
Effects of a Bioprocessed Soybean Meal Ingredient on the Intestinal Microbiota of Hybrid Striped Bass, Morone chrysops x M. saxatilis

 ,
, 
 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Diet Formulations
2.2. Feeding Trial
2.3. Microbial DNA Isolation and PCR Amplification
2.4. Computational Analysis of PCR Generated 16S rRNA Amplicon Sequences
2.5. Statistical Analysis
2.6. Next-Generation Sequencing Data Accessibility
3. Results
3.1. Feeding Trial Performance
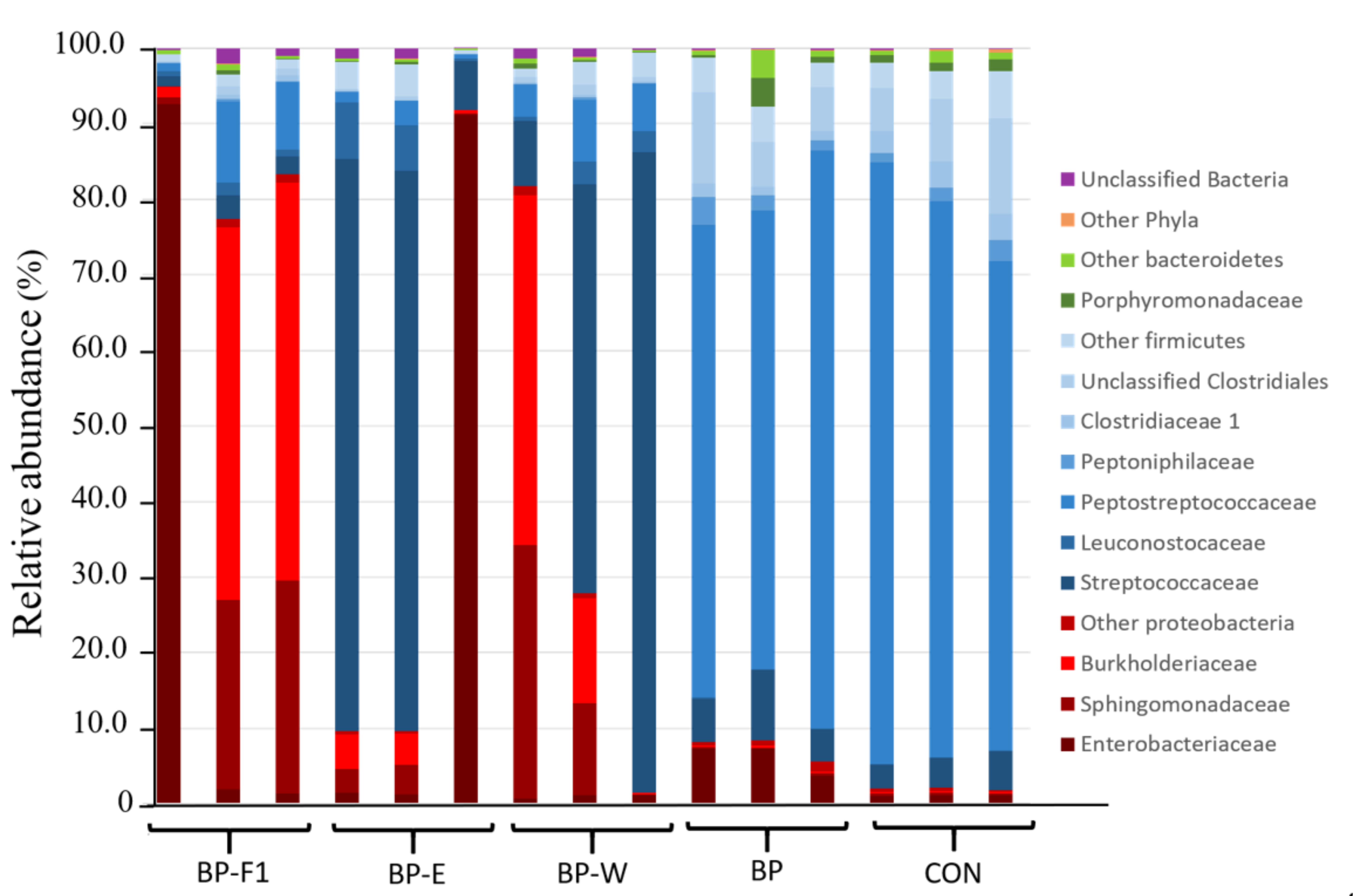
3.2. Taxonomic Composition Analysis
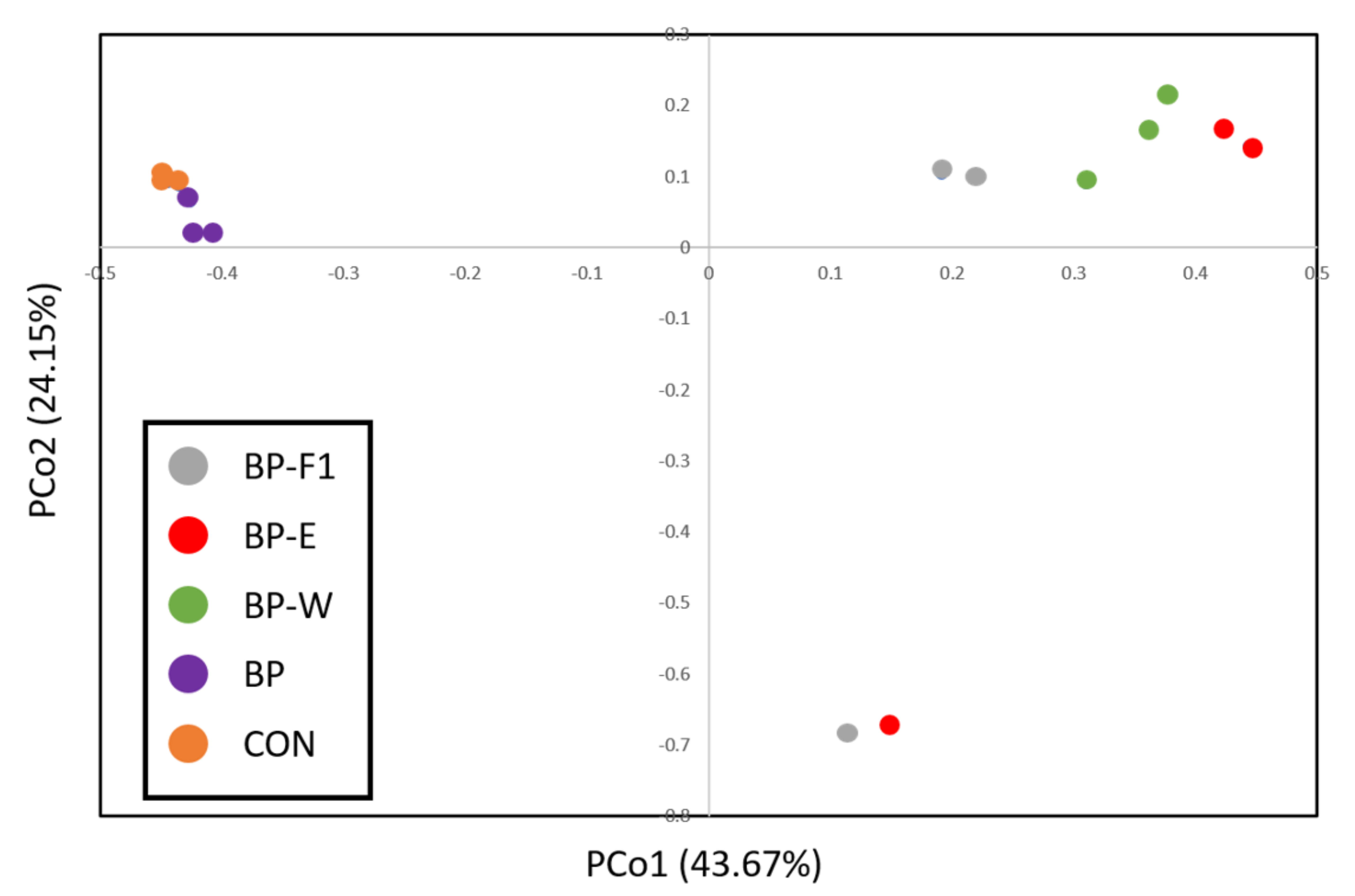
3.3. Alpha and Beta Diversity
3.4. OTU Composition Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Anguiano, M.; Pohlenz, C.; Buentello, A.; Gatlin, D.M., 3rd. The effects of prebiotics on the digestive enzymes and gut histomorphology of red drum (Sciaenops ocellatus) and hybrid striped bass (Morone chrysops × M. saxatilis). Br. J. Nutr. 2013, 109, 623–629. [Google Scholar] [CrossRef] [PubMed]
- Garber, A.F.; Sullivan, C.V. Selective breeding for the hybrid striped bass (Morone chrysops, Rafinesque × M. saxatilis, Walbaum) industry: Status and perspectives. Aquac. Res. 2006, 37, 319–338. [Google Scholar] [CrossRef]
- FAO 2016–2021. Cultured Aquatic Species Information Programme. In Morone Hybrid; Harrell, R.M., Ed.; FAO Fisheries Division: Rome, Italy, 2012. [Google Scholar]
- Quagrainie, K. Profitability of Hybrid Striped Bass Cage Aquaculture in the Midwest; Purdue University: West Lafayette, IN, USA, 2015. [Google Scholar]
- Lougheed, M.; Nelson, B. Hybrid Striped Bass Production, Markets and Marketing; Michigan State University, North Central Regional Aquaculture Center: East Lansing, MI, USA, 2001. [Google Scholar]
- Tacon, A.G.J.; Metian, M. Global overview on the use of fish meal and fish oil in industrially compounded aquafeeds: Trends and future prospects. Aquaculture 2008, 285, 146–158. [Google Scholar]
- Ytrestøyl, T.; Aas, T.S.; Åsgård, T. Utilisation of feed resources in production of Atlantic salmon (Salmo salar) in Norway. Aquaculture 2015, 448, 365–374. [Google Scholar]
- Asche, F.; Oglend, A.; Tveteras, S. Regime shifts in the fish meal/soybean meal price ratio. J. Agric. Econ. 2013, 64, 97–111. [Google Scholar] [CrossRef]
- Yaghoubi, M.; Mozanzadeh, M.T.; Marammazi, J.G.; Safari, O.; Gisbert, E. Dietary replacement of fish meal by soy products (soybean meal and isolated soy protein) in silvery-black porgy juveniles (Sparidentex hasta). Aquaculture 2016, 464, 50–59. [Google Scholar] [CrossRef]
- Sindelar, S.C. Utilization of Soybean Products as Fish-Meal Protein Replacements in Yellow Perch Perca flavescens Feeds. Master’s Thesis, South Dakota State University, Brookings, SD, USA, 2014. [Google Scholar]
- Perkins, E.G. Composition of soybeans and soybean products. In Practical Handbook of Soybean Processing and Utilization; Erickson, D.R., Ed.; AOCS Press: Champaign, IL, USA, 1995; pp. 9–28. [Google Scholar]
- Bruce, T.J.; Neiger, R.D.; Brown, M.L. Gut histology, immunology and the intestinal microbiota of rainbow trout, Oncorhynchus mykiss (Walbaum), fed process variants of soybean meal. Aquac. Res. 2018, 49, 492–504. [Google Scholar] [CrossRef]
- Lathrop, S.K.; Bloom, S.M.; Rao, S.M.; Nutsch, K.; Lio, C.W.; Santacruz, N.; Peterson, D.A.; Stappenbeck, T.S.; Hsieh, C.S. Peripheral education of the immune system by colonic commensal microbiota. Nature 2011, 478, 250–254. [Google Scholar] [CrossRef]
- Ley, R.E.; Lozupone, C.A.; Hamady, M.; Knight, R.; Gordon, J.I. Worlds within worlds: Evolution of the vertebrate gut microbiota. Nat. Rev. Microbiol. 2008, 6, 776–788. [Google Scholar] [CrossRef]
- Ciarlo, E.; Heinonen, T.; Herderschee, J.; Fenwick, C.; Mombelli, M.; Le Roy, D.; Roger, T. Impact of the microbial derived short chain fatty acid propionate on host susceptibility to bacterial and fungal infections in vivo. Sci. Rep. 2016, 6, 37944. [Google Scholar] [CrossRef]
- Gómez, G.D.; Balcázar, J.L. A review on the interactions between gut microbiota and innate immunity of fish. FEMS Immunol. Med. Microbiol. 2008, 52, 145–154. [Google Scholar] [CrossRef]
- Llewellyn, M.S.; Boutin, S.; Hoseinifar, S.H.; Derome, N. Teleost microbiomes: The state of the art in their characterization, manipulation and importance in aquaculture and fisheries. Front. Microbiol. 2014, 5, 207. [Google Scholar] [CrossRef]
- Talwar, C.; Nagar, S.; Lal, R.; Negi, R.K. Fish gut microbiome: Current approaches and future perspectives. Indian J. Microbiol. 2018, 58, 397–414. [Google Scholar] [CrossRef]
- Sullam, K.E.; Essinger, S.D.; Lozupone, C.A.; O’Connor, M.P.; Rosen, G.L.; Knight, R.; Kilham, S.S.; Russell, J.A. Environmental and ecological factors that shape the gut bacterial communities of fish: A meta-analysis. Mol. Ecol. 2012, 21, 3363–3378. [Google Scholar] [CrossRef]
- Liu, H.; Guo, X.; Gooneratne, R.; Lai, R.; Zeng, C.; Zhan, F.; Wang, W. The gut microbiome and degradation enzyme activity of wild freshwater fishes influenced by their trophic levels. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Uren Webster, T.M.; Rodriguez-Barreto, D.; Castaldo, G.; Gough, P.; Consuegra, S.; Garcia de Leaniz, C. Environmental plasticity and colonisation history in the Atlantic salmon microbiome: A translocation experiment. Mol. Ecol. 2020, 29, 886–898. [Google Scholar] [CrossRef] [PubMed]
- Egerton, S.; Wan, A.; Murphy, K.; Collins, F.; Ahern, G.; Sugrue, I.; Busca, K.; Egan, F.; Muller, N.; Whooley, J. Replacing fishmeal with plant protein in Atlantic salmon (Salmo salar) diets by supplementation with fish protein hydrolysate. Sci. Rep. 2020, 10, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Huyben, D.; Roehe, B.K.; Bekaert, M.; Ruyter, B.; Glencross, B. Dietary lipid: Protein ratio and n-3 long-chain polyunsaturated fatty acids alters the gut microbiome of Atlantic salmon under hypoxic and normoxic conditions. Front. Microbiol. 2020, 11, 3385. [Google Scholar] [CrossRef]
- Fogarty, C.; Burgess, C.M.; Cotter, P.D.; Cabrera-Rubio, R.; Whyte, P.; Smyth, C.; Bolton, D.J. Diversity and composition of the gut microbiota of Atlantic salmon (Salmo salar) farmed in Irish waters. J. Appl. Microbiol. 2019, 127, 648–657. [Google Scholar] [CrossRef]
- Gupta, S.; Fečkaninová, A.; Lokesh, J.; Koščová, J.; Sørensen, M.; Fernandes, J.; Kiron, V. Lactobacillus dominate in the intestine of Atlantic salmon fed dietary probiotics. Front. Microbiol. 2019, 9, 3247. [Google Scholar] [CrossRef]
- Brown, R.M.; Wiens, G.D.; Salinas, I. Analysis of the gut and gill microbiome of resistant and susceptible lines of rainbow trout (Oncorhynchus mykiss). Fish Shellfish. Immunol. 2019, 86, 497–506. [Google Scholar] [CrossRef]
- Giang, P.T.; Sakalli, S.; Fedorova, G.; Tilami, S.K.; Bakal, T.; Najmanova, L.; Grabicova, K.; Kolarova, J.; Sampels, S.; Zamaratskaia, G. Biomarker response, health indicators, and intestinal microbiome composition in wild brown trout (Salmo trutta m. fario L.) exposed to a sewage treatment plant effluent-dominated stream. Sci. Total Environ. 2018, 625, 1494–1509. [Google Scholar] [CrossRef]
- Lyons, P.; Turnbull, J.; Dawson, K.A.; Crumlish, M. Phylogenetic and functional characterization of the distal intestinal microbiome of rainbow trout Oncorhynchus mykiss from both farm and aquarium settings. J. Appl. Microbiol. 2017, 122, 347–363. [Google Scholar] [CrossRef]
- Michl, S.C.; Beyer, M.; Ratten, J.-M.; Hasler, M.; LaRoche, J.; Schulz, C. A diet-change modulates the previously established bacterial gut community in juvenile brown trout (Salmo trutta). Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Michl, S.C.; Ratten, J.-M.; Beyer, M.; Hasler, M.; LaRoche, J.; Schulz, C. The malleable gut microbiome of juvenile rainbow trout (Oncorhynchus mykiss): Diet-dependent shifts of bacterial community structures. PLoS ONE 2017, 12, e0177735. [Google Scholar] [CrossRef]
- Rimoldi, S.; Terova, G.; Ascione, C.; Giannico, R.; Brambilla, F. Next generation sequencing for gut microbiome characterization in rainbow trout (Oncorhynchus mykiss) fed animal by-product meals as an alternative to fishmeal protein sources. PLoS ONE 2018, 13, e0193652. [Google Scholar] [CrossRef]
- Valdés, N.; Gonzalez, A.; Garcia, V.; Tello, M. Analysis of the microbiome of rainbow trout (Oncorhynchus mykiss) exposed to the pathogen Flavobacterium psychrophilum 10094. Microbiol. Resour. Announc. 2020, 9. [Google Scholar] [CrossRef]
- Bledsoe, J.W.; Waldbieser, G.C.; Swanson, K.S.; Peterson, B.C.; Small, B.C. Comparison of channel catfish and blue catfish gut microbiota assemblages shows minimal effects of host genetics on microbial structure and inferred function. Front. Microbiol. 2018, 9, 1073. [Google Scholar] [CrossRef]
- Burgos, F.A.; Ray, C.L.; Arias, C.R. Bacterial diversity and community structure of the intestinal microbiome of Channel Catfish (Ictalurus punctatus) during ontogenesis. Syst. Appl. Microbiol. 2018, 41, 494–505. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, H.; Hu, K.; Jiao, L.; Zhao, M.; Yang, X.; Xia, L. Effect of Dietary supplementation of Lactobacillus casei YYL3 and L. plantarum YYL5 on growth, immune response and intestinal microbiota in channel catfish. Animals 2019, 9, 1005. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, D.; Refaey, M.M.; Xu, W. High spatial and temporal variations of microbial community along the southern catfish gastrointestinal tract: Insights into dynamic food digestion. Front. Microbiol. 2017, 8, 1531. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, D.; Refaey, M.M.; Xu, W.; Tang, R.; Li, L. Host age affects the development of southern catfish gut bacterial community divergent from that in the food and rearing water. Front. Microbiol. 2018, 9, 495. [Google Scholar] [CrossRef]
- Zhong, L.; Li, D.; Wang, M.; Chen, X.; Bian, W.; Zhu, G. Dynamics of the bacterial community in a channel catfish nursery pond with a cage–pond integration system. Can. J. Microbiol. 2018, 64, 954–967. [Google Scholar] [CrossRef]
- MacFarlane, R.D.; McLaughlin, J.J.; Bullock, G.L. Quantitative and qualitative studies of gut flora in striped bass from estuarine and coastal marine environments. J. Wildl. Dis. 1986, 22, 344–348. [Google Scholar] [CrossRef]
- Nedoluha, P.C.; Westhoff, D. Microbiology of striped bass grown in three aquaculture systems. Food Microbiol. 1997, 14, 255–264. [Google Scholar] [CrossRef]
- Nedoluha, P.C.; Westhoff, D. Microbiological analysis of striped bass (Morone saxatilis) grown in flow-through tanks. J. Food Prot. 1995, 58, 1363–1368. [Google Scholar] [CrossRef]
- Nedoluha, P.C.; Westhoff, D. Microbiological analysis of striped bass (Morone saxatilis) grown in a recirculating system (†). J. Food Prot. 1997, 60, 948–953. [Google Scholar] [CrossRef]
- Nedoluha, P.C.; Westhoff, D. Microbiological flora of aquacultured hybrid striped bass. J. Food Prot. 1993, 56, 1054–1060. [Google Scholar] [CrossRef]
- Aly, S.M.; Abd-El-Rahman, A.M.; John, G.; Mohamed, M.F. Characterization of some bacteria isolated from Oreochromis niloticus and their potential use as probiotics. Aquaculture 2008, 277, 1–6. [Google Scholar] [CrossRef]
- Barrows, F.; Gaylord, T.; Sealey, W.; Rawles, S. Database of Nutrient Digestibility’s of Traditional and Novel Feed Ingredients for Trout and Hybrid Striped Bass. USDA-ARS; 2021. Available online: http://www.ars.usda.gov/Main/docs.htm (accessed on 16 October 2020).
- Yu, Z.; Morrison, M. Improved extraction of PCR-quality community DNA from digesta and fecal samples. Biotechniques 2004, 36, 808–812. [Google Scholar] [CrossRef]
- Edwards, U.; Rogall, T.; Blöcker, H.; Emde, M.; Böttger, E.C. Isolation and direct complete nucleotide determination of entire genes. Characterization of a gene coding for 16S ribosomal RNA. Nucleic Acids Res. 1989, 17, 7843–7853. [Google Scholar] [CrossRef] [PubMed]
- Lane, D.J.; Pace, B.; Olsen, G.J.; Stahl, D.A.; Sogin, M.L.; Pace, N.R. Rapid determination of 16S ribosomal RNA sequences for phylogenetic analyses. Proc. Natl. Acad. Sci. USA 1985, 82, 6955–6959. [Google Scholar] [CrossRef] [PubMed]
- Opdahl, L.J.; Gonda, M.G.; St-Pierre, B. Identification of uncultured bacterial species from Firmicutes, Bacteroidetes and CANDIDATUS Saccharibacteria as candidate cellulose utilizers from the rumen of beef cows. Microorganisms 2018, 6, 17. [Google Scholar] [CrossRef] [PubMed]
- Poudel, P.; Levesque, C.L.; Samuel, R.; St-Pierre, B. Dietary inclusion of Peptiva, a peptide-based feed additive, can accelerate the maturation of the fecal bacterial microbiome in weaned pigs. BMC Vet. Res. 2020, 16, 60. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef]
- Haas, B.J.; Gevers, D.; Earl, A.M.; Feldgarden, M.; Ward, D.V.; Giannoukos, G.; Ciulla, D.; Tabbaa, D.; Highlander, S.K.; Sodergren, E. Chimeric 16S rRNA sequence formation and detection in Sanger and 454-pyrosequenced PCR amplicons. Genome Res. 2011, 21, 494–504. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Walron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biology 2011, 12, 1–8. [Google Scholar] [CrossRef]
- Naylor, R.L.; Goldburg, R.J.; Primavera, J.H.; Kautsky, N.; Beveridge, M.C.M.; Clay, J.; Folke, C.; Lubchenco, J.; Mooney, H.; Troell, M. Effects of aquaculture on world fish supplies. Nature 2000, 405, 1017–1024. [Google Scholar] [CrossRef]
- Francis, G.; Makkar, H.P.S.; Becker, K. Antinutritional factors present in plant-derived alternate fish feed ingredients and their effects in fish. Aquaculture 2001, 199, 197–227. [Google Scholar] [CrossRef]
- Dersjant-Li, Y. The use of soy protein in aquafeeds. In Proceedings of the Avances en nutrición acuícola VI. Memorias del VI Simposium Internacional de Nutrición Acuícola., Cancún, Quintana Roo, México, 3–6 September 2002. [Google Scholar]
- Gatlin, D.M.; Barrows, F.T.; Brown, P.; Dabrowski, K.; Gaylord, T.G.; Hardy, R.W.; Herman, E.; Hu, G.; Krogdahl, A.; Nelson, R.; et al. Expanding the utilization of sustainable plant products in aquafeeds: A review. Aquac. Res. 2007, 38, 551–579. [Google Scholar] [CrossRef]
- Suman, G.; Nupur, M.; Anuradha, S.; Pradeep, B. Single-cell protein production: A review. Int. J. Curr. Microbiol. Appl. Sci. 2015, 4, 11. [Google Scholar]
- Whitehead, T.R.; Cotta, M.A.; Falsen, E.; Moore, E.; Lawson, P.A. Peptostreptococcus russellii sp. nov., isolated from a swine-manure storage pit. Int. J. Syst. Evol. Microbiol. 2011, 61, 1875–1879. [Google Scholar] [CrossRef]
- Attwood, G.T.; Klieve, A.V.; Ouwerkerk, D.; Patel, B.K. Ammonia-hyperproducing bacteria from New Zealand ruminants. Appl. Environ. Microbiol. 1998, 64, 1796–1804. [Google Scholar] [CrossRef]
- Whitehead, T.R.; Cotta, M.A. Isolation and identification of hyper-ammonia producing bacteria from swine manure storage pits. Curr. Microbiol. 2004, 48, 20–26. [Google Scholar] [CrossRef]
- Chapagain, P.; Arivett, B.; Cleveland, B.M.; Walker, D.M.; Salem, M. Analysis of the fecal microbiota of fast- and slow-growing rainbow trout (Oncorhynchus mykiss). BMC Genom. 2019, 20, 788. [Google Scholar] [CrossRef]
- Nyman, A.; Huyben, D.; Lundh, T.; Dicksved, J. Effects of microbe-and mussel-based diets on the gut microbiota in Arctic charr (Salvelinus alpinus). Aquac. Rep. 2017, 5, 34–40. [Google Scholar] [CrossRef]
- Wlodarska, M.; Luo, C.; Kolde, R.; d’Hennezel, E.; Annand, J.W.; Heim, C.E.; Krastel, P.; Schmitt, E.K.; Omar, A.S.; Creasey, E.A.; et al. Indoleacrylic acid produced by commensal Peptostreptococcus species suppresses inflammation. Cell Host Microbe 2017, 22, 25–37.e6. [Google Scholar] [CrossRef]
- Cavanagh, D.; Fitzgerald, G.F.; McAuliffe, O. From field to fermentation: The origins of Lactococcus lactis and its domestication to the dairy environment. Food Microbiol. 2015, 47, 45–61. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Lu, Y.; Teng, K.L.; Chen, M.L.; Zheng, H.J.; Zhu, Y.Q.; Zhong, J. Complete genome sequence of Lactococcus lactis subsp. lactis CV56, a probiotic strain isolated from the vaginas of healthy women. J. Bacteriol. 2011, 193, 2886–2887. [Google Scholar] [CrossRef] [PubMed]
- Kato, H.; Shiwa, Y.; Oshima, K.; Machii, M.; Araya-Kojima, T.; Zendo, T.; Shimizu-Kadota, M.; Hattori, M.; Sonomoto, K.; Yoshikawa, H. Complete genome sequence of Lactococcus lactis IO-1, a lactic acid bacterium that utilizes xylose and produces high levels of L-lactic acid. J. Bacteriol. 2012, 194, 2102–2103. [Google Scholar] [CrossRef] [PubMed]
- Kimoto, H.; Kurisaki, J.; Tsuji, N.; Ohmomo, S.; Okamoto, T. Lactococci as probiotic strains: Adhesion to human enterocyte-like Caco-2 cells and tolerance to low pH and bile. Lett. Appl. Microbiol. 1999, 29, 313–316. [Google Scholar] [CrossRef]
- Meyrand, M.; Guillot, A.; Goin, M.; Furlan, S.; Armalyte, J.; Kulakauskas, S.; Cortes-Perez, N.G.; Thomas, G.; Chat, S.; Pechoux, C. Surface proteome analysis of a natural isolate of Lactococcus lactis reveals the presence of pili able to bind human intestinal epithelial cells. Mol. Cell Proteom. 2013, 12, 3935–3947. [Google Scholar] [CrossRef]
- Mercier-Bonin, M.; Chapot-Chartier, M.-P. Surface Proteins of Lactococcus lactis: Bacterial resources for muco-adhesion in the gastrointestinal tract. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Koshio, S.; Ishikawa, M.; Yokoyama, S.; El Basuini, M.F.; Hossain, M.S.; Nhu, T.H.; Dossou, S.; Moss, A.S. Effects of dietary supplementation of Lactobacillus rhamnosus or/and Lactococcus lactis on the growth, gut microbiota and immune responses of red sea bream, Pagrus major. Fish Shellfish Immunol. 2016, 49, 275–285. [Google Scholar] [CrossRef]
- Nguyen, T.L.; Chun, W.-K.; Kim, A.; Kim, N.; Roh, H.J.; Lee, Y.; Yi, M.; Kim, S.; Park, C.-I.; Kim, D.-H. Dietary probiotic effect of Lactococcus lactis WFLU12 on low-molecular-weight metabolites and growth of olive flounder (Paralichythys olivaceus). Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Girijakumari Nisha, R.; Rajathi, V.; Manikandan, R.; Marimuthu Prabhu, N. Isolation of Plesiomonas shigelloides from Infected Cichlid fishes using 16S rRNA characterization and its control with probiotic Pseudomonas sp. Acta Sci. Vet. 2014, 42, 1–7. [Google Scholar]
- Roeselers, G.; Mittge, E.K.; Stephens, W.Z.; Parichy, D.M.; Cavanaugh, C.M.; Guillemin, K.; Rawls, J.F. Evidence for a core gut microbiota in the zebrafish. ISME J. 2011, 5, 1595–1608. [Google Scholar] [CrossRef]
- Liu, Z.; Ke, X.; Lu, M.; Gao, F.; Cao, J.; Zhu, H.; Wang, M. Identification and pathological observation of a pathogenic Plesiomonas shigelloides strain isolated from cultured tilapia (Oreochromis niloticus). Acta Microbiol. Sin. 2015, 55, 96–106. [Google Scholar]
- Bai, Z.; Ren, T.; Han, Y.; Rahman, M.M.; Hu, Y.; Li, Z.; Jiang, Z. Influences of dietary selenomethionine exposure on tissue accumulation, blood biochemical profiles, gene expression and intestinal microbiota of Carassius auratus. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2019, 218, 21–29. [Google Scholar] [CrossRef]
- Ryan, M.P.; Pembroke, J.T.; Adley, C.C. Ralstonia pickettii in environmental biotechnology: Potential and applications. J. Appl. Microbiol. 2007, 103, 754–764. [Google Scholar] [CrossRef]
- Makaritsis, K.P.; Neocleous, C.; Gatselis, N.; Petinaki, E.; Dalekos, G.N. An immunocompetent patient presenting with severe septic arthritis due to Ralstonia pickettii identified by molecular-based assays: A case report. Cases J. 2009, 2, 8125. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Ingredient | Diet | ||||
|---|---|---|---|---|---|
| BP-F1 | BP-E | BP-W | BP | CON | |
| BP-SBM Fraction #1 a | 25.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| BP-SBM Fraction #2 a | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| BP-SBM Fraction #3 a | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| BP-SBM + Enzyme a | 0.00 | 25.00 | 0.00 | 0.00 | 0.00 |
| BP-SBM Base + Wash a | 0.00 | 0.00 | 25.00 | 0.00 | 0.00 |
| BP-SBM Base a | 0.00 | 0.00 | 0.00 | 25.00 | 0.00 |
| Blood Meal b | 2.50 | 2.50 | 2.50 | 2.50 | 2.50 |
| Wheat Midds c | 10.00 | 10.00 | 10.00 | 10.00 | 21.92 |
| Whole Cleaned Wheat d | 16.67 | 16.67 | 16.67 | 16.67 | 15.00 |
| Poultry Meal e | 12.00 | 12.00 | 12.00 | 12.00 | 23.00 |
| Feather Meal e | 2.50 | 2.50 | 2.50 | 2.50 | 7.50 |
| Fish Meal f | 10.00 | 10.00 | 10.00 | 10.00 | 10.00 |
| Vitamin Premix g | 1.25 | 1.25 | 1.25 | 1.25 | 1.25 |
| Lysine h | 1.00 | 1.00 | 1.00 | 1.00 | 1.75 |
| Methionine h | 0.50 | 0.50 | 0.50 | 0.50 | 0.50 |
| Choline Chloride i | 0.58 | 0.58 | 0.58 | 0.58 | 0.58 |
| Mineral Premix j | 0.75 | 0.75 | 0.75 | 0.75 | 0.75 |
| Stay C k | 0.25 | 0.25 | 0.25 | 0.25 | 0.25 |
| Fish Oil l | 6.50 | 6.50 | 6.50 | 6.50 | 4.50 |
| Dicalcium phosphate m | 0.50 | 0.50 | 0.50 | 0.50 | 0.50 |
| Defatted SBM n | 10.00 | 10.00 | 10.00 | 10.00 | 10.00 |
| Totals | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 |
| Diet | Ash | Fat | Fiber | Protein | NFE |
|---|---|---|---|---|---|
| BP-F1 | 7.71 | 8.79 | 4.65 | 47.32 | 31.53 |
| BP-F2 | 7.89 | 8.53 | 3.86 | 47.92 | 31.80 |
| BP-F3 | 7.63 | 8.97 | 4.47 | 47.25 | 31.68 |
| BP-E | 7.71 | 8.99 | 5.33 | 47.32 | 30.64 |
| BP-W | 7.77 | 8.49 | 5.62 | 47.02 | 31.10 |
| BP | 8.07 | 8.13 | 5.27 | 47.65 | 30.88 |
| CON | 8.88 | 8.42 | 4.52 | 45.49 | 32.69 |
| Diet | Gain 1 | Consumption 2 | FCR 3 | VSI 4 | HIS 5 |
|---|---|---|---|---|---|
| BP-F1 | 128.2 a | 185.0 ab | 1.44 c | 8.85 a | 1.49 bc |
| BP-F2 | 119.9 a | 178.5 b | 1.49 b | 8.61 a | 1.44 bc |
| BP-F3 | 110.8 a | 179.6 b | 1.62 b | 9.09 a | 1.40 c |
| BP-E | 105.6 a | 163.5 b | 1.55 b | 9.28 a | 1.56 abc |
| BP-W | 126.6 a | 183.8 ab | 1.45 bc | 8.57 a | 1.44 bc |
| BP | 119.7 a | 184.0 ab | 1.54 bc | 9.32 a | 1.64 ab |
| CON | 109.8 a | 211.2 a | 1.93 a | 8.60 a | 1.76 a |
| Taxonomic Affiliation | CON | BP-F1 | BP-E | BP-W | BP | p Value |
|---|---|---|---|---|---|---|
| Firmicutes | 95.32 | 12.83 | 61.53 | 61.30 | 88.92 | 0.08 |
| Peptostreptococcaceae# | 72.66 c | 6.90 abc | 1.67 a | 6.22 ab | 66.54 bc | 0.02 |
| Streptococcaceae# | 4.09 ab | 2.29 a | 52.09 b | 49.18 b | 6.40 ab | 0.02 |
| Leuconostocaceae# | 0.10 b | 1.05 a | 4.64a | 2.10 a | 0.17 ab | 0.03 |
| Peptoniphilaceae# | 1.90 b | 0.22 ab | 0.07a | 0.23 ab | 2.35 b | 0.02 |
| Clostridiaceae 1 # | 3.27 d | 0.44 abc | 0.06 b | 0.25 abc | 1.38 cd | 0.01 |
| unclass. Clostridiales x | 8.89 | 0.78 | 0.29 | 0.84 | 7.94 | - |
| Other Firmicutes x | 4.42 | 1.16 | 2.70 | 2.47 | 4.15 | - |
| Proteobacteria | 2.00 | 85.23 | 37.04 | 37.03 | 7.43 | 0.06 |
| Enterobacteriaceae# | 1.12 c | 31.99 a | 31.28 ac | 1.02 bc | 6.10 a | 0.03 |
| Sphingomonadaceae | 0.30 | 18.04 | 2.46 | 15.29 | 0.25 | 0.26 |
| Burkholderiaceae | 0.24 | 34.43 | 3.01 | 20.06 | 0.26 | 0.11 |
| Other Proteobacteria x | 0.35 | 0.77 | 0.29 | 0.65 | 0.83 | - |
| Bacteriodetes | 2.25 | 0.79 | 0.49 | 0.74 | 3.31 | 0.06 |
| Porphyromonadaceae | 1.25 | 0.28 | 0.18 | 0.29 | 1.63 | 0.06 |
| Other Bacteroidetes x | 1.00 | 0.52 | 0.31 | 0.45 | 1.67 | - |
| Other Bacteria x$ | 0.42 | 1.14 | 0.94 | 0.93 | 0.34 | - |
| Index | CON | BP-F1 | BP-E | BP-W | BP | p-Value |
|---|---|---|---|---|---|---|
| Observed OTUs # | 266.67 b | 172.67 a | 134.00 a | 169.67 a | 301.67 b | <0.001 |
| Ace # | 799.75 b | 397.48 a | 305.62 a | 377.39 a | 725.13 b | <0.001 |
| Chao # | 550.36 b | 284.72 a | 269.85 a | 306.63 a | 545.64 b | <0.001 |
| Shannon | 2.81 | 1.94 | 1.59 | 2.17 | 3.03 | 0.078 |
| Simpson | 0.24 | 0.40 | 0.53 | 0.32 | 0.20 | 0.287 |
| OTUs | CON | BP-F1 | BP-E | BP-W | BP | p-Value | Closest Taxon (id%) |
|---|---|---|---|---|---|---|---|
| Proteobacteria | |||||||
| SD_McMs-00002 # | 1.03 ab | 30.28 a | 29.96 ab | 0.84 b | 4.82 a | 0.05 | Pl. shigelloides (99%) |
| SD_McMs-00003 | 0.18 | 27.22 | 2.44 | 16.41 | 0.21 | 0.09 | R. pickettii (99%) |
| SD_McMs-00004 | 0.23 | 13.54 | 1.90 | 11.97 | 0.18 | 0.29 | Sp. elodea (99%) |
| SD_McMs-00005 | 0.02 | 1.92 | 0.14 | 0.92 | 0.01 | 0.16 | R. pickettii (98%) |
| SD_McMs-00006 | 0.03 | 3.39 | 0.26 | 2.24 | 0.03 | 0.24 | R. pickettii (99%) |
| Firmicutes | |||||||
| SD_McMs-00001 # | 47.61 a | 4.29 abc | 1.15 c | 3.91 bc | 43.13 ab | 0.02 | Ps. russellii (91%) |
| SD_McMs-00007 # | 0.86 a | 1.35 ab | 43.47 b | 41.01 b | 1.59 ab | 0.02 | La. lactis (100%) |
| SD_McMs-00008 | 0.16 | 0.16 | 1.72 | 3.19 | 0.34 | 0.08 | La. lactis (96%) |
| SD_McMs-00009 # | 0.03 a | 0.10 ab | 2.25 b | 1.20 b | 0.10 ab | 0.02 | Le. citreum (100%) |
| SD_McMs-00010 # | 0.98 c | 0.05 a | 0.10 ac | 0.29 abc | 1.97 bc | 0.02 | St. dysgalactiae (100%) |
| SD_McMs-00011 # | 7.41 a | 0.86 ab | 0.11 b | 0.75 ab | 7.20 a | 0.02 | Ps. russellii (99%) |
| SD_McMs-00012 # | 3.99 a | 0.31 ab | 0.07 b | 0.27 ab | 3.98 a | 0.02 | Ps. russellii (91%) |
| SD_McMs-00013 # | 3.97 a | 0.28 b | 0.11 b | 0.37 ab | 3.30 ab | 0.02 | Pe. stercorisuis (89%) |
| SD_McMs-00014 # | 1.98 bc | 0.16 ac | 0.04 a | 0.17 ab | 2.32 b | 0.02 | Ps. russellii (94%) |
| SD_McMs-00015 # | 1.81 a | 0.17 ab | 0.04 b | 0.10 ab | 1.51 a | 0.02 | Ps. russellii (94%) |
| SD_McMs-00016 # | 0.02 c | 0.31 ab | 1.96 b | 0.72 ab | 0.04 ac | 0.03 | La. lactis (96%) |
| Bacteroidetes | |||||||
| SD_McMs-00017 | 1.18 | 0.25 | 0.16 | 0.27 | 1.42 | 0.06 | F. endometrii (99%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fowler, E.C.; Poudel, P.; White, B.; St-Pierre, B.; Brown, M. Effects of a Bioprocessed Soybean Meal Ingredient on the Intestinal Microbiota of Hybrid Striped Bass, Morone chrysops x M. saxatilis. Microorganisms 2021, 9, 1032. https://doi.org/10.3390/microorganisms9051032
Fowler EC, Poudel P, White B, St-Pierre B, Brown M. Effects of a Bioprocessed Soybean Meal Ingredient on the Intestinal Microbiota of Hybrid Striped Bass, Morone chrysops x M. saxatilis. Microorganisms. 2021; 9(5):1032. https://doi.org/10.3390/microorganisms9051032
Chicago/Turabian StyleFowler, Emily Celeste, Prakash Poudel, Brandon White, Benoit St-Pierre, and Michael Brown. 2021. "Effects of a Bioprocessed Soybean Meal Ingredient on the Intestinal Microbiota of Hybrid Striped Bass, Morone chrysops x M. saxatilis" Microorganisms 9, no. 5: 1032. https://doi.org/10.3390/microorganisms9051032
APA StyleFowler, E. C., Poudel, P., White, B., St-Pierre, B., & Brown, M. (2021). Effects of a Bioprocessed Soybean Meal Ingredient on the Intestinal Microbiota of Hybrid Striped Bass, Morone chrysops x M. saxatilis. Microorganisms, 9(5), 1032. https://doi.org/10.3390/microorganisms9051032