
The Role of Metal Ions in Fungal Organic Acid Accumulation


Abstract
1. Introduction
2. Biology of First-Row Transition Metal Ions in Fungi
3. The Influence of Transition Metal Ions on Accumulation of Mono-and Dicarboxylic Organic Acids by Fungi
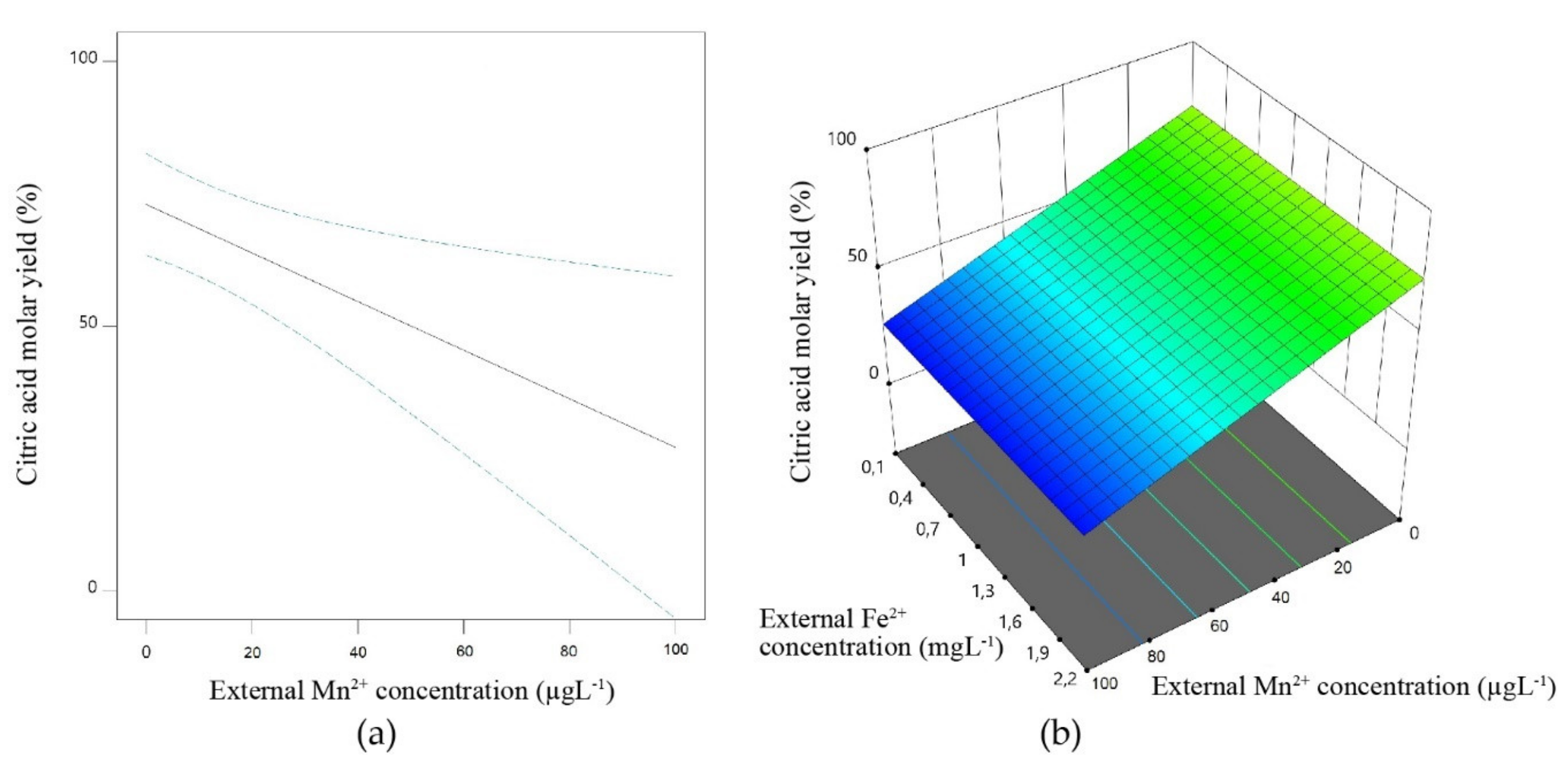
4. The Influence of Transition Metal Ions on Accumulation of Citric- and Itaconic Acid by Fungi
5. Biochemical and Physiological Interpretations of the Roles of Manganese Ions on Citric and Itaconic Acid Accumulation
6. Metal Ions-Related Aspects of Submerged Organic Acid Fermentations
7. Metal Ion Analysis in Organic Acid Fermentations
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Werpy, T.; Petersen, G. Top Value Added Chemicals from Biomass: Volume I—Results of Screening for Potential Candidates from Sugars and Synthesis Gas; Pacific Northwest National Laboratory (PNNL) and National Renewable Energy Laboratory (NREL), Office of Scientific and Technical Information (OSTI): Golden, CO, USA, 2004. Available online: https://www.osti.gov/servlets/purl/15008859 (accessed on 10 June 2021).
- Karaffa, L.; Kubicek, C.P. Production of organic acids by fungi. In Encyclopedia of Mycology, 1st ed.; Zaragoza, O., Casadevall, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Foster, J.W. Chemical Activities of Fungi; New York Academic Press Inc.: New York, NY, USA, 1949. [Google Scholar]
- Shu, P.; Johnson, M.J. Effect of the composition of the sporulation medium on citric acid production by Aspergillus niger in submerged culture. J. Bacteriol. 1947, 54, 161–167. [Google Scholar] [CrossRef]
- Shu, P.; Johnson, M.J. The interdependence of medium constituents in citric acid production by submerged fermentation. J. Bacteriol. 1948, 56, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Shu, P.; Johnson, M.J. Citric acid production by submerged fermentation with Aspergillus niger. Ind. Eng. Chem. 1948, 40, 1202–1205. [Google Scholar] [CrossRef]
- Bernhauer, K.; Knobloch, H.; Iglauer, A. Formation of acid from sugar by Aspergillus niger. XI. Factors determining the accumulation of citric acid. Biochem. Z. 1941, 309, 151–178. [Google Scholar]
- Butkewitsch, W.; Timofeewa, A.G. Einfluss einzelner mineralischer Elemente des Niahrmediums auf die Säurebildung bei Aspergillus niger. Biochem. Z. 1935, 275, 405–415. [Google Scholar]
- Chrzaszcz, T.; Peyros, E. Optimale Bedingungen der Citronensiaureanhaufung, sowie einige Beobachtungen zur Theorie der Zitronen säurebildung. Biochem. Z. 1935, 280, 325–336. [Google Scholar]
- Currie, J.N. The citric acid fermentation of Aspergillus niger. J. Biol. Chem. 1917, 31, 15–37. [Google Scholar] [CrossRef]
- Doelger, W.P.; Prescott, S.C. Citric Acid Fermentation. Ind. Eng. Chem. Res. 1934, 26, 1142–1149. [Google Scholar] [CrossRef]
- Gerhardt, P.; Dorrell, W.W.; Baldwin, I.L. Citric acid fermentation of beet molasses. J. Bacteriol. 1946, 52, 555–564. [Google Scholar] [CrossRef]
- Von Loesecke, H.W. A review of information on: Mycological citric acid production. Chem. Eng. News 1945, 23, 1952–1959. [Google Scholar] [CrossRef]
- Perlman, D.; Dorrell, W.W.; Johnson, M.J. Effect of metallic ions on the production of citric acid by Aspergillus niger. Arch. Biochem. Biophys. 1946, 11, 131–143. [Google Scholar]
- Perlman, D.; Kita, D.A.; Peterson, W.H. Production of citric acid from cane molasses. Arch. Biochem. Biophys. 1946, 11, 123–129. [Google Scholar]
- Porges, N. Citric acid production by Aspergillus niger. Am. J. Bot. 1932, 19, 559–567. [Google Scholar] [CrossRef]
- Xu, D.B.; Röhr, M.; Kubicek, C.P. Aspergillus niger cyclic AMP levels are not influenced by manganese deficiency and do not correlate with citric acid accumulation. Appl. Microbiol. Biotechnol. 1989, 32, 124–128. [Google Scholar] [CrossRef]
- Kaplan, C.D.; Kaplan, J. Iron acquisition and transcriptional regulation. Chem. Rev. 2009, 109, 4536–4552. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Gutteridge, J.M.C. Oxygen toxicity, oxygen radicals, transition metals and disease. Biochem. J. 1984, 219, 1–14. [Google Scholar] [CrossRef]
- Hansberg, W.; Salas-Lizana, R.; Domínguez, L. Fungal catalases: Function, phylogenetic origin and structure. Arch. Biochem. Biophys. 2012, 525, 170–180. [Google Scholar] [CrossRef]
- Auld, D.S. Zinc coordination sphere in biochemical zinc sites. BioMetals 2001, 14, 271–313. [Google Scholar] [CrossRef] [PubMed]
- Kadrmas, J.L.; Beckerle, M.C. The LIM domain: From the cytoskeleton to the nucleus. Nat. Rev. Mol. Cell Biol. 2004, 5, 920–931. [Google Scholar] [CrossRef]
- Borden, K.L.B.; Freemont, P.S. The RING finger domain: A recent example of a sequence-structure family. Curr. Opin. Struct. Biol. 1996, 6, 395–401. [Google Scholar] [CrossRef]
- Eide, D.J. The molecular biology of metal ion transport in Saccharomyces cerevisiae. Annu. Rev. Nutr. 1998, 18, 441–469. [Google Scholar] [CrossRef] [PubMed]
- Eide, D.J. Multiple regulatory mechanisms maintain zinc homeostasis in Saccharomyces cerevisiae. J. Nutr. 2003, 133 (Suppl. S1), 1532S–1535S. [Google Scholar] [CrossRef] [PubMed]
- Andreini, C.; Bertini, I. A bioinformatics view of zinc enzymes. J. Inorg. Biochem. 2012, 111, 150–156. [Google Scholar] [CrossRef]
- Festa, R.A.; Thiele, D.J. Copper: An essential metal in biology. Curr. Biol. 2011, 21, R877–R883. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.D.; Logeman, B.L.; Thiele, D.J. Copper acquisition and utilization in fungi. Annu. Rev. Microbiol. 2017, 71, 597–623. [Google Scholar] [CrossRef]
- Antsotegi-Uskola, M.; Markina-Iñarrairaegui, A.; Ugalde, U. Copper homeostasis in Aspergillus nidulans involves coordinated transporter function, expression and cellular dynamics. Front. Microbiol. 2020, 11, 555306. [Google Scholar] [CrossRef]
- Ding, C.; Yin, J.; Tovar, E.M.M.; Fitzpatrick, D.A.; Higgins, D.G.; Thiele, D.J. The copper regulon of the human fungal pathogen Cryptococcus neoformans H99. Mol. Microbiol. 2011, 81, 1560–1576. [Google Scholar] [CrossRef]
- García-Santamarina, S.; Thiele, D.J. Copper at the fungal pathogen-host axis. J. Biol. Chem. 2015, 290, 18945–18953. [Google Scholar] [CrossRef] [PubMed]
- Sándor, E.; Kolláth, I.S.; Fekete, E.; Bíró, V.; Flipphi, M.; Kovács, B.; Kubicek, C.P.; Karaffa, L. Carbon-source dependent interplay of copper and manganese ions modulates the morphology and itaconic acid production in Aspergillus terreus. Front. Microbiol. 2021, 12, 680420. [Google Scholar] [CrossRef]
- Shaw, C.E.; Kapica, L. Production of diagnostic pigment by phenoloxidase activity of Cryptococcus neoformans. Appl. Microbiol. 1972, 24, 824–830. [Google Scholar] [CrossRef]
- Williamson, P.R. Biochemical and molecular characterization of the diphenol oxidase of Cryptococcus neoformans: Identification as a laccase. J. Bacteriol. 1994, 176, 656–664. [Google Scholar] [CrossRef]
- Reddi, A.R.; Jensen, L.T.; Culotta, V.C. Manganese homeostasis in Saccharomyces cerevisiae. Chem. Rev. 2009, 109, 4722–4732. [Google Scholar] [CrossRef] [PubMed]
- Pisat, N.P.; Pandey, A.; MacDiarmid, C.W. MNR2 regulates intracellular magnesium storage in Saccharomyces cerevisiae. Genetics 2009, 183, 873–884. [Google Scholar] [CrossRef] [PubMed]
- Karaffa, L.; Kubicek, C.P. Citric acid and itaconic acid accumulation: Variations of the same story? Appl. Microbiol. Biotechnol. 2019, 103, 2889–2902. [Google Scholar] [CrossRef] [PubMed]
- Battat, E.; Peleg, Y.; Bercovitz, A.; Rokem, J.S.; Goldberg, I. Optimization of L-malic acid production by Aspergillus flavus in a stirred fermentor. Biotechnol. Bioeng. 1991, 37, 1108–1116. [Google Scholar] [CrossRef]
- Xu, Y.; Zhou, Y.; Cao, W.; Liu, H. Improved Production of malic acid in Aspergillus niger by abolishing citric acid accumulation and enhancing glycolytic flux. ACS Synth. Biol. 2020, 9, 1418–1425. [Google Scholar] [CrossRef] [PubMed]
- Waksman, S.A.; Foster, J.W. Respiration and lactic acid production by a fungus of the genus Rhizopus. J. Agric. Res. 1938, 57, 873. [Google Scholar]
- Ge, C.; Pan, R.; Zhang, J.; Cai, J.; Yu, Z. Effect of ZnSO4 on L-lactic acid production by Rhizopus oryzae. Wei Sheng Wu Xue Bao 2013, 53, 515–520. [Google Scholar]
- Lockwood, L.B.; Ward, G.E.; May, O.E. The physiology of Rhizopus oryzae. J. Agric. Res. 1936, 53, 849–857. [Google Scholar]
- Noguchi, Y.; Johnson, M.J. Citric acid fermentation of sugars purified with chelating resin. J. Bacteriol. 1961, 82, 538–541. [Google Scholar] [CrossRef]
- Clark, D.S.; Ito, K.; Horitsu, H. Effect of manganese and other heavy metals on submerged citric acid fermentation of molasses. Biotechnol. Bioeng. 1966, 8, 465–471. [Google Scholar] [CrossRef]
- Kitos, P.A.; Campbell, J.J.; Tomlinson, N. Influence of temperature on the trace element requirements for citric acid production by Aspergillus niger. Appl. Microbiol. 1953, 1, 156–159. [Google Scholar] [CrossRef]
- Sánchez-Marroquín, A.; Carreño, R.; Ledezma, M. Effect of trace elements on citric acid fermentation by Aspergillus niger. Appl. Microbiol. 1970, 20, 888–892. [Google Scholar] [CrossRef] [PubMed]
- Papagianni, M.; Mattey, M. Morphological development of Aspergillus niger in submerged citric acid fermentation as a function of the spore inoculum level. Application of neural network and cluster analysis for characterization of mycelial morphology. Microb. Cell Fact. 2006, 5, 3. [Google Scholar] [CrossRef]
- Guilherme, A.A.; Pinto, A.S.; Rodrigues, S. Optimization of trace metals concentration on citric acid production by Aspergillus niger NRRL 2001. Food Bioprocess Technol. 2008, 1, 246–253. [Google Scholar] [CrossRef]
- Fejes, B.; Ouedraogo, J.P.; Fekete, E.; Sándor, E.; Flipphi, M.; Soós, Á.; Molnár, Á.P.; Kovács, B.; Kubicek, C.P.; Tsang, A.; et al. The effects of external Mn2+ concentration on hyphal morphology and citric acid production are mediated primarily by the NRAMP-family transporter DmtA in Aspergillus niger. Microb. Cell Fact. 2020, 19, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Szczodrak, J.; Ilczuk, Z. Effect of iron on the activity of aconitate hydratase and synthesis of citric acid by Aspergillus niger. Zentralbl. Mikrobiol. 1985, 140, 567–574. [Google Scholar] [CrossRef]
- Odoni, D.I.; van Gaal, M.P.; Schonewille, T.; Tamayo-Ramos, J.A.; Martins dos Santos, V.A.P.; Suarez-Diez, M.; Schaap, P.J. Aspergillus niger secretes citrate to increase iron bioavailability. Front. Microbiol. 2017, 8, 1424. [Google Scholar] [CrossRef] [PubMed]
- Wold, W.S.M.; Suzuki, I. The citric acid fermentation by Aspergillus niger: Regulation by zinc of growth and acidogenesis. Can. J. Microbiol. 1976, 22, 1083–1092. [Google Scholar] [CrossRef]
- Wold, W.S.M.; Suzuki, I. Regulation by zinc and adenosine 3′,5′ cyclic monophosphate of growth and citric acid accumulation in Aspergillus niger. Can. J. Microbiol. 1976, 22, 1093–1101. [Google Scholar] [CrossRef]
- Kubicek-Pranz, E.M.; Mozelt, M.; Röhr, M.; Kubicek, C.P. Changes in the concentration of fructose 2,6-bisphosphate in Aspergillus niger during stimulation of acidogenesis by elevated sucrose concentration. BBA Gen. Subj. 1990, 1033, 250–255. [Google Scholar] [CrossRef]
- Xu, D.B.; Kubicek, C.P.; Röhr, M. A comparison of factors influencing citric acid production by Aspergillus niger grown in submerged culture and on filter paper. Appl. Microbiol. Biotechnol. 1989, 30, 444–449. [Google Scholar] [CrossRef]
- Batti, M.; Schweiger, L.B. Process for the Production of Itaconic Acid. U.S. Patent US3078217A, 19 February 1963. [Google Scholar]
- Karaffa, L.; Díaz, R.; Papp, B.; Fekete, E.; Sándor, E.; Kubicek, C.P. A deficiency of manganese ions in the presence of high sugar concentrations is the critical parameter for achieving high yields of itaconic acid by Aspergillus terreus. Appl. Microbiol. Biotechnol. 2015, 99, 7937–7944. [Google Scholar] [CrossRef]
- Saha, B.C.; Kennedy, G.J.; Bowman, M.J.; Qureshi, N.; Dunn, R.O. Factors affecting production of itaconic acid from mixed sugars by Aspergillus terreus. Appl. Biochem. Biotechnol. 2019, 187, 449–460. [Google Scholar] [CrossRef] [PubMed]
- Shin, W.S.; Lee, D.; Kim, S.; Jeong, Y.S.; Chun, G.T. Application of scale-up criterion of constant oxygen mass transfer coefficient (Kla) for production of itaconic acid in a 50 l pilot-scale fermentor by fungal cells of aspergillus terreus. J. Microbiol. Biotechnol. 2013, 23, 1445–1453. [Google Scholar] [CrossRef]
- Shin, W.S.; Park, B.; Lee, D.; Oh, M.K.; Chun, G.T.; Kim, S. Enhanced production of itaconic acid through development of transformed fungal strains of Aspergillus terreus. J. Microbiol. Biotechnol. 2017, 27, 306–315. [Google Scholar] [CrossRef]
- Ma, H.; Kubicek, C.P.; Röhr, M. Metabolic effects of manganese deficiency in Aspergillus niger: Evidence for increased protein degradation. Arch. Microbiol. 1985, 141, 266–268. [Google Scholar] [CrossRef]
- Hockertz, S.; Plönzig, J.; Auling, G. Impairment of DNA formation is an early event in Aspergillus niger under manganese starvation. Appl. Microbiol. Biotechnol. 1987, 25, 590–593. [Google Scholar] [CrossRef]
- Meixner, O.; Mischak, H.; Kubicek, C.P.; Röhr, M. Effect of manganese deficiency on plasma-membrane lipid composition and glucose uptake in Aspergillus niger. FEMS Microbiol. Lett. 1985, 26, 271–274. [Google Scholar] [CrossRef]
- Kisser, M.; Kubicek, C.P.; Röhr, M. Influence of manganese on morphology and cell wall composition of Aspergillus niger during citric acid fermentation. Arch. Microbiol. 1980, 128, 26–33. [Google Scholar] [CrossRef]
- Borrello, S.; De Leo, M.E.; Galeotti, T. Transcriptional regulation of MnSOD by manganese in the liver of manganese-deficent mice and during rat development. Biochem. Int. 1992, 28, 595–601. [Google Scholar]
- Allen, M.D.; Kropat, J.; Tottey, S.; Del Campo, J.A.; Merchant, S.S. Manganese deficiency in chlamydomonas results in loss of photosystem II and MnSOD function, sensitivity to peroxides, and secondary phosphorus and iron deficiency. Plant Physiol. 2007, 143, 263–277. [Google Scholar] [CrossRef]
- Onodera, J.; Ohsumi, Y. Autophagy is required for maintenance of amino acid levels and protein synthesis under nitrogen starvation. J. Biol. Chem. 2005, 280, 31582–31586. [Google Scholar] [CrossRef]
- Kaur, B.; Punekar, N.S. Autophagy is important to the acidogenic metabolism of Aspergillus niger. PLoS ONE 2019, 14, e0223895. [Google Scholar] [CrossRef]
- Richie, D.L.; Fuller, K.K.; Fortwendel, J.; Miley, M.D.; McCarthy, J.W.; Feldmesser, M.; Rhodes, J.C.; Askew, D.S. Unexpected link between metal ion deficiency and autophagy in Aspergillus fumigatus. Eukaryot. Cell 2007, 6, 2437–2447. [Google Scholar] [CrossRef]
- Schreferl, G.; Kubicek, C.P.; Röhr, M. Inhibition of citric acid accumulation by manganese ions in Aspergillus niger mutants with reduced citrate control of phosphofructokinase. J. Bacteriol. 1986, 165, 1019–1022. [Google Scholar] [CrossRef]
- Habison, A.; Kubicek, C.P.; Röhr, M. Partial purification and regulatory properties of phosphofructokinase from Aspergillus niger. Biochem. J. 1983, 209, 669–676. [Google Scholar] [CrossRef]
- Sakai, A.; Kusumoto, A.; Kiso, Y.; Furuya, E. Itaconate reduces visceral fat by inhibiting fructose 2,6-bisphosphate synthesis in rat liver. Nutrition 2004, 20, 997–1002. [Google Scholar] [CrossRef]
- Tevž, G.; Benčina, M.; Legiša, M. Enhancing itaconic acid production by Aspergillus terreus. Appl. Microbiol. Biotechnol. 2010, 87, 1657–1664. [Google Scholar] [CrossRef]
- Kontopidis, G.; Mattey, M.; Kristiansen, B. Citrate transport during the citric acid fermentation by Aspergillus niger. Biotechnol. Lett. 1995, 17, 1101–1106. [Google Scholar] [CrossRef]
- Netik, A.; Torres, N.V.; Riol, J.M.; Kubicek, C.P. Uptake and export of citric acid by Aspergillus niger is reciprocally regulated by manganese ions. Biochim. Biophys. Acta Biomembr. 1997, 1326, 287–294. [Google Scholar] [CrossRef][Green Version]
- Jensen, L.T.; Ajua-Alemanji, M.; Culotta, V.C. The Saccharomyces cerevisiae high affinity phosphate transporter encoded by PHO84 also functions in manganese homeostasis. J. Biol. Chem. 2003, 278, 42036–42040. [Google Scholar] [CrossRef] [PubMed]
- Matzapetakis, M.; Karligiano, N.; Bino, A.; Dakanali, M.; Raptopoulou, C.P.; Tangoulis, V.; Terzis, A.; Giapintzakis, J.; Salifoglou, A. Manganese citrate chemistry: Syntheses, spectroscopic studies, and structural characterizations of novel mononuclear, water-soluble manganese citrate complexes. Inorg. Chem. 2000, 39, 4044–4051. [Google Scholar] [CrossRef]
- Steiger, M.G.; Rassinger, A.; Mattanovich, D.; Sauer, M. Engineering of the citrate exporter protein enables high citric acid production in Aspergillus niger. Metab. Eng. 2019, 52, 224–231. [Google Scholar] [CrossRef]
- Odoni, D.I.; Vazquez-Vilar, M.; Van Gaal, M.P.; Schonewille, T.; Martins Dos Santos, V.A.P.; Tamayo-Ramos, J.A.; Suarez-Diez, M.; Schaap, P.J. Aspergillus niger citrate exporter revealed by comparison of two alternative citrate producing conditions. FEMS Microbiol. Lett. 2019, 366, fnz071. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.; Dai, Z.; Swita, M.; Pomraning, K.R.; Hofstad, B.; Panisko, E.; Baker, S.; Magnuson, J. Deletion analysis of the itaconic acid biosynthesis gene cluster components in Aspergillus pseudoterreus ATCC32359. Appl. Microbiol. Biotechnol. 2020, 104, 3981–3992. [Google Scholar] [CrossRef] [PubMed]
- Cox, P.W.; Thomas, C.R. Classification and measurement of fungal pellets by automated image analysis. Biotechnol. Bioeng. 1992, 39, 945–952. [Google Scholar] [CrossRef]
- Schweiger, L.B. Production of Citric Acid by Fermentation. U.S. Patent 2970084, 31 January 1961. [Google Scholar]
- Michel, F.C.; Grulke, E.A.; Reddy, C.A. Determination of the respiration kinetics for mycelial pellets of Phanerochaete chrysosporium. Appl. Environ. Microbiol. 1992, 58, 1740–1745. [Google Scholar] [CrossRef]
- Kubicek, C.P.; Zehentgruber, O.; El-Kalak, H.; Röhr, M. Regulation of citric acid production by oxygen: Effect of dissolved oxygen tension on adenylate levels and respiration in Aspergillus niger. Eur. J. Appl. Microbiol. Biotechnol. 1980, 9, 101–115. [Google Scholar] [CrossRef]
- Park, Y.S.; Ohta, N.; Okabe, M. Effect of dissolved oxygen concentration and impeller tip speed on itaconic acid production by Aspergillus terreus. Biotechnol. Lett. 1993, 15, 583–586. [Google Scholar] [CrossRef]
- Molnár, Á.P.; Németh, Z.; Kolláth, I.S.; Fekete, E.; Flipphi, M.; Ág, N.; Soós, Á.; Kovács, B.; Sándor, E.; Kubicek, C.P.; et al. High oxygen tension increases itaconic acid accumulation, glucose consumption, and the expression and activity of alternative oxidase in Aspergillus terreus. Appl. Microbiol. Biotechnol. 2018, 102, 8799–8808. [Google Scholar] [CrossRef] [PubMed]
- Gyamerah, M. Factors affecting the growth form of Aspergillus terreus NRRL 1960 in relation to itaconic acid fermentation. Appl. Microbiol. Biotechnol. 1995, 44, 356–361. [Google Scholar] [CrossRef]
- Dai, Z.; Mao, X.; Magnuson, J.K.; Lasure, L.L. Identification of genes associated with morphology in Aspergillus niger by using suppression subtractive hybridization. Appl. Environ. Microbiol. 2004, 70, 2474–2485. [Google Scholar] [CrossRef][Green Version]
- Vandenberghe, L.P.S.; Soccol, C.R.; Pandey, A.; Lebeault, J.-M. Microbial production of citric acid. Braz. Arch. Biol. Technol. 1999, 42, 263–276. [Google Scholar] [CrossRef]
- Shackelford, J.F.; William, A. The CRC Materials Science and Engineering Handbook, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2001. [Google Scholar]
- Clark, D.S.; Ito, K.; Tymchuk, P. Effect of potassium ferrocyanide on the chemical composition of molasses mash used in the citric acid fermentation. Biotechnol. Bioeng. 1965, 7, 269–278. [Google Scholar] [CrossRef]
- Lönnerdal, B. Phytic acid-trace element (Zn, Cu, Mn) interactions. Int. J. Food Sci. Technol. 2002, 37, 749–758. [Google Scholar] [CrossRef]
- Moyer, A.J. Effect of alcohols on the mycological production of citric acid in surface and submerged culture. Appl. Microbiol. 1953, 1, 1–7. [Google Scholar] [CrossRef]
- Batti, M.A. Fermentation Process for Producing Citric Acid. U.S. Patent 3438863A, 15 April 1969. [Google Scholar]
- Paluch, E.; Szperlik, J.; Czuj, T.; Cal, M.; Lamch, Ł.; Wilk, K.A.; Obłąk, E. Multifunctional cationic surfactants with a labile amide linker as efficient antifungal agents—mechanisms of action. Appl. Microbiol. Biotechnol. 2021, 105, 1237–1251. [Google Scholar] [CrossRef] [PubMed]
- Yalage Don, S.M.; Schmidtke, L.M.; Gambetta, J.M.; Steel, C.C. Volatile organic compounds produced by Aureobasidium pullulans induce electrolyte loss and oxidative stress in Botrytis cinerea and Alternaria alternata. Res. Microbiol. 2021, 172, 103788. [Google Scholar] [CrossRef]
- Types of Ion Detector for Mass Spectrometry | Technology Networks. Available online: https://www.technologynetworks.com/analysis/articles/types-of-ion-detector-for-mass-spectrometry-347890 (accessed on 8 June 2021).


{kind=link}
| Organic Acid | Producer Micro-Organism(s) | Applications |
|---|---|---|
| L-lactic acid | Lactobacillus spp. Lactococcus spp. Rhizopus oryzae | Excipient in food, cosmetics, pharmaceutical and chemical industries. Building block for poly-lactic acid. |
| Oxalic acid | Aspergillus niger | Food industry, pharmaceuticals, waste water treatment, hydrometallurgy |
| L-malic acid | Aspergillus niger Aspergillus oryzae Rhizopus oryzae | Nutritional bars, protein drinks, functional beverages, pharmaceutical, cosmetic and personal care products |
| Fumaric acid | Rhizopus oryzae | Food acidulant, mordant for dyes. Its esters are used to treat relapsing-remitting multiple sclerosis and have immunomodulating activities. |
| Succinic Acid | Saccharomyces cerevisiae Escherichia coli Anaerobiospirillum succiniciproducens Actinobacillus succinogenes Basfia succiniciproducens | Food additives, detergents, pigments, toners, cosmetics, cement additives, pharmaceuticals, resins coatings Building block for 1,4-butanediol, poly-butyl succinate, polybutylene succinate-co-butylene terephthalate. |
| Trans-2,3-Epoxysuccinic acid | Aspergillus clavatus Paecilomyces varioti Paecilomyces elegans Penicillium vineferum Talaromyces wortamannii | Building block for optically specific single β-lactam antibiotics and polyepoxysuccinic acid plasticizer stabilizer, corrosion inhibitor, biotransformation to meso-tartaric acid. |
| Citric acid | Aspergillus niger | Flavoring agent and preservative in food and beverages. Emulsifying agent. Delivery of metal ions as citrate salts in dietary supplements. Acidifier and chelating agent in the chemical and pharmaceutical industries. |
| Itaconic acid | Aspergillus terreus | Pharmaceutical, architectural, paper, paint, and medical industries as plastics, resins, paints, synthetic fibers, plasticizers, detergents Drug delivery in medicine. |
| Gluconic acid | Gluconobacter sp. Aspergillus niger | Flavoring agent in meat, wine and dairy products. Counterion during therapeutic calcium and/or iron administration. Removal of calcareous and rust deposits from surfaces. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karaffa, L.; Fekete, E.; Kubicek, C.P. The Role of Metal Ions in Fungal Organic Acid Accumulation. Microorganisms 2021, 9, 1267. https://doi.org/10.3390/microorganisms9061267
Karaffa L, Fekete E, Kubicek CP. The Role of Metal Ions in Fungal Organic Acid Accumulation. Microorganisms. 2021; 9(6):1267. https://doi.org/10.3390/microorganisms9061267
Chicago/Turabian StyleKaraffa, Levente, Erzsébet Fekete, and Christian P. Kubicek. 2021. "The Role of Metal Ions in Fungal Organic Acid Accumulation" Microorganisms 9, no. 6: 1267. https://doi.org/10.3390/microorganisms9061267
APA StyleKaraffa, L., Fekete, E., & Kubicek, C. P. (2021). The Role of Metal Ions in Fungal Organic Acid Accumulation. Microorganisms, 9(6), 1267. https://doi.org/10.3390/microorganisms9061267