
Salmonella Bacterin Vaccination Decreases Shedding and Colonization of Salmonella Typhimurium in Pigs




Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Selection
2.2. Experimental Design and Sample Collection
2.3. Blood Count
2.4. Serological Analysis (IgG)
2.5. Bacteriological Analysis
2.6. Statistical Analysis
3. Results
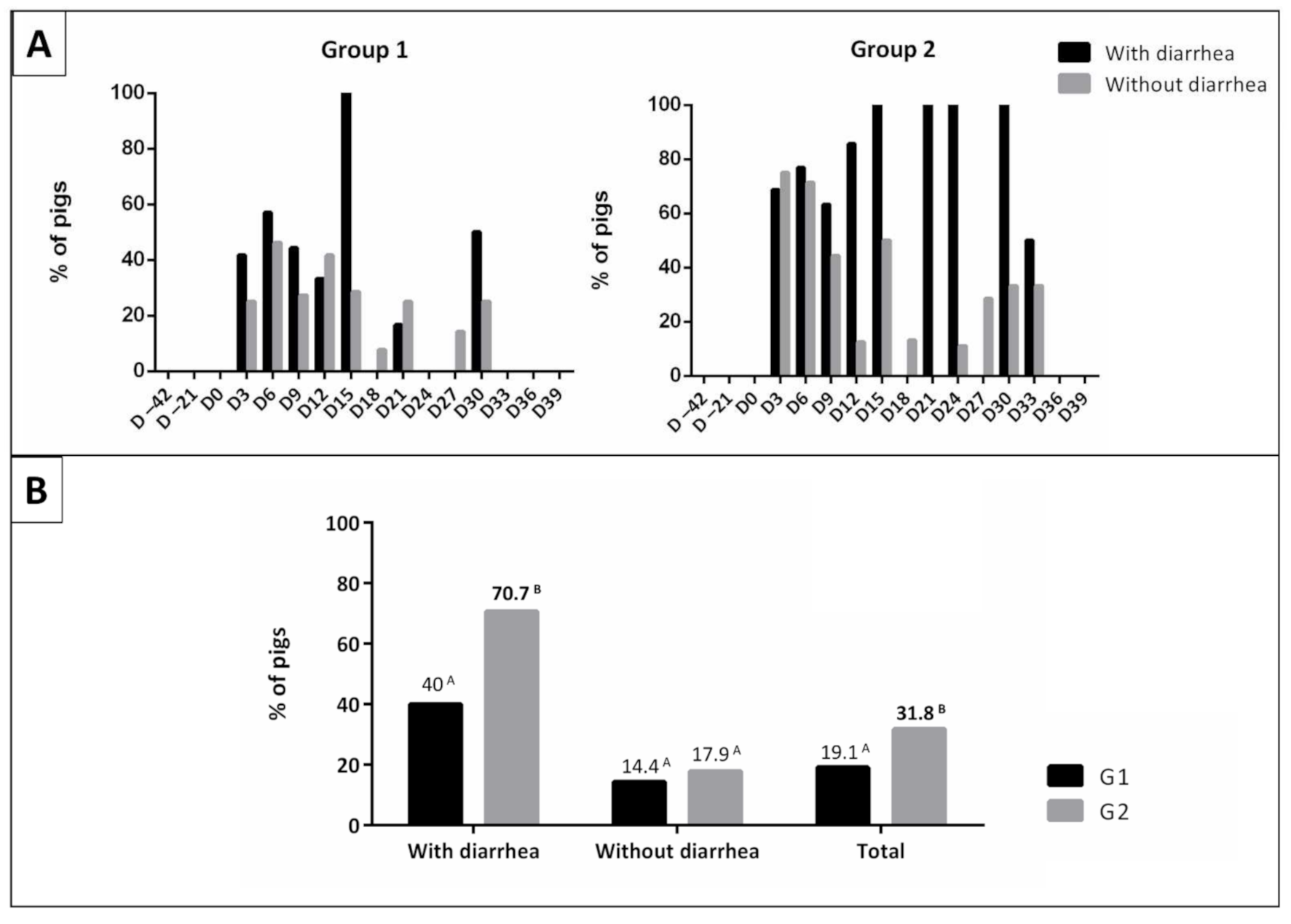
3.1. Physical Examination
3.2. Erythrogram and Platelet Count
3.3. Leukogram
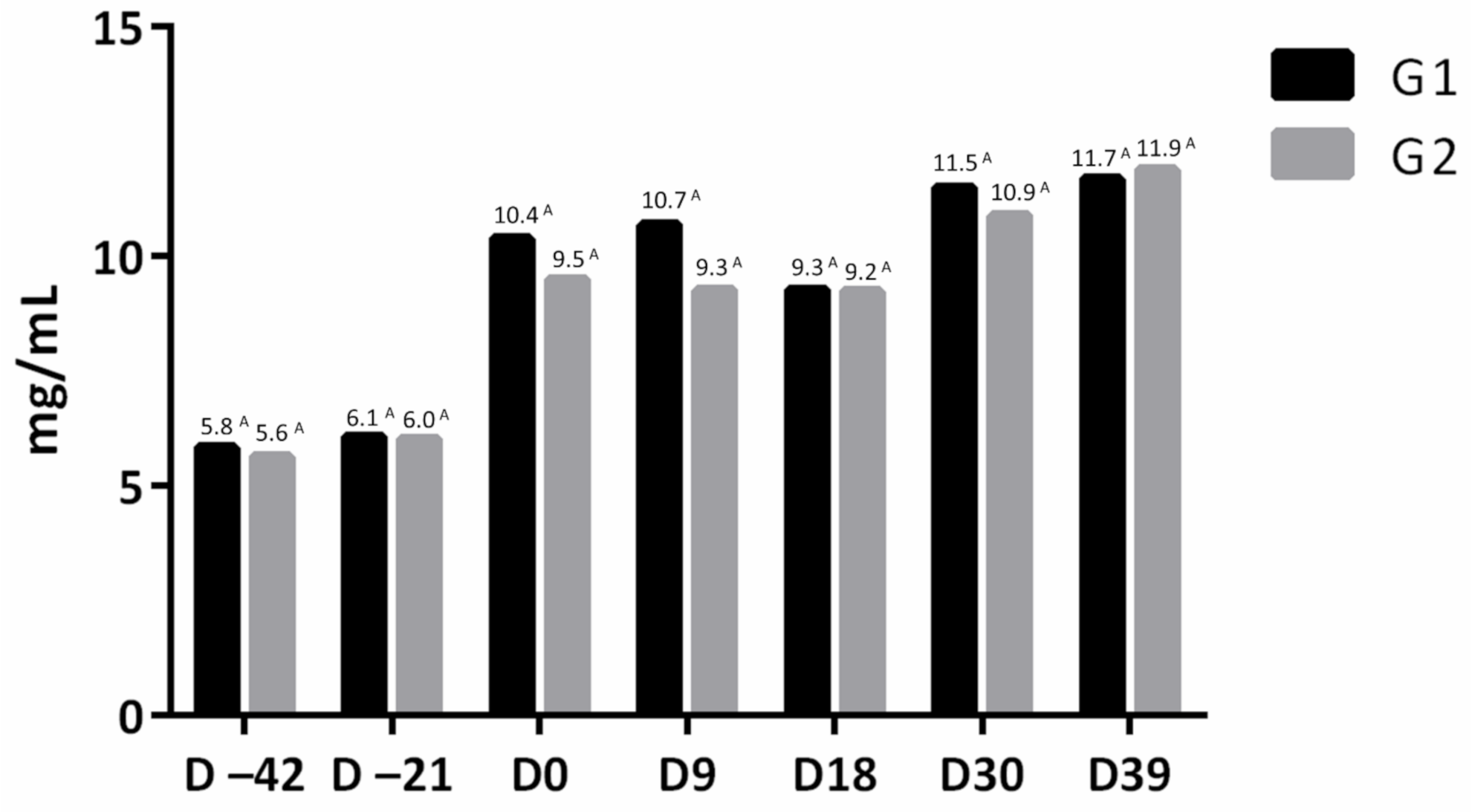
3.4. Serum IgG Concentration
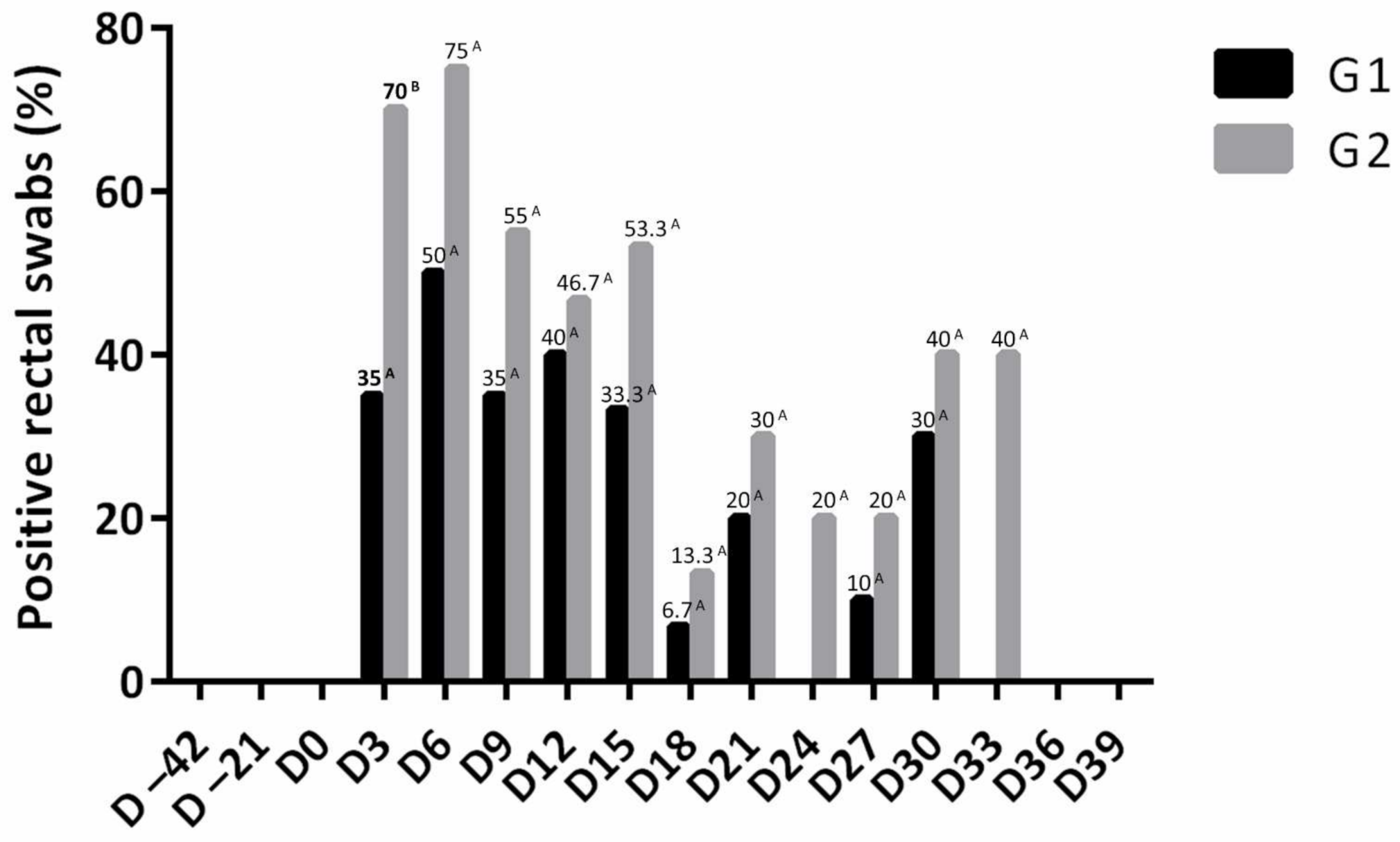
3.5. Bacteriology and Necroscopic Evaluation
Salmonella Isolation from Fecal Swabs and Organ Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barrow, P.A.; Methner, U. Salmonella in Domestic Animals, 2nd ed.; CAB International: London, UK, 2013; 547p. [Google Scholar]
- Issenhuth-Jeanjean, S.; Roggentin, P.; Mikoleit, M.; Guibourdenche, M.; de Pinna, E.; Nair, S.; Fields, P.I.; Weill, F.X. Supplement 2008-2010 (no. 48) to the White-Kauffmann-Le Minor scheme. Res. Microbiol. 2014, 165, 526–530. [Google Scholar] [CrossRef]
- Griffith, R.W.; Carlson, S.A.; Krull, A.C. Salmonellosis. In Disease of Swine; Zimmerman, J.J., Karrike, L.A., Ramirez, A.A., Schwartz, K.J., Stevenson, G.W., Zhang, J., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2019; pp. 912–925. ISBN 9781119350903. [Google Scholar]
- Demirbilek, S.K. Salmonellosis in Animals. In Salmonella—A Re-Emerging Pathogen; Mascellino, M.T., Ed.; IntechOpen: Budapest, Hungary, December 2017. [Google Scholar] [CrossRef]
- Bersot, L.S.; Cavicchioli, V.Q.; Viana, C.; Burin, R.C.K.; Camargo, A.C.; Pinto, J.P.A.N.; Nero, L.A.; Destro, M.T. Prevalence, Antimicrobial Resistance, and Diversity of Salmonella along the Pig Production Chain in Southern Brazil. Pathogens 2019, 8, 204. [Google Scholar] [CrossRef] [PubMed]
- Morningstar-Shaw, B.R.; Mackie, T.A.; Barker, D.K.; Palmer, E.A. Salmonella Serotypes Isolated from Animals and Related Sources January; CDC: Ames, IA, USA, 2016; pp. 1–4. [Google Scholar]
- Ferrari, R.G.; Rosario, D.K.A.; Cunha-Neto, A.; Mano, S.B.; Figueiredo, E.E.S.; Conte-Junior, C.A. Worldwide Epidemiology of Salmonella Serovars in Animal-Based Foods: A Meta-Analysis. Appl. Environ. Microbiol. 2019, 85, 1–21. [Google Scholar] [CrossRef]
- Arya, G.; Holtslander, R.; Robertson, J.; Yoshida, C.; Harris, J.; Parmley, J.; Nichani, A.; Johnson, R.; Poppe, C. Epidemiology, Pathogenesis, Genoserotyping, Antimicrobial Resistance, and Prevention and Control of Non-Typhoidal Salmonella Serovars. Curr. Clin. Microbiol. Rep. 2017, 4, 43–53. [Google Scholar] [CrossRef]
- Campos, J.; Mourão, J.; Peixe, L.; Antunes, P. Non-Typhoidal Salmonella in the Pig Production Chain: A Comprehensive Analysis of Its Impact on Human Health. Pathogens 2019, 8, 19. [Google Scholar] [CrossRef] [PubMed]
- Williams, H.; Jones, J.E.T. Observations on Experimental Oral Infection with Salmonella Dublin in Calves and Salmonella Choleraesuis in Pigs. Anim. Health Trust 1967, 93, 141–156. [Google Scholar] [CrossRef]
- Bonardi, S. Salmonella in the Pork Production Chain and Its Impact on Human Health in the European Union. Epidemiol. Infect. 2017, 145, 1513–1526. [Google Scholar] [CrossRef]
- Vidic, B.; Savic, S.; Prica, N.; Suvajdzic, L. Epizootiology and Control Measures for Salmonella in Pigs. Procedia Food Sci. 2015, 5, 312–315. [Google Scholar] [CrossRef]
- de Busser, E.V.; de Zutter, L.; Dewulf, J.; Houf, K.; Maes, D. Salmonella Control in Live Pigs and at Slaughter. Vet. J. 2013, 196, 20–27. [Google Scholar] [CrossRef]
- Martínez-Avilés, M.; Garrido-Estepa, M.; Álvarez, J.; de la Torre, A. Salmonella Surveillance Systems in Swine and Humans in Spain: A Review. Vet. Sci. 2019, 6, 20. [Google Scholar] [CrossRef]
- Patterson, S.K.; Kim, H.B.; Borewicz, K.; Isaacson, R.E. Towards an Understanding of Salmonella Enterica Serovar Typhimurium Persistence in Swine. Anim. Health Res. Rev. 2016, 17, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Eng, S.K.; Pusparajah, P.; Ab Mutalib, N.S.; Ser, H.L.; Chan, K.G.; Lee, L.H. Salmonella: A Review on Pathogenesis, Epidemiology and Antibiotic Resistance. Front. Life Sci. 2015, 8, 284–293. [Google Scholar] [CrossRef]
- Wales, A.D.; Davies, R.H. Salmonella Vaccination in Pigs: A Review. Zoonoses Public Health 2017, 64, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Boyen, F.; Haesebrouck, F.; Maes, D.; van Immerseel, F.; Ducatelle, R.; Pasmans, F. Non-Typhoidal Salmonella Infections in Pigs: A Closer Look at Epidemiology, Pathogenesis and Control. Vet. Microbiol. 2008, 130, 1–19. [Google Scholar] [CrossRef]
- Peeters, L.; Dewulf, J.; Boyen, F.; Brossé, C.; Vandersmissen, T.; Rasschaert, G.; Heyndrickx, M.; Cargnel, M.; Mattheus, W.; Pasmans, F.; et al. Evaluation of Group Vaccination of Sows and Gilts against Salmonella Typhimurium with an Attenuated Vaccine in Subclinically Infected Pig Herds. Prev. Vet. Med. 2020, 182, 104884. [Google Scholar] [CrossRef]
- Takaya, A.; Yamamoto, T.; Tokoyoda, K. Humoral Immunity vs. Salmonella. Front. Immunol. 2020, 10, 1–7. [Google Scholar] [CrossRef]
- Mastroeni, P.; Ménager, N. Development of Acquired Immunity to Salmonella. J. Med. Microbiol. 2003, 52, 453–459. [Google Scholar] [CrossRef]
- de la Cruz, M.L.; Conrado, I.; Nault, A.; Perez, A.; Dominguez, L.; Alvarez, J. Vaccination as a Control Strategy against Salmonella Infection in Pigs: A Systematic Review and Meta-Analysis of the Literature. Res. Vet. Sci. 2017, 114, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Wood, R.L.; Rose, R.; Coe, N.E.; Ferris, K.E. Experimental Establishment of Persistent Infection in Swine with a Zoonotic Strain of Salmonella Newport. Am. J. Vet. Res. 1991, 52, 813–819. [Google Scholar]
- Oliveira, L.G.; Carvalho, L.F.O.S.; Masson, G.C.I.H.; Feliciano, M.A.R. Experimental Infection by Salmonella enterica ssp. enterica serovar Panama and Tentative of Nose-to-Nose Transmission in Weaned Pigs. Arq. Bras. Med. Vet. Zootec. 2010, 62, 1340–1347. [Google Scholar] [CrossRef]
- Bauer, A.W.; Kirby, W.M.M.; Sherris, J.C.; Turck, M. Antibiotic Susceptibility Testing by a Standardized Single Disk Method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Abmm, D.; Tamma, D.; Kirn, J.; Cullen, S.K. Clinical and Laboratory Standards Institute (CLSI). In Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI supplement M100: Wayne, PA, USA, 2020. [Google Scholar]
- Feitosa, F.L.F. Semiologia Veterinária: A Arte Do Diagnóstico, 4th ed.; Roca: Rio de Janeiro, Brazil, 2020; 704p. [Google Scholar]
- Thrall, M.A.; Weiser, G.; Allison, R.W.; Campbell, T.W. Hematologia e Bioquímica Clínica Veterinária, 2nd ed.; Roca: Rio de Janeiro, Brazil, 2015; 678p. [Google Scholar]
- Triola, M.F. Introdução à Estatística, 12th ed.; LTC: Rio de Janeiro, Brazil, 2017; 812p. [Google Scholar]
- Ruggeri, J.; Pesciaroli, M.; Foresti, F.; Giacomini, E.; Lazzaro, M.; Ossiprandi, M.C.; Corradi, A.; Lombardi, G.; Pasquali, P.; Alborali, G.L. Inactivated Salmonella enterica Serovar Typhimurium Monophasic Variant (S. Typhimurium 1,4,[5],12:I-) in Sows Is Effective to Control Infection in Piglets under Field Condition. Vet. Microbiol. 2015, 180, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Alborali, G.L.; Ruggeri, J.; Pesciaroli, M.; Martinelli, N.; Chirullo, B.; Ammendola, S.; Battistoni, A.; Ossiprandi, M.C.; Corradi, A.; Pasquali, P. Prime-Boost Vaccination with Attenuated Salmonella Typhimurium ΔznuABC and Inactivated Salmonella Choleraesuis Is Protective against Salmonella Choleraesuis Challenge Infection in Piglets. BMC Vet. Res. 2017, 13, 284. [Google Scholar] [CrossRef]
- Gradassi, M.; Pesciaroli, M.; Martinelli, N.; Ruggeri, J.; Petrucci, P.; Hassan, W.H.; Raffatellu, M.; Scaglione, F.E.; Ammendola, S.; Battistoni, A.; et al. Attenuated Salmonella enterica serovar Typhimurium lacking the ZnuABC transporter: An efficacious orally-administered mucosal vaccine against salmonellosis in pigs. Vaccine 2013, 31, 3695–3701. [Google Scholar] [CrossRef]
- Farzan, A.; Friendship, R.M. A Clinical Field Trial to Evaluate the Efficacy of Vaccination in Controlling Salmonella Infection and the Association of Salmonella-Shedding and Weight Gain in Pigs. Can. J. Vet. Res. 2010, 74, 258–263. [Google Scholar]
- Schwarz, P.; Kich, J.D.; Kolb, J.; Cardoso, M. Use of an Avirulent Live Salmonella Choleraesuis Vaccine to Reduce the Prevalence of Salmonella Carrier Pigs at Slaughter. Vet. Rec. 2011, 169, 553. [Google Scholar] [CrossRef]
- Murtaugh, M.P. Advances in Swine Immunology Help Move Vaccine Technology Forward. Vet. Immunol. Immunopathol. 2014, 159, 202–207. [Google Scholar] [CrossRef]
- Mostaço, G.M.; Miranda, K.O.S.; Condotta, I.C.F.S.; Salgado, D.D. Determination of Piglets’ Rectal Temperature and Respiratory Rate through Skin Surface Temperature Under Climatic Chamber Conditions. Eng. Agríc. 2015, 35, 979–989. [Google Scholar] [CrossRef]
- Burdick Sanchez, N.C.; Carroll, J.A.; Corley, J.R.; Broadway, P.R.; Callaway, T.R. Changes in the Hematological Variables in Pigs Supplemented With Yeast Cell Wall in Response to a Salmonella Challenge in Weaned Pigs. Front. Vet. Sci. 2019, 6, 1–13. [Google Scholar] [CrossRef]
- Perri, A.M.; O’Sullivan, T.L.; Harding, J.C.S.; Wood, R.D.; Friendship, R.M. Hematology and Biochemistry Reference Intervals for Ontario Commercial Nursing Pigs Close to the Time of Weaning. Can. Vet. J. 2017, 58, 371–376. [Google Scholar]
- Rostagno, M.H.; Eicher, S.D.; Lay, D.C., Jr. Immunological, Physiological, and Behavioral Effects of Salmonella enterica Carriage and Shedding in Experimentally Infected Finishing Pigs. Foodborne Pathog. Dis. 2011, 8, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Arguello, H.; Carvajal, A.; Naharro, G.; Rubio, P. Evaluation of Protection Conferred by a Salmonella Typhimurium Inactivated Vaccine in Salmonella-Infected Finishing Pig Farms. Comp. Immunol. Microbiol. Infect. Dis. 2013, 36, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Wales, A.D.; Cook, A.J.C.; Davies, R.H. Producing Salmonella-Free Pigs: A Review Focusing on Interventions at Weaning. Vet. Rec. 2011, 168, 267–276. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Rectal Temperature (°C) 1 | Pigs Showing Signs of Diarrhea (%) 2 | Weight (Kg) 1 | ||||||
|---|---|---|---|---|---|---|---|---|
| Moments | Groups | Groups | Groups | |||||
| G1 | G2 | G1 | G2 | G1 | G2 | |||
| D–42 | 38.2 ± 0.34 A | 38.1 ± 0.55 A | 0.0 | (0/20) | 0.0 | (0/20) | 6.43 ± 0.45 A | 6.61 ± 1.29 A |
| D–21 | 38.9 ± 0.43 A | 38.6 ± 0.33 A | 5.00 A | (1/20) | 0.00 A | (0/20) | 10.7 ± 1.51 A | 10.9 ± 1.77 A |
| D0 | 38.5 ± 0.69 A | 38.6 ± 0.57 A | 0.0 | (0/20) | 0.0 | (0/20) | 17.0 ± 3.14 A | 17.5 ± 2.37 A |
| D3 | 38.4 ± 0.49 A | 38.8 ± 0.72 A | 60.0 A | (12/20) | 80.0 A | (16/20) | 17.8 ± 3.38 A | 18.4 ± 3.01 A |
| D6 | 38.7 ± 0.30 A | 39.3 ± 0.50 B | 35.0 A | (7/20) | 65.0 B | (13/20) | 19.7 ± 3.54 A | 20.1 ± 3.35 A |
| D9 | 38.7 ± 0.35 A | 38.7 ± 0.40 A | 45.0 A | (9/20) | 55.0 A | (11/20) | 21.8 ± 3.94 A | 21.8 ± 3.55 A |
| D12 | 38.8 ± 0.38 A | 38.7 ± 0.29 A | 20.0 A | (3/15) | 46.7 A | (7/15) | 24.5 ± 3.66 A | 24.8 ± 3.51 A |
| D15 | 38.6 ± 0.24 A | 38.7 ± 0.36 A | 6.67 A | (1/15) | 6.67 A | (1/15) | 26.3 ± 3.74 A | 26.2 ± 3.82 A |
| D18 | 38.9 ± 0.23 A | 38.9 ± 0.34 A | 13.3 A | (2/15) | 0.00 A | (0/15) | 27.7 ± 3.60 A | 26.7 ± 3.94 A |
| D21 | 38.7 ± 0.22 A | 39.0 ± 0.17 A | 60.0 A | (6/10) | 30.0 A | (3/10) | 29.0 ± 4.39 A | 28.8 ± 3.84 A |
| D24 | 38.9 ± 0.38 A | 38.8 ± 0.40 A | 0.00 A | (0/10) | 10.0 A | (1/10) | 29.7 ± 5.38 A | 29.0 ± 4.02 A |
| D27 | 38.1 ± 0.48 A | 38.1 ± 0.40 A | 30.0 A | (3/10) | 30.0 A | (3/10) | 30.8 ± 5.37 A | 30.4 ± 4.14 A |
| D30 | 38.2 ± 0.40 A | 38.2 ± 0.37 A | 20.0 A | (2/10) | 10.0 A | (1/10) | 32.3 ± 5.42 A | 31.7 ± 4.62 A |
| D33 | 38.6 ± 0.47 A | 39.0 ± 0.36 A | 0.00 A | (0/5) | 40.0 A | (2/5) | 35.7 ± 7.15 A | 34.2 ± 6.43 A |
| D36 | 38.2 ± 0.54 A | 38.5 ± 0.22 A | 0.0 | (0/5) | 0.0 | (0/5) | 37.2 ± 7.16 A | 35.3 ± 5.65 A |
| D39 | 37.9 ± 0.61 A | 38.0 ± 0.39 A | 0.0 | (0/5) | 0.0 | (0/5) | 38.6 ± 7.53 A | 36.2 ± 5.87 A |
| Groups | Organ | Timepoint | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| D10 | D20 | D30 | D40 | Total | |||||||
| +/Total | % | +/Total | % | +/Total | % | +/Total | % | +/Total | % | ||
| G1 | Lung | 2/5 | 40.0 | 1/5 | 20.0 | 1/5 | 20.0 | 0/4 | 0.0 | 4/19 | 21.0 |
| Liver | 0/5 | 0.0 | 0/5 | 0.0 | 0/5 | 0.0 | 0/4 | 0.0 | 0/19 | 0.0 | |
| Spleen | 0/5 | 0.0 | 0/5 | 0.0 | 0/5 | 0.0 | 0/4 | 0.0 | 0/19 | 0.0 | |
| Mesentery lymph node | 2/5 | 40.0 | 0/5 | 0.0 | 1/5 | 20.0 | 0/4 | 0.0 | 3/19 | 15.8 | |
| Ileum | 4/5 | 80.0 | 3/5 | 60.0 | 3/5 | 60.0 | 3/4 | 75.0 | 13/19 | 68.4 | |
| Cecum | 5/5 | 100 | 5/5 | 100 | 5/5 | 100 | 3/4 | 75.0 | 18/19 | 94.7 | |
| Ileocolic lymph node | 4/5 | 80.0 | 1/5 | 20.0 | 2/5 | 40.0 | 0/4 | 0.0 | 7/19 | 36.8 | |
| Total | 17/35 | 48.6 A | 10/35 | 28.6 A | 12/35 | 34.3 A | 6/28 | 21.4 | 45/133 | 33.8 A | |
| G2 | Lung | 1/5 | 20.0 | 1/5 | 20.0 | 2/5 | 40.0 | 1/5 | 20.0 | 5/20 | 25.0 |
| Liver | 1/5 | 20.0 | 0/5 | 0.0 | 0/5 | 0.0 | 0/5 | 0.0 | 1/20 | 5.00 | |
| Spleen | 0/5 | 0.0 | 0/5 | 0.0 | 0/5 | 0.0 | 0/5 | 0.0 | 0/20 | 0.0 | |
| Mesentery lymph node | 4/5 | 80.0 | 1/5 | 20.0 | 1/5 | 20.0 | 0/5 | 0.0 | 6/20 | 30.0 | |
| Ileum | 5/5 | 100 | 3/5 | 60.0 | 4/5 | 80.0 | 3/5 | 60.0 | 15/20 | 75.0 | |
| Cecum | 5/5 | 100 | 5/5 | 100 | 5/5 | 100 | 4/5 | 80.0 | 19/20 | 95.0 | |
| Ileo-colic lymph node | 5/5 | 100 | 2/5 | 40.0 | 0/5 | 0.0 | 1/5 | 20.0 | 8/20 | 40.0 | |
| Total | 21/35 | 60.0 A | 12/35 | 34.3 A | 12/35 | 34.3 A | 9/35 | 25.7 | 54/140 | 38.6 A | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moura, E.A.G.d.O.; Silva, D.G.d.; Turco, C.H.; Sanches, T.V.C.; Storino, G.Y.; Almeida, H.M.d.S.; Mechler-Dreibi, M.L.; Rabelo, I.P.; Sonalio, K.; Oliveira, L.G.d. Salmonella Bacterin Vaccination Decreases Shedding and Colonization of Salmonella Typhimurium in Pigs. Microorganisms 2021, 9, 1163. https://doi.org/10.3390/microorganisms9061163
Moura EAGdO, Silva DGd, Turco CH, Sanches TVC, Storino GY, Almeida HMdS, Mechler-Dreibi ML, Rabelo IP, Sonalio K, Oliveira LGd. Salmonella Bacterin Vaccination Decreases Shedding and Colonization of Salmonella Typhimurium in Pigs. Microorganisms. 2021; 9(6):1163. https://doi.org/10.3390/microorganisms9061163
Chicago/Turabian StyleMoura, Eduarda Alexandra Gonçalves de Oliveira, Daniela Gomes da Silva, Caio Henrique Turco, Thainara Vitoria Carnevalli Sanches, Gabriel Yuri Storino, Henrique Meiroz de Souza Almeida, Marina Lopes Mechler-Dreibi, Isabela Peixoto Rabelo, Karina Sonalio, and Luís Guilherme de Oliveira. 2021. "Salmonella Bacterin Vaccination Decreases Shedding and Colonization of Salmonella Typhimurium in Pigs" Microorganisms 9, no. 6: 1163. https://doi.org/10.3390/microorganisms9061163
APA StyleMoura, E. A. G. d. O., Silva, D. G. d., Turco, C. H., Sanches, T. V. C., Storino, G. Y., Almeida, H. M. d. S., Mechler-Dreibi, M. L., Rabelo, I. P., Sonalio, K., & Oliveira, L. G. d. (2021). Salmonella Bacterin Vaccination Decreases Shedding and Colonization of Salmonella Typhimurium in Pigs. Microorganisms, 9(6), 1163. https://doi.org/10.3390/microorganisms9061163