
Feedborne Salmonella enterica Serovar Jerusalem Outbreak in Different Organic Poultry Flocks in Switzerland and Italy Linked to Soya Expeller

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates
2.2. Whole-Genome Sequencing Analysis and Bioinformatics
2.3. Antimicrobial Susceptibility Testing
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tindall, B.J.; Grimont, P.; Garrity, G.; Euzéby, J.P. Nomenclature and taxonomy of the genus Salmonella. Int. J. Syst. Evol. Microbiol. 2005, 55, 521–524. [Google Scholar] [CrossRef] [Green Version]
- Grimont, P.; Weill, F.-X. Antigenic Formulae of the Salmonella Servovars: WHO Collaborating Centre for Reference and Research on Salmonella, 9th ed.; Institute Pasteur: Paris, France, 2007. [Google Scholar]
- European Food Safety Authority and European Centre for Disease Prevention and Control (EFSA and ECDC). The European Union One Health 2019 Zoonoses Report. EFSA J. 2021, 19, e05926. [Google Scholar] [CrossRef]
- Murray, C. Salmonellae in the environement. Rev. Sci. Tech. 1991, 10, 765–785. [Google Scholar] [CrossRef]
- Hoop, R.K. The Swiss control programme for Salmonella enteritidis in laying hens: Experiences and problems. Rev. Sci. Tech. l’OIE 1997, 16, 885–890. [Google Scholar] [CrossRef] [PubMed]
- Anonymous. Tierseuchenverordnung SR 916.401, Art. 255–261. 2021. Available online: https://www.fedlex.admin.ch/eli/cc/1995/3716_3716_3716/de (accessed on 22 June 2021).
- Anonymous. Technische Weisungen über die Entnahme von Proben und deren Untersuchung auf Salmonella-Infektion des Geflügels und über das Vorgehen im Verdachts- und Seuchenfall vom 01.05.2021. 2021. Available online: https://www.blv.admin.ch/blv/de/home/tiere/rechts--und-vollzugsgrundlagen/hilfsmittel-und-vollzugsgrundlagen/technische-weisungen.html (accessed on 4 June 2021).
- Maciorowski, K.G.; Herrera, P.; Kundinger, M.M.; Ricke, S.C. Animal Feed Production and Contamination by Foodborne Salmonella. J. Verbrauch. Lebensm. 2006, 1, 197–209. [Google Scholar] [CrossRef]
- Crump, J.A.; Griffin, P.M.; Angulo, F.J. Bacterial Contamination of Animal Feed and Its Relationship to Human Foodborne Illness. Clin. Infect. Dis. 2002, 35, 859–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, F.T. A review of practical Salmonella control measures in animal feed. J. Appl. Poult. Res. 2011, 20, 102–113. [Google Scholar] [CrossRef]
- Schleifer, J.H.; Juven, B.J.; Beard, C.W.; Cox, N.A. The Susceptibility of Chicks to Salmonella montevideo in Artificially Contaminated Poultry Feed. Avian Dis. 1984, 28, 497. [Google Scholar] [CrossRef] [PubMed]
- Jones, F.; Axtell, R.; Tarver, F.; Rives, D.; Scheideler, S.; Wineland, M. Environmental factors contributing to Salmonella colonization of chickens. In Colonization Control of Human Bacterial Enteropathologens in Poultry; Elsevier: Amsterdam, The Netherlands, 1991; pp. 3–21. [Google Scholar]
- Thakur, S.; Brake, J.; Keelara, S.; Zou, M.; Susick, E. Farm and environmental distribution of Campylobacter and Salmonella in broiler flocks. Res. Veter. Sci. 2013, 94, 33–42. [Google Scholar] [CrossRef]
- Rodriguez, A.; Pangloli, P.; Richards, H.A.; Mount, J.R.; Draughon, F.A. Prevalence of Salmonella in Diverse Environmental Farm Samples. J. Food Prot. 2006, 69, 2576–2580. [Google Scholar] [CrossRef]
- Byrd, J.A.; Corrier, D.E.; Deloach, J.R.; Nisbet, D.J.; Stanker, L.H. Horizontal Transmission of Salmonella typhimurium in Broiler Chicks. J. Appl. Poult. Res. 1998, 7, 75–80. [Google Scholar] [CrossRef]
- Shirota, K.; Katoh, H.; Murase, T.; Ito, T.; Otsuki, K. Monitoring of Layer Feed and Eggs for Salmonella in Eastern Japan between 1993 and 1998. J. Food Prot. 2001, 64, 734–737. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, M.A.; Bains, B.S. Dissemination of Salmonella Serotypes from Raw Feed Ingredients to Chicken Carcases. Poult. Sci. 1976, 55, 957–960. [Google Scholar] [CrossRef] [PubMed]
- Jones, F.T.; Axtell, R.C.; Rives, D.V.; Scheideler, S.E.; Tarver, F.R.; Walker, R.L.; Wineland, M.J. A Survey of Salmonella Contamination in Modern Broiler Production. J. Food Prot. 1991, 54, 502–507. [Google Scholar] [CrossRef] [PubMed]
- Veldman, A.; Vahl, H.; Borggreve, G.J.; Fuller, D.C. A survey of the incidence of Salmonella species and Enterobacteriaceae in poultry feeds and feed components. Veter. Rec. 1995, 136, 169–172. [Google Scholar] [CrossRef]
- Magossi, G.; Cernicchiaro, N.; Dritz, S.; Houser, T.; Woodworth, J.; Jones, C.; Trinetta, V. Evaluation of Salmonella presence in selected United States feed mills. MicrobiologyOpen 2019, 8, e00711. [Google Scholar] [CrossRef]
- Anonymous. Verordnung über die biologische Landwirtschaft und die Kennzeichnung Biologisch Produzierter Erzeugnisse und Lebensmittel (Bio-Verordnung) Art. 16a 7. 2021. Available online: https://www.fedlex.admin.ch/eli/oc/2004/734/de (accessed on 4 June 2021).
- RASFF notification 2020.3066. 2020. Available online: https://webgate.ec.europa.eu/rasff-window/screen/notification/435586 (accessed on 4 June 2021).
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.; Dvorkin, M.; Kulikov, A.; Lesin, V.M.; Nikolenko, S.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Shovill. 2019. Available online: https://github.com/tseemann/shovill (accessed on 4 June 2021).
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef] [PubMed]
- Davis, S.; Pettengill, J.B.; Luo, Y.; Payne, J.; Shpuntoff, A.; Rand, H.; Strain, E. CFSAN SNP Pipeline: An automated method for constructing SNP matrices from next-generation sequence data. PeerJ Comput. Sci. 2015, 1, e20. [Google Scholar] [CrossRef] [Green Version]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Treangen, T.J.; Ondov, B.D.; Koren, S.; Phillippy, A.M. The Harvest suite for rapid core-genome alignment and visualization of thousands of intraspecific microbial genomes. Genome Biol. 2014, 15, 524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CLSI, M100 Performance Standards for Antimicrobial Susceptibility Testing, 31st ed.; Clinical and Laboratory Standards Institute: Annapolis Junction, MD, USA, 2009.
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 23, 127–128. [Google Scholar] [CrossRef]
- Pedersen, K.; Sørensen, G.; Szabo, I.; Hächler, H.; Le Hello, S. Repeated isolation of Salmonella enterica Goverdhan, a very rare serovar, from Danish poultry surveillance samples. Veter. Microbiol. 2014, 174, 596–599. [Google Scholar] [CrossRef]
- Gallati, C.; Stephan, R.; Hächler, H.; Malorny, B.; Schroeter, A.; Nüesch-Inderbinen, M. Characterization of Salmonella enterica Subsp. enterica Serovar 4,[5],12:i:- Clones Isolated from Human and Other Sources in Switzerland between 2007 and 2011. Foodborne Pathog. Dis. 2013, 10, 549–554. [Google Scholar] [CrossRef]
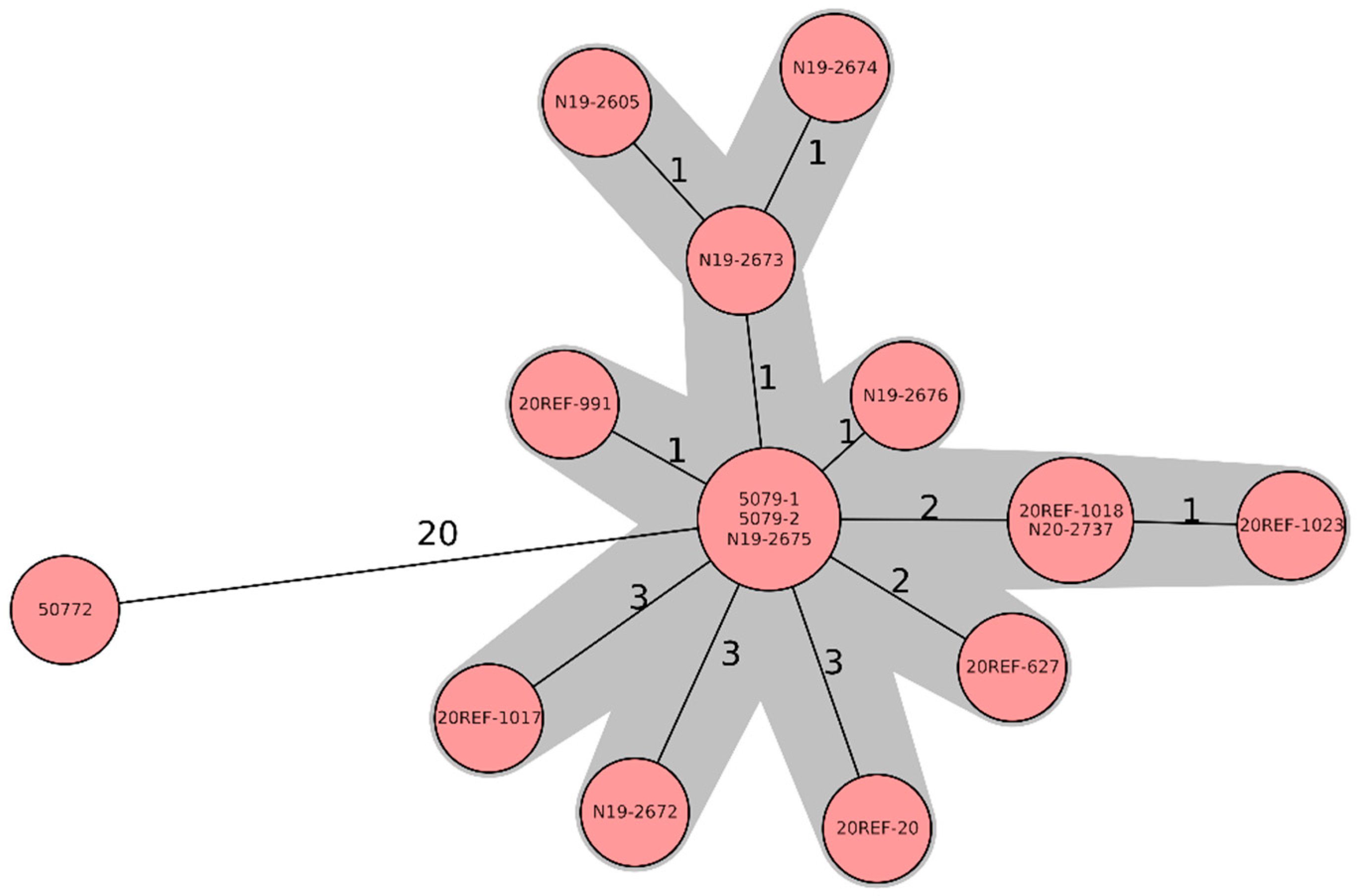

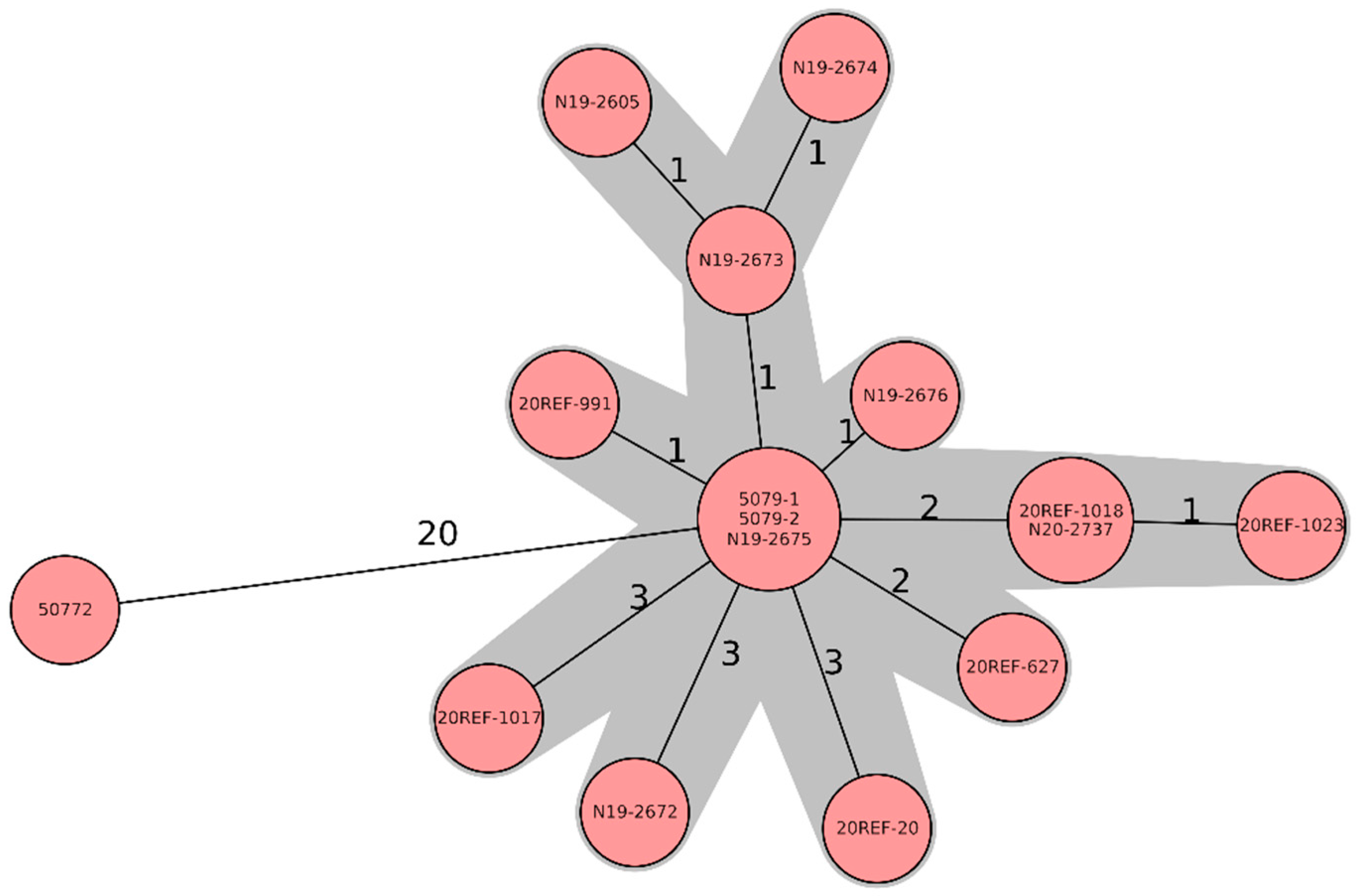{kind=link}
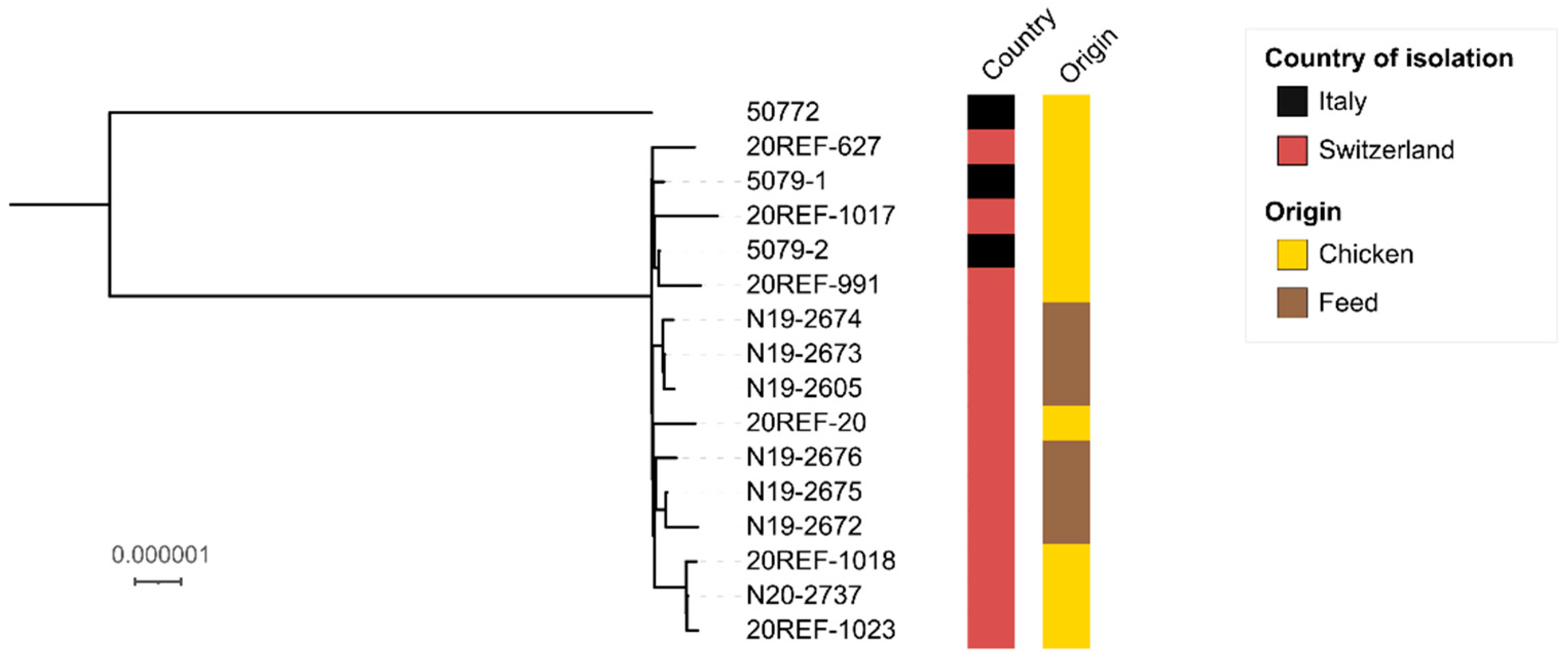
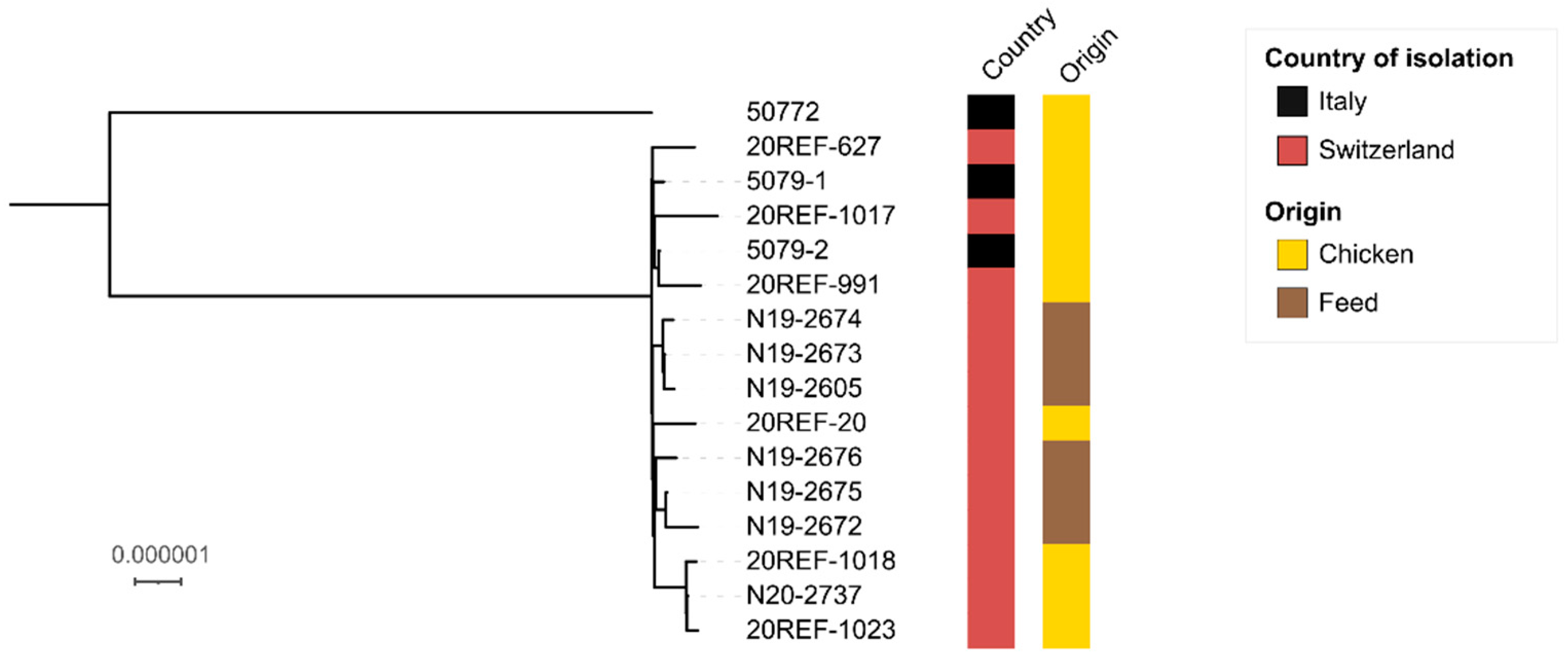{kind=link}
| Strain ID | Sample Type | Host | Production Type | Date of Isolation | Swiss Canton/Italian Region |
|---|---|---|---|---|---|
| N19-2605 | soya cake | n.a. | organic | 30 October 2019 | n.a. |
| N19-2672 | soya cake | n.a. | organic | 5 November 2019 | n.a. |
| N19-2673 | soya cake | n.a. | organic | 5 November 2019 | n.a. |
| N19-2674 | soya cake | n.a. | organic | 5 November 2019 | n.a. |
| N19-2675 | soya cake | n.a. | organic | 5 November 2019 | n.a. |
| N19-2676 | soya cake | n.a. | organic | 5 November 2019 | n.a. |
| N20-2737 | Feces | layer breeder | organic | 21 December 2020 | ZH |
| 20REF-20 | boot sock | laying hen | organic | 12 January 2020 | ZH |
| 20REF-627 | boot sock | laying hen | organic | 29 September 2020 | SG |
| 20REF-991 | Dust | laying hen | organic | 10 November 2020 | SO |
| 20REF-1017 | boot sock | laying hen | organic | 16 December 2020 | ZG |
| 20REF-1018 | Dust | laying hen | organic | 15 December 2020 | AG |
| 20REF-1023 | boot sock | laying hen | organic | 18 December 2020 | AG |
| 5079_1 | boot sock | broiler | organic | 25 January 2021 | Marche |
| 5079_2 | boot sock | broiler | organic | 25 January 2021 | Marche |
| 50772 | poultry manure from composting plant | poultry | unknown | 3 November 2003 | Marche |
| Strain | Year of Isolation | Origin | Nreads a | Coverage | Length (bp) | Contigs | N50 | L50 | N Genes | Accession No. |
|---|---|---|---|---|---|---|---|---|---|---|
| N19-2605 | 2019 | Feed | 966,373 | 61 | 4,782,067 | 40 | 334,862 | 5 | 4435 | JAFJZF000000000 |
| N19-2672 | 2019 | Feed | 887,841 | 56 | 4,781,041 | 35 | 402,221 | 4 | 4432 | JAFJZG000000000 |
| N19-2673 | 2019 | Feed | 957,477 | 61 | 4,780,566 | 34 | 348,029 | 5 | 4434 | JAFJZH000000000 |
| N19-2674 | 2019 | Feed | 982,883 | 62 | 4,780,191 | 41 | 347,991 | 6 | 4437 | JAFJZI000000000 |
| N19-2675 | 2019 | Feed | 992,783 | 63 | 4,781,953 | 36 | 348,407 | 5 | 4432 | JAFJZJ000000000 |
| N19-2676 | 2019 | Feed | 802,128 | 51 | 4,779,956 | 34 | 347,991 | 5 | 4433 | JAFJZK000000000 |
| N20-2737 | 2019 | Chicken | 1,070,707 | 68 | 4,787,949 | 40 | 334,862 | 5 | 4439 | JAFJZL000000000 |
| 20REF-20 | 2020 | Chicken | 891,287 | 56 | 4,781,073 | 38 | 348,023 | 5 | 4434 | JAFJZC000000000 |
| 20REF-627 | 2020 | Chicken | 695,228 | 44 | 4,778,515 | 40 | 348,023 | 6 | 4435 | JAFJZD000000000 |
| 20REF-991 | 2020 | Chicken | 919,591 | 58 | 4,779,174 | 43 | 325,448 | 6 | 4438 | JAFJZE000000000 |
| 20REF-1017 | 2020 | Chicken | 1,624,592 | 102 | 4,774,988 | 40 | 300,861 | 6 | 4435 | JAFJYZ000000000 |
| 20REF-1018 | 2020 | Chicken | 1,263,023 | 80 | 4,780,243 | 36 | 348,254 | 5 | 4434 | JAFJZA000000000 |
| 20REF-1023 | 2020 | Chicken | 1,187,079 | 75 | 4,780,375 | 38 | 348,393 | 5 | 4437 | JAFJYY000000000 |
| 5079-1 | 2021 | Chicken | 921,944 | 58 | 4,779,660 | 36 | 347,885 | 5 | 4430 | JAGYFQ000000000 |
| 5079-2 | 2021 | Chicken | 1,024,829 | 64 | 4,773,619 | 39 | 347,983 | 6 | 4434 | JAGYFP000000000 |
| 50772 | 2003 | Chicken | 907,688 | 58 | 4,725,864 | 39 | 287,235 | 6 | 4377 | JAGYFN000000000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Horlbog, J.A.; Stephan, R.; Stevens, M.J.A.; Overesch, G.; Kittl, S.; Napoleoni, M.; Silenzi, V.; Nüesch-Inderbinen, M.; Albini, S. Feedborne Salmonella enterica Serovar Jerusalem Outbreak in Different Organic Poultry Flocks in Switzerland and Italy Linked to Soya Expeller. Microorganisms 2021, 9, 1367. https://doi.org/10.3390/microorganisms9071367
Horlbog JA, Stephan R, Stevens MJA, Overesch G, Kittl S, Napoleoni M, Silenzi V, Nüesch-Inderbinen M, Albini S. Feedborne Salmonella enterica Serovar Jerusalem Outbreak in Different Organic Poultry Flocks in Switzerland and Italy Linked to Soya Expeller. Microorganisms. 2021; 9(7):1367. https://doi.org/10.3390/microorganisms9071367
Chicago/Turabian StyleHorlbog, Jule Anna, Roger Stephan, Marc J. A. Stevens, Gudrun Overesch, Sonja Kittl, Maira Napoleoni, Valentina Silenzi, Magdalena Nüesch-Inderbinen, and Sarah Albini. 2021. "Feedborne Salmonella enterica Serovar Jerusalem Outbreak in Different Organic Poultry Flocks in Switzerland and Italy Linked to Soya Expeller" Microorganisms 9, no. 7: 1367. https://doi.org/10.3390/microorganisms9071367
APA StyleHorlbog, J. A., Stephan, R., Stevens, M. J. A., Overesch, G., Kittl, S., Napoleoni, M., Silenzi, V., Nüesch-Inderbinen, M., & Albini, S. (2021). Feedborne Salmonella enterica Serovar Jerusalem Outbreak in Different Organic Poultry Flocks in Switzerland and Italy Linked to Soya Expeller. Microorganisms, 9(7), 1367. https://doi.org/10.3390/microorganisms9071367