M.; Luengo, E.; Saldana, G.; Alvarez, I.; Raso, J. C-phycocyanin extraction assisted by pulsed electric field from Artrosphira platensis. Food Res. Int. 2017, 99, 1042–1047. [Google Scholar] [CrossRef]
- Martínez, J.M.; Delso, C.; Álvarez, I.; Raso, J. Pulsed electric field permeabilization and extraction of phycoerythrin from Porphyridium cruentum. Algal Res. 2019, 37, 51–56. [Google Scholar] [CrossRef]
- Luengo, E.; Condón-Abanto, S.; Álvarez, I.; Raso, J. Effect of pulsed electric field treatments on permeabilization and extraction of pigments from Chlorella vulgaris. J. Membr. Biol. 2014, 247, 1269–1277. [Google Scholar] [CrossRef]
- Kurokawa, M.; King, P.M.; Wu, X.; Joyce, E.M.; Mason, T.J.; Yamamoto, K. Effect of sonication frequency on the disruption of algae. Ultrason. Sonochem. 2016, 31, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Schoenbach, K.H.; Peterkin, F.E.; Alden, R.W.; Beebe, S.J. The effect of pulsed electric fields on biological cells: Experiments and applications. IEEE Trans. Plasma Sci. 1997, 25, 284–292. [Google Scholar] [CrossRef]
- Qin, S.; Timoshkin, I.V.; Maclean, M.; Wilson, M.P.; MacGregor, S.J.; Given, M.J.; Anderson, J.G.; Wang, T. Pulsed Electric Field Treatment of Microalgae: Inactivation Tendencies and Energy Consumption. IEEE Trans. Plasma Sci. 2014, 42, 3191–3196. [Google Scholar] [CrossRef]
- Kotnik, T.; Frey, W.; Sack, M.; Haberl Meglic, S.; Peterka, M.; Miklavcic, D. Electroporation-based applications in biotechnology. Trends Biotechnol. 2015, 33, 480–488. [Google Scholar] [CrossRef]
- Scherer, D.; Krust, D.; Frey, W.; Mueller, G.; Nick, P.; Gusbeth, C. Pulsed electric field (PEF)-assisted protein recovery from Chlorella vulgaris is mediated by an enzymatic process after cell death. Algal Res. 2019, 41, 101536. [Google Scholar] [CrossRef]
- Zocher, K.; Banaschik, R.; Schulze, C.; Schulz, T.; Kredl, J.; Miron, C.; Schmidt, M.; Mundt, S.; Frey, W.; Kolb, J.F. Comparison of Extraction of Valuable Compounds from Microalgae by Atmospheric Pressure Plasmas and Pulsed Electric Fields. Plasma Med. 2016, 6, 273–302. [Google Scholar] [CrossRef]
- Zocher, K.; Lackmann, J.-W.; Volzke, J.; Steil, L.; Lalk, M.; Weltmann, K.-D.; Wende, K.; Kolb, J.F. Profiling microalgal protein extraction by microwave burst heating in comparison to spark plasma exposures. Algal Res. 2019, 39. [Google Scholar] [CrossRef]
- Zocher, K.; Rataj, R.; Steuer, A.; Weltmann, K.D.; Kolb, J.F. Mechanism of microalgae disintegration by spark discharge treatment for compound extraction. J. Phys. D Appl. Phys. 2020, 53. [Google Scholar] [CrossRef]
- Locke, B.R.; Thagard, S.M. Analysis and Review of Chemical Reactions and Transport Processes in Pulsed Electrical Discharge Plasma Formed Directly in Liquid Water. Plasma Chem. Plasma Process. 2012, 32, 875–917. [Google Scholar] [CrossRef]
- Eing, C.; Goettel, M.; Straessner, R.; Gusbeth, C.; Frey, W. Pulsed Electric Field Treatment of Microalgae—Benefits for Microalgae Biomass Processing. IEEE Trans. Plasma Sci. 2013, 41, 2901–2907. [Google Scholar] [CrossRef]
- Goettel, M.; Eing, C.; Gusbeth, C.; Straessner, R.; Frey, W. Pulsed electric field assisted extraction of intracellular valuables from microalgae. Algal Res. 2013, 2, 401–408. [Google Scholar] [CrossRef]
- Bennett, A.; Bogorad, L. Complementary chromatic adaptation in a filamentous blue-green alga. J. Cell Biol. 1973, 58, 419–435. [Google Scholar] [CrossRef]
- UniProt Consortium. UniProt: The Universal Protein Knowledgebase. Available online: https://www.uniprot.org/ (accessed on 25 February 2021).
- Van Eykelenburg, C. On the morphology and ultrastructure of the cell wall of Spirulina platensis. Antonie Leeuwenhoek 1977, 43, 89–99. [Google Scholar] [CrossRef]
- Sutanto, H.; Suzery, M. Phyocyanin extraction from microalgae Spirulina platensis assisted by ultrasound irradiation: Effect of time and temperature. Songklanakarin J. Sci. Technol. 2016, 38, 391–398. [Google Scholar]
- Li, Y.; Zhang, Z.; Paciulli, M.; Abbaspourrad, A. Extraction of phycocyanin-A natural blue colorant from dried spirulina biomass: Influence of processing parameters and extraction techniques. J. Food Sci. 2020, 85, 727–735. [Google Scholar] [CrossRef]
- Fratelli, C.; Burck, M.; de Amarante, M.C.A.; Braga, A.R.C. Antioxidant potential of nature’s “something blue”: Something new in the marriage of biological activity and extraction methods applied to C-phycocyanin. Trends Food Sci. Technol. 2020, 107, 309–323. [Google Scholar] [CrossRef]
- Akaberi, S.; Krust, D.; MŘller, G.; Frey, W.; Gusbeth, C. Impact of incubation conditions on protein and C-Phycocyanin recovery from Arthrospira platensis post-pulsed electric field treatment. Bioresour. Technol. 2020, 306, 123099. [Google Scholar] [CrossRef]
- Zhang, R.; Marchal, L.; Lebovka, N.; Vorobiev, E.; Grimi, N. Two-step procedure for selective recovery of bio-molecules from microalga Nannochloropsis oculata assisted by high voltage electrical discharges. Bioresour. Technol. 2020, 302, 122893. [Google Scholar] [CrossRef]
- Apt, K.E.; Collier, J.L.; Grossman, A.R. Evolution of the Phycobiliproteins. J. Molecular Biol. 1995, 248, 79–96. [Google Scholar] [CrossRef]
- Kremer, B.P.; Feige, G.B. Accumulation of Photoassimilatory Products by Phycobiliprotein- Containing Algae with Special Reference to Cyanidium caldarium. Z. Naturforschung C 1979, 34, 1209–1214. [Google Scholar] [CrossRef][Green Version]
- Troxler, R.F.; Ehrhardt, M.M.; Brown-Mason, A.S.; Offner, G.D. Primary structure of phycocyanin from the unicellular rhodophyte Cyanidium caldarium. II. Complete amino acid sequence of the beta subunit. J. Biol. Chem. 1981, 256, 12176–12184. [Google Scholar] [CrossRef]
- Böcker, L.; Ortmann, S.; Surber, J.; Leeb, E.; Reineke, K.; Mathys, A. Biphasic short time heat degradation of the blue microalgae protein phycocyanin from Arthrospira platensis. Innov. Food Sci. Emerg. Technol. 2019, 52, 116–121. [Google Scholar] [CrossRef]
- Ocarra, P. Purification and N-terminal analysis of algal biliproteins. Biochem. J. 1965, 94, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Scheer, H.; Zhao, K.H. Biliprotein maturation: The chromophore attachment. Mol. Microbiol. 2008, 68, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Stadnichuk, I.N. Phycobiliproteins: Determination of chromophore composition and content. Phytochem. Anal. 1995, 6, 281–288. [Google Scholar] [CrossRef]
- Romay, C.; Gonzalez, R.; Pizarro, M.; Lissi, E. Kinetics of c-phycocyanin reaction with hypochlorite. J. Protein Chem. 2000, 19, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Stec, B.; Troxler, R.F.; Teeter, M.M. Crystal Structure of C-Phycocyanin from Cyanidium caldarium Provides a New Perspective on Phycobilisome Assembly. Biophys. J. 1999, 76, 2912–2921. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Eluent B | Time [min] | Flow [µL/min] |
|---|---|---|
| 5% | 0 | 0.300 |
| 5% | 4 | 0.300 |
| 5% | 6 | 0.300 |
| 35% | 30 | 0.300 |
| 80% | 33 | 0.300 |
| 80% | 35 | 0.300 |
| 5% | 36 | 0.300 |
| 5% | 48 | 0.300 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sommer, M.-C.; Balazinski, M.; Rataj, R.; Wenske, S.; Kolb, J.F.; Zocher, K. Assessment of Phycocyanin Extraction from Cyanidium caldarium by Spark Discharges, Compared to Freeze-Thaw Cycles, Sonication, and Pulsed Electric Fields. Microorganisms 2021, 9, 1452. https://doi.org/10.3390/microorganisms9071452
Sommer M-C, Balazinski M, Rataj R, Wenske S, Kolb JF, Zocher K. Assessment of Phycocyanin Extraction from Cyanidium caldarium by Spark Discharges, Compared to Freeze-Thaw Cycles, Sonication, and Pulsed Electric Fields. Microorganisms. 2021; 9(7):1452. https://doi.org/10.3390/microorganisms9071452
Chicago/Turabian StyleSommer, Marie-Christine, Martina Balazinski, Raphael Rataj, Sebastian Wenske, Juergen F. Kolb, and Katja Zocher. 2021. "Assessment of Phycocyanin Extraction from Cyanidium caldarium by Spark Discharges, Compared to Freeze-Thaw Cycles, Sonication, and Pulsed Electric Fields" Microorganisms 9, no. 7: 1452. https://doi.org/10.3390/microorganisms9071452
APA StyleSommer, M.-C., Balazinski, M., Rataj, R., Wenske, S., Kolb, J. F., & Zocher, K. (2021). Assessment of Phycocyanin Extraction from Cyanidium caldarium by Spark Discharges, Compared to Freeze-Thaw Cycles, Sonication, and Pulsed Electric Fields. Microorganisms, 9(7), 1452. https://doi.org/10.3390/microorganisms9071452