
A Comparative Pilot Study of Bacterial and Fungal Dysbiosis in Neurodevelopmental Disorders and Gastrointestinal Disorders: Commonalities, Specificities and Correlations with Lifestyle

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Recruitment
2.2. Questionnaire
2.3. Sample Collection
2.4. DNA Extraction
2.5. 16sRNA Sequencing and Phylogenetic Analysis
2.6. ITS Sequencing and Phylogenetic Analysis
2.7. Statistical Analyses
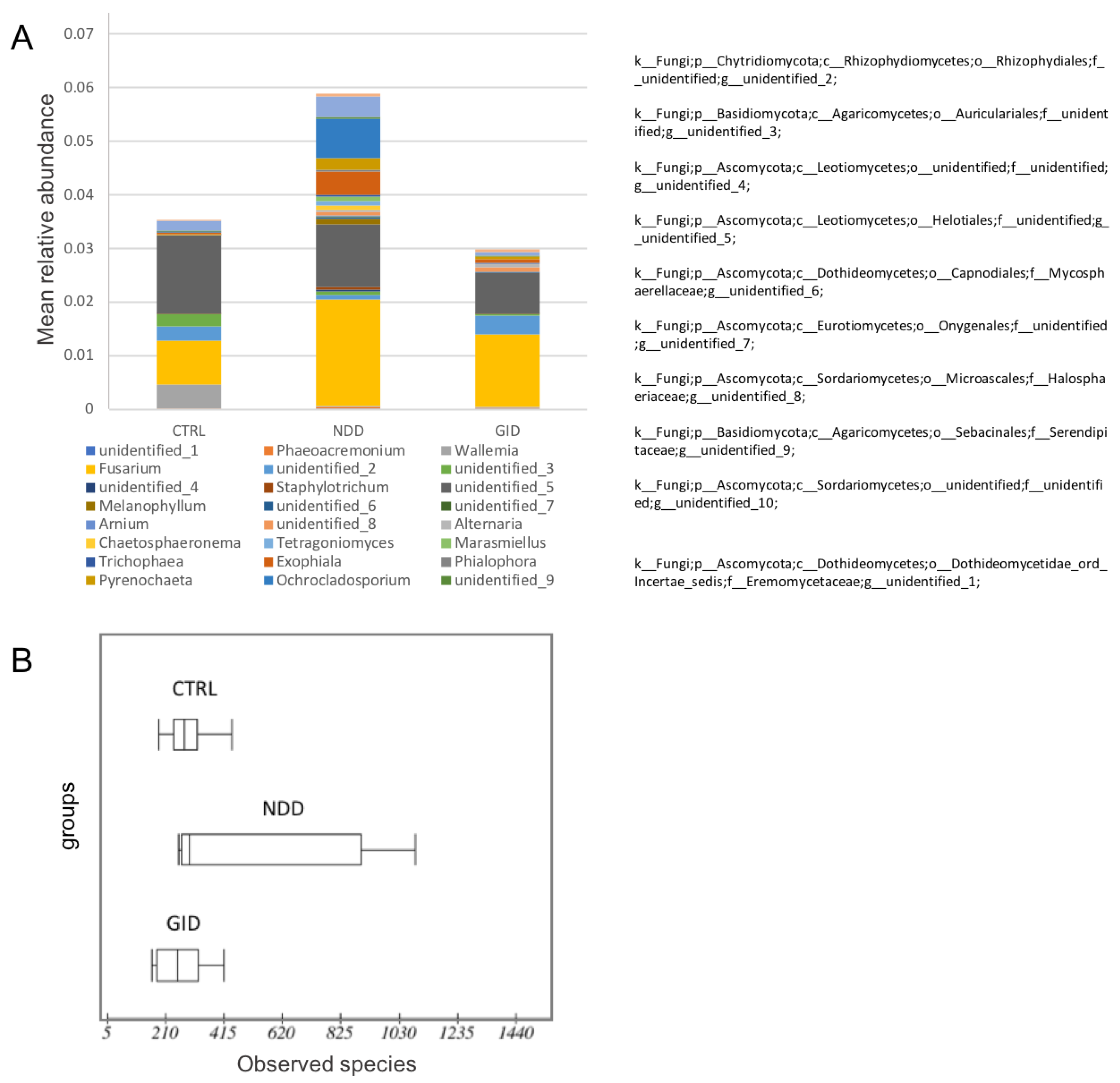
3. Results
3.1. Characterisistics of the Studied Cohort
3.2. Bacterial Microbiome in NDD, GID and CTRL
3.2.1. An Altered Bacterial Diversity in NDD and GID Groups
3.2.2. Significant Taxa Abundance Variability in NDD Compared to CTRL
3.2.3. Comparing NDD Group to GID Group: Commonalities and Specificities in the Bacteriome
3.3. The Fungal Microbiome or Mycobiome: A Neglected but Important Player
3.3.1. An Increased Fungal Diversity in the NDD Group
3.3.2. An Increased Fungal Abundance in the NDD Group
3.3.3. Mycobiome Is Key to Differentiate between NDD and GID Dysbiosis
3.4. Exposure to Antibiotics at Early Age Is Associated with a Significant Fungal Dysbiosis
3.4.1. Effect of Recurrent Exposure to Antibiotics before 3 Years: An Independent t-Test
3.4.2. Regression Analysis for the Effect of ABX
4. Discussion
4.1. From GID Group to NDD Group: What Do They Have in Common, and in What Ways Are They Different?
4.2. Need for Standard Measurements and Limitations of the Study
4.3. Effect of Probiotics and Diet
4.4. Effect of Early Antibiotic Exposure
4.5. Important Taxa to Explore Further
4.6. Importance of the Gut Microbiome in Immunity Development
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Level | NDD Absolute Abundance Compared to CTRL | Name | Basic Covariates | Adding Probiotics | Adding Diet | Adding Probiotics and Diet | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| DF | R-Squared | Beta | SEbeta | p-Value | p-Value 1 | p-Value 2 | p-Value 3 | |||
| Phylum | Chytridiomycota | k__Fungi;p__Chytridiomycota; | 14 | 0.4210 | −1.8554 | 0.5119 | 0.0028 | 0.0019 | 0.0029 | 0.0013 |
| Class | Rhizophydiomycetes | k__Fungi;p__Chytridiomycota;c__Rhizophydiomycetes; | 14 | 0.4610 | −1.7178 | 0.4920 | 0.0036 | 0.0249 | NS | 0.0111 |
| Orbiliomycetes | k__Fungi;p__Ascomycota;c__Orbiliomycetes; | 14 | 0.2538 | 0.7096 | 0.3110 | 0.0387 | NS | NS | NS | |
| Pezizomycetes | k__Fungi;p__Ascomycota;c__Pezizomycetes; | 14 | 0.1742 | −1.3952 | 0.6216 | 0.0415 | 0.0144 | 0.0378 | 0.0073 | |
| Ustilaginomycetes | k__Fungi;p__Basidiomycota;c__Ustilaginomycetes; | 14 | 0.1186 | 1.6869 | 0.7778 | 0.0478 | NS | 0.0538 | NS | |
| Spizellomycetes | k__Fungi;p__Chytridiomycota;c__Spizellomycetes; | 12 | 0.2570 | −1.8647 | 0.7718 | NS | 0.0325 | NS | 0.0496 | |
| Agaricomycetes | k__Fungi;p__Basidiomycota;c__Agaricomycetes; | 12 | 0.7201 | −1.1365 | 0.4985 | NS | 0.0417 | NS | NS | |
| Eurotiomycetes | k__Fungi;p__Ascomycota;c__Eurotiomycetes; | 13 | 0.1476 | 0.7961 | 0.3702 | NS | NS | 0.0509 | NS | |
| unidentified | k__Fungi;p__Ascomycota;c__unidentified; | 11 | 0.2480 | −1.7915 | 0.7848 | NS | NS | NS | 0.0433 | |
| Family | Alphamycetaceae | k__Fungi;p__Chytridiomycota;c__Rhizophydiomycetes;o__Rhizophydiales;f__Alphamycetaceae; | 14 | 0.3128 | −1.1437 | 0.3700 | 0.0080 | 0.0042 | 0.0510 | |
| unidentified | k__Fungi;p__Ascomycota;c__Leotiomycetes;o__unidentified;f__unidentified; | 14 | 0.4182 | 1.3737 | 0.4703 | 0.0112 | 0.0264 | 0.0180 | 0.0499 | |
| unidentified | k__Fungi;p__Chytridiomycota;c__Rhizophydiomycetes;o__Rhizophydiales;f__unidentified; | 14 | 0.3771 | −1.3958 | 0.5048 | 0.0152 | 0.0215 | 0.0164 | 0.0183 | |
| Nectriaceae | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Hypocreales;f__Nectriaceae; | 14 | 0.3147 | 1.6213 | 0.6036 | 0.0177 | NS | 0.0253 | NS | |
| Valsaceae | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Diaporthales;f__Valsaceae; | 14 | 0.3067 | 1.1744 | 0.4458 | 0.0196 | NS | 0.0239 | NS | |
| Sordariales_fam_Incertae_sedis | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Sordariales;f__Sordariales_fam_Incertae_sedis; | 14 | 0.3809 | 0.2410 | 0.0986 | 0.0284 | NS | NS | NS | |
| Entolomataceae | k__Fungi;p__Basidiomycota;c__Agaricomycetes;o__Agaricales;f__Entolomataceae; | 14 | 0.2524 | −1.8388 | 0.7595 | 0.0296 | 0.0316 | 0.0306 | 0.0255 | |
| Glomerellaceae | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Glomerellales;f__Glomerellaceae; | 14 | 0.2860 | 1.5580 | 0.6711 | 0.0358 | NS | 0.0406 | NS | |
| Ustilaginaceae | k__Fungi;p__Basidiomycota;c__Ustilaginomycetes;o__Ustilaginales;f__Ustilaginaceae; | 14 | 0.1186 | 1.6869 | 0.7778 | 0.0478 | NS | 0.0538 | NS | |
| unidentified | k__Fungi;p__Basidiomycota;c__Agaricomycetes;o__Auriculariales;f__unidentified; | 12 | 0.3450 | −2.1705 | 0.6456 | NS | 0.0057 | NS | 0.0124 | |
| unidentified | k__Fungi;p__Ascomycota;c__Leotiomycetes;o__Helotiales;f__unidentified; | 12 | 0.4483 | −0.7760 | 0.2780 | NS | 0.0163 | NS | 0.0160 | |
| Herpotrichiellaceae | k__Fungi;p__Ascomycota;c__Eurotiomycetes;o__Chaetothyriales;f__Herpotrichiellaceae; | 12 | 0.2111 | 2.3400 | 0.9128 | NS | 0.0248 | NS | 0.0304 | |
| Halosphaeriaceae | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Microascales;f__Halosphaeriaceae; | 12 | 0.2201 | 1.7823 | 0.7234 | NS | 0.0298 | NS | 0.0246 | |
| unidentified | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Branch06;f__unidentified; | 12 | 0.3074 | −1.8874 | 0.7661 | NS | 0.0298 | NS | 0.0428 | |
| unidentified | k__Fungi;p__Ascomycota;c__Eurotiomycetes;o__Onygenales;f__unidentified; | 12 | 0.2042 | 1.1675 | 0.4939 | NS | 0.0358 | NS | 0.0266 | |
| Tetragoniomycetaceae | k__Fungi;p__Basidiomycota;c__Tremellomycetes;o__Trichosporonales;f__Tetragoniomycetaceae; | 12 | 0.1551 | 2.3156 | 0.9845 | NS | 0.0366 | NS | 0.0413 | |
| Omphalotaceae | k__Fungi;p__Basidiomycota;c__Agaricomycetes;o__Agaricales;f__Omphalotaceae; | 12 | 0.1551 | 2.3156 | 0.9845 | NS | 0.0366 | NS | 0.0413 | |
| Dipodascaceae | k__Fungi;p__Ascomycota;c__Saccharomycetes;o__Saccharomycetales;f__Dipodascaceae; | 12 | 0.0943 | −0.1554 | 0.0662 | NS | 0.0368 | NS | 0.0438 | |
| Cucurbitariaceae | k__Fungi;p__Ascomycota;c__Dothideomycetes;o__Pleosporales;f__Cucurbitariaceae; | 12 | 0.1366 | 1.5240 | 0.6699 | NS | 0.0421 | NS | 0.0469 | |
| Sordariales_fam_Incertae_sedis | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Sordariales;f__Sordariales_fam_Incertae_sedis; | 13 | 0.3350 | 0.2445 | 0.1040 | NS | NS | 0.0351 | NS | |
| Plectosphaerellaceae | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Glomerellales;f__Plectosphaerellaceae; | 11 | 0.3344 | −1.5347 | 0.6468 | NS | NS | NS | 0.0370 | |
| unidentified | k__Fungi;p__Ascomycota;c__unidentified;o__unidentified;f__unidentified; | 11 | 0.2480 | −1.7915 | 0.7848 | NS | NS | NS | 0.0433 | |
| Order | Rhizophydiales | k__Fungi;p__Chytridiomycota;c__Rhizophydiomycetes;o__Rhizophydiales; | 14 | 0.4610 | −1.7178 | 0.4920 | 0.0036 | 0.0249 | 0.0017 | 0.0111 |
| unidentified | k__Fungi;p__Ascomycota;c__Leotiomycetes;o__unidentified; | 14 | 0.4182 | 1.3737 | 0.4703 | 0.0112 | 0.0264 | 0.0180 | 0.0499 | |
| Diaporthales | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Diaporthales; | 14 | 0.3451 | 1.4422 | 0.4998 | 0.0120 | 0.0534 | 0.0146 | NS | |
| Orbiliales | k__Fungi;p__Ascomycota;c__Orbiliomycetes;o__Orbiliales; | 14 | 0.3375 | 0.4525 | 0.1919 | 0.0334 | NS | 0.0520 | NS | |
| Pezizales | k__Fungi;p__Ascomycota;c__Pezizomycetes;o__Pezizales; | 14 | 0.1964 | −1.3952 | 0.6162 | 0.0400 | 0.0149 | 0.0352 | 0.0070 | |
| Ustilaginales | k__Fungi;p__Basidiomycota;c__Ustilaginomycetes;o__Ustilaginales; | 14 | 0.1186 | 1.6869 | 0.7778 | 0.0478 | NS | 0.0538 | NS | |
| Auriculariales | k__Fungi;p__Basidiomycota;c__Agaricomycetes;o__Auriculariales; | 12 | 0.3452 | −2.1724 | 0.6460 | NS | 0.0056 | NS | 0.0124 | |
| Branch06 | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Branch06; | 12 | 0.3074 | −1.8874 | 0.7661 | NS | 0.0298 | NS | 0.0428 | |
| Spizellomycetales | k__Fungi;p__Chytridiomycota;c__Spizellomycetes;o__Spizellomycetales; | 12 | 0.2570 | −1.8647 | 0.7718 | NS | 0.0325 | NS | 0.0496 | |
| Chaetothyriales | k__Fungi;p__Ascomycota;c__Eurotiomycetes;o__Chaetothyriales; | 12 | 0.1421 | 2.1636 | 0.9841 | NS | 0.0483 | NS | NS | |
| Leotiomycetes_ord_Incertae_sedis | k__Fungi;p__Ascomycota;c__Leotiomycetes;o__Leotiomycetes_ord_Incertae_sedis; | 11 | 0.1224 | −2.1509 | 0.8922 | NS | NS | NS | 0.0346 | |
| unidentified | k__Fungi;p__Ascomycota;c__unidentified;o__unidentified; | 11 | 0.2480 | −1.7915 | 0.7848 | NS | NS | NS | 0.0433 | |
| Genus | Fusarium | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Hypocreales;f__Nectriaceae;g__Fusarium; | 14 | 0.4268 | 1.8384 | 0.5123 | 0.0030 | 0.0358 | 0.0051 | NS |
| Saccharomyces | k__Fungi;p__Ascomycota;c__Saccharomycetes;o__Saccharomycetales;f__Saccharomycetaceae;g__Saccharomyces; | 14 | 0.4150 | 1.1286 | 0.3333 | 0.0044 | 0.0474 | 0.0069 | NS | |
| Betamyces | k__Fungi;p__Chytridiomycota;c__Rhizophydiomycetes;o__Rhizophydiales;f__Alphamycetaceae;g__Betamyces; | 14 | 0.3128 | −1.1437 | 0.3700 | 0.0080 | 0.0042 | 0.0510 | ||
| unidentified | k__Fungi;p__Ascomycota;c__Leotiomycetes;o__unidentified;f__unidentified;g__unidentified; | 14 | 0.4182 | 1.3737 | 0.4703 | 0.0112 | 0.0264 | 0.0180 | 0.0499 | |
| unidentified | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Diaporthales;f__Valsaceae;g__unidentified; | 14 | 0.3207 | 1.9895 | 0.7075 | 0.0139 | 0.0166 | NS | ||
| unidentified | k__Fungi;p__Chytridiomycota;c__Rhizophydiomycetes;o__Rhizophydiales;f__unidentified;g__unidentified; | 14 | 0.3771 | −1.3958 | 0.5048 | 0.0152 | 0.0215 | 0.0164 | 0.0183 | |
| Staphylotrichum | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Sordariales;f__Sordariales_fam_Incertae_sedis;g__Staphylotrichum; | 14 | 0.5841 | 0.7721 | 0.2800 | 0.0154 | 0.0454 | 0.0210 | NS | |
| Engyodontium | k__Fungi;p__Ascomycota;c__Ascomycota_cls_Incertae_sedis;o__Ascomycota_ord_Incertae_sedis;f__Ascomycota_fam_Incertae_sedis;g__Engyodontium; | 14 | 0.3469 | −0.7645 | 0.2794 | 0.0161 | NS | 0.0116 | NS | |
| Colletotrichum | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Glomerellales;f__Glomerellaceae;g__Colletotrichum; | 14 | 0.2860 | 1.5580 | 0.6711 | 0.0358 | NS | 0.0406 | NS | |
| Neonectria | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Hypocreales;f__Nectriaceae;g__Neonectria; | 14 | 0.1046 | 1.6340 | 0.7143 | 0.0382 | NS | 0.0533 | NS | |
| Melanophyllum | k__Fungi;p__Basidiomycota;c__Agaricomycetes;o__Agaricales;f__Agaricaceae;g__Melanophyllum; | 14 | 0.2073 | 1.7648 | 0.7733 | 0.0386 | 0.0156 | 0.0468 | 0.0190 | |
| unidentified | k__Fungi;p__Mucoromycota;c__Mucoromycetes;o__Mucorales;f__Lichtheimiaceae;g__unidentified; | 14 | 0.0957 | 0.0873 | 0.0391 | 0.0423 | NS | 0.0463 | NS | |
| unidentified | k__Fungi;p__Basidiomycota;c__Agaricomycetes;o__Auriculariales;f__unidentified;g__unidentified; | 11 | 0.3016 | −2.0728 | 0.6945 | NS | 0.0057 | NS | 0.0124 | |
| unidentified | k__Fungi;p__Ascomycota;c__Leotiomycetes;o__Helotiales;f__unidentified;g__unidentified; | 11 | 0.4276 | −0.8382 | 0.2949 | NS | 0.0163 | NS | 0.0160 | |
| Mortierella | k__Fungi;p__Mortierellomycota;c__Mortierellomycetes;o__Mortierellales;f__Mortierellaceae;g__Mortierella; | 11 | 0.4528 | −1.7326 | 0.6124 | NS | 0.0222 | NS | 0.0164 | |
| unidentified | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Microascales;f__Halosphaeriaceae;g__unidentified; | 11 | 0.2080 | 1.9764 | 0.7597 | NS | 0.0298 | NS | 0.0246 | |
| unidentified | k__Fungi;p__Ascomycota;c__Dothideomycetes;o__Capnodiales;f__Mycosphaerellaceae;g__unidentified; | 11 | 0.1891 | 1.1674 | 0.4489 | NS | 0.0210 | NS | 0.0247 | |
| unidentified | k__Fungi;p__Ascomycota;c__Eurotiomycetes;o__Onygenales;f__unidentified;g__unidentified; | 11 | 0.2070 | 1.3142 | 0.5135 | NS | 0.0358 | NS | 0.0266 | |
| Arnium | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Sordariales;f__Lasiosphaeriaceae;g__Arnium; | 11 | 0.0973 | 2.1493 | 0.9062 | NS | 0.0277 | NS | 0.0370 | |
| Chaetosphaeronema | k__Fungi;p__Ascomycota;c__Dothideomycetes;o__Pleosporales;f__Phaeosphaeriaceae;g__Chaetosphaeronema; | 11 | 0.0922 | 2.4429 | 1.0354 | NS | 0.0273 | NS | 0.0379 | |
| Tetragoniomyces | k__Fungi;p__Basidiomycota;c__Tremellomycetes;o__Trichosporonales;f__Tetragoniomycetaceae;g__Tetragoniomyces; | 11 | 0.0954 | 2.4512 | 1.0611 | NS | NS | NS | 0.0413 | |
| Marasmiellus | k__Fungi;p__Basidiomycota;c__Agaricomycetes;o__Agaricales;f__Omphalotaceae;g__Marasmiellus; | 11 | 0.0954 | 2.4512 | 1.0611 | NS | 0.0366 | NS | 0.0413 | |
| Trichophaea | k__Fungi;p__Ascomycota;c__Pezizomycetes;o__Pezizales;f__Pyronemataceae;g__Trichophaea; | 11 | 0.0954 | 2.4512 | 1.0611 | NS | 0.0366 | NS | 0.0413 | |
| unidentified | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Branch06;f__unidentified;g__unidentified; | 11 | 0.2450 | −1.9076 | 0.8332 | NS | 0.0298 | NS | 0.0428 | |
| unidentified | k__Fungi;p__Ascomycota;c__unidentified;o__unidentified;f__unidentified;g__unidentified; | 11 | 0.2480 | −1.7915 | 0.7848 | NS | NS | NS | 0.0433 | |
| Exophiala | k__Fungi;p__Ascomycota;c__Eurotiomycetes;o__Chaetothyriales;f__Herpotrichiellaceae;g__Exophiala; | 11 | 0.0584 | 2.3498 | 1.0471 | NS | 0.0383 | NS | 0.0464 | |
| Pyrenochaeta | k__Fungi;p__Ascomycota;c__Dothideomycetes;o__Pleosporales;f__Cucurbitariaceae;g__Pyrenochaeta; | 11 | 0.0663 | 1.6738 | 0.7547 | NS | 0.0435 | NS | 0.0486 | |
| Plectosphaerella | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Glomerellales;f__Plectosphaerellaceae;g__Plectosphaerella; | 11 | 0.1903 | −1.3601 | 0.6277 | NS | NS | NS | 0.0531 | |
| Species | Saccharomyces_cerevisiae | k__Fungi;p__Ascomycota;c__Saccharomycetes;o__Saccharomycetales;f__Saccharomycetaceae;g__Saccharomyces;s__Saccharomyces_cerevisiae; | 14 | 0.4150 | 1.1286 | 0.3333 | 0.0044 | 0.0474 | 0.0069 | NS |
| Betamyces_americae-meridionalis | k__Fungi;p__Chytridiomycota;c__Rhizophydiomycetes;o__Rhizophydiales;f__Alphamycetaceae;g__Betamyces;s__Betamyces_americae-meridionalis; | 14 | 0.3128 | −1.1437 | 0.3700 | 0.0080 | NS | 0.0042 | 0.0510 | |
| unidentified | k__Fungi;p__Ascomycota;c__Leotiomycetes;o__unidentified;f__unidentified;g__unidentified;s__unidentified; | 14 | 0.4182 | 1.3737 | 0.4703 | 0.0112 | 0.0264 | 0.0180 | 0.0499 | |
| Peziza_nivalis | k__Fungi;p__Ascomycota;c__Pezizomycetes;o__Pezizales;f__Pezizaceae;g__Peziza;s__Peziza_nivalis; | 14 | 0.2416 | 0.4322 | 0.1520 | 0.0130 | 0.0267 | 0.0145 | 0.0260 | |
| unidentified | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Diaporthales;f__Valsaceae;g__unidentified;s__unidentified; | 14 | 0.3207 | 1.9895 | 0.7075 | 0.0139 | NS | 0.0166 | NS | |
| Fusarium_oxysporum | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Hypocreales;f__Nectriaceae;g__Fusarium;s__Fusarium_oxysporum; | 14 | 0.3128 | 1.4464 | 0.5171 | 0.0143 | NS | 0.0163 | NS | |
| unidentified | k__Fungi;p__Chytridiomycota;c__Rhizophydiomycetes;o__Rhizophydiales;f__unidentified;g__unidentified;s__unidentified; | 14 | 0.3771 | −1.3958 | 0.5048 | 0.0152 | 0.0215 | 0.0164 | 0.0183 | |
| Staphylotrichum_coccosporum | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Sordariales;f__Sordariales_fam_Incertae_sedis;g__Staphylotrichum;s__Staphylotrichum_coccosporum; | 14 | 0.5851 | 0.8130 | 0.2963 | 0.0158 | 0.0449 | 0.0214 | NS | |
| Engyodontium_album | k__Fungi;p__Ascomycota;c__Ascomycota_cls_Incertae_sedis;o__Ascomycota_ord_Incertae_sedis;f__Ascomycota_fam_Incertae_sedis;g__Engyodontium;s__Engyodontium_album; | 14 | 0.3469 | −0.7645 | 0.2794 | 0.0161 | NS | 0.0116 | NS | |
| Chaetomium_erectum | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Sordariales;f__Chaetomiaceae;g__Chaetomium;s__Chaetomium_erectum; | 14 | 0.2427 | 1.8071 | 0.7522 | 0.0307 | 0.0143 | 0.0378 | 0.0176 | |
| Neonectria_candida | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Hypocreales;f__Nectriaceae;g__Neonectria;s__Neonectria_candida; | 14 | 0.1046 | 1.6340 | 0.7143 | 0.0382 | NS | 0.0533 | NS | |
| Melanophyllum_haematospermum | k__Fungi;p__Basidiomycota;c__Agaricomycetes;o__Agaricales;f__Agaricaceae;g__Melanophyllum;s__Melanophyllum_haematospermum; | 14 | 0.2073 | 1.7648 | 0.7733 | 0.0386 | 0.0156 | 0.0468 | 0.0190 | |
| unidentified | k__Fungi;p__Mucoromycota;c__Mucoromycetes;o__Mucorales;f__Lichtheimiaceae;g__unidentified;s__unidentified; | 14 | 0.0957 | 0.0873 | 0.0391 | 0.0423 | NS | 0.0463 | NS | |
| Spizellomyces_dolichospermus | k__Fungi;p__Chytridiomycota;c__Spizellomycetes;o__Spizellomycetales;f__Spizellomycetaceae;g__Spizellomyces;s__Spizellomyces_dolichospermus; | 14 | 0.2033 | −1.7153 | 0.7748 | 0.0439 | 0.0455 | 0.0501 | 0.0442 | |
| Hypochnicium_bombycinum | k__Fungi;p__Basidiomycota;c__Agaricomycetes;o__Corticiales;f__Corticiaceae;g__Hypochnicium;s__Hypochnicium_bombycinum; | 14 | 0.1286 | 0.7133 | 0.3367 | 0.0525 | NS | NS | NS | |
| Microascus_paisii | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Microascales;f__Microascaceae;g__Microascus;s__Microascus_paisii; | 14 | 0.1093 | 0.7894 | 0.3730 | 0.0527 | NS | 0.0442 | NS | |
| unidentified | k__Fungi;p__Ascomycota;c__Leotiomycetes;o__Helotiales;f__unidentified;g__unidentified;s__unidentified; | 12 | 0.4483 | −0.7760 | 0.2780 | NS | 0.0163 | NS | 0.0160 | |
| unidentified | k__Fungi;p__Basidiomycota;c__Agaricomycetes;o__Auriculariales;f__unidentified;g__unidentified;s__unidentified; | 12 | 0.3450 | −2.1705 | 0.6456 | NS | 0.0057 | NS | 0.0124 | |
| unidentified | k__Fungi;p__Ascomycota;c__Dothideomycetes;o__Capnodiales;f__Mycosphaerellaceae;g__unidentified;s__unidentified; | 12 | 0.2411 | 1.1072 | 0.4170 | NS | 0.0210 | NS | 0.0247 | |
| unidentified | k__Fungi;p__Ascomycota;c__Dothideomycetes;o__Pleosporales;f__Phaeosphaeriaceae;g__Chaetosphaeronema;s__unidentified; | 12 | 0.1657 | 2.3941 | 0.9530 | NS | 0.0273 | NS | NS | |
| Arnium_arizonense | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Sordariales;f__Lasiosphaeriaceae;g__Arnium;s__Arnium_arizonense; | 12 | 0.1686 | 2.0909 | 0.8350 | NS | 0.0277 | NS | 0.0370 | |
| Chaetomium_grande | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Sordariales;f__Chaetomiaceae;g__Chaetomium;s__Chaetomium_grande; | 12 | 0.5311 | −1.3565 | 0.5441 | NS | 0.0283 | NS | NS | |
| unidentified | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Microascales;f__Halosphaeriaceae;g__unidentified;s__unidentified; | 12 | 0.2197 | 1.7837 | 0.7239 | NS | 0.0298 | NS | 0.0246 | |
| unidentified | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Branch06;f__unidentified;g__unidentified;s__unidentified; | 12 | 0.3074 | −1.8874 | 0.7661 | NS | 0.0298 | NS | NS | |
| Phialophora_mustea | k__Fungi;p__Ascomycota;c__Eurotiomycetes;o__Chaetothyriales;f__Herpotrichiellaceae;g__Phialophora;s__Phialophora_mustea; | 12 | 0.1589 | 2.3167 | 0.9793 | NS | 0.0357 | NS | 0.0404 | |
| unidentified | k__Fungi;p__Ascomycota;c__Eurotiomycetes;o__Onygenales;f__unidentified;g__unidentified;s__unidentified; | 12 | 0.2042 | 1.1675 | 0.4939 | NS | 0.0358 | NS | 0.0266 | |
| Exophiala_dermatitidis | k__Fungi;p__Ascomycota;c__Eurotiomycetes;o__Chaetothyriales;f__Herpotrichiellaceae;g__Exophiala;s__Exophiala_dermatitidis; | 12 | 0.1551 | 2.2601 | 0.9609 | NS | 0.0366 | NS | 0.0413 | |
| Dioszegia_fristingensis | k__Fungi;p__Basidiomycota;c__Tremellomycetes;o__Tremellales;f__Bulleribasidiaceae;g__Dioszegia;s__Dioszegia_fristingensis; | 12 | 0.1551 | 2.3156 | 0.9845 | NS | 0.0366 | NS | 0.0413 | |
| Pyrenochaeta_keratinophila | k__Fungi;p__Ascomycota;c__Dothideomycetes;o__Pleosporales;f__Cucurbitariaceae;g__Pyrenochaeta;s__Pyrenochaeta_keratinophila; | 12 | 0.1551 | 1.7072 | 0.7258 | NS | 0.0366 | NS | 0.0413 | |
| Exophiala_oligosperma | k__Fungi;p__Ascomycota;c__Eurotiomycetes;o__Chaetothyriales;f__Herpotrichiellaceae;g__Exophiala;s__Exophiala_oligosperma; | 12 | 0.1551 | 2.3156 | 0.9845 | NS | 0.0366 | NS | 0.0413 | |
| Alternaria_hungarica | k__Fungi;p__Ascomycota;c__Dothideomycetes;o__Pleosporales;f__Pleosporaceae;g__Alternaria;s__Alternaria_hungarica; | 12 | 0.1551 | 0.5971 | 0.2539 | NS | 0.0366 | NS | 0.0413 | |
| Peziza_buxea | k__Fungi;p__Ascomycota;c__Pezizomycetes;o__Pezizales;f__Pezizaceae;g__Peziza;s__Peziza_buxea; | 12 | 0.1551 | 2.3156 | 0.9845 | NS | 0.0366 | NS | 0.0413 | |
| Tetragoniomyces_uliginosus | k__Fungi;p__Basidiomycota;c__Tremellomycetes;o__Trichosporonales;f__Tetragoniomycetaceae;g__Tetragoniomyces;s__Tetragoniomyces_uliginosus; | 12 | 0.1551 | 2.3156 | 0.9845 | NS | 0.0366 | NS | 0.0413 | |
| unidentified | k__Fungi;p__Ascomycota;c__Dothideomycetes;o__Pleosporales;f__Leptosphaeriaceae;g__Leptosphaeria;s__unidentified; | 12 | 0.1551 | 2.3152 | 0.9843 | NS | 0.0366 | NS | 0.0413 | |
| unidentified | k__Fungi;p__Basidiomycota;c__Agaricomycetes;o__Agaricales;f__Omphalotaceae;g__Marasmiellus;s__unidentified; | 12 | 0.1551 | 2.3156 | 0.9845 | NS | 0.0366 | NS | 0.0413 | |
| Exophiala_phaeomuriformis | k__Fungi;p__Ascomycota;c__Eurotiomycetes;o__Chaetothyriales;f__Herpotrichiellaceae;g__Exophiala;s__Exophiala_phaeomuriformis; | 12 | 0.1551 | 2.3156 | 0.9845 | NS | 0.0366 | NS | 0.0413 | |
| unidentified | k__Fungi;p__Ascomycota;c__Pezizomycetes;o__Pezizales;f__Pyronemataceae;g__Trichophaea;s__unidentified; | 12 | 0.1551 | 2.3156 | 0.9845 | NS | 0.0366 | NS | 0.0413 | |
| Aspergillus_ochraceus | k__Fungi;p__Ascomycota;c__Eurotiomycetes;o__Eurotiales;f__Aspergillaceae;g__Aspergillus;s__Aspergillus_ochraceus; | 12 | 0.1551 | 1.4435 | 0.6137 | NS | 0.0366 | NS | 0.0413 | |
| Mortierella_zonata | k__Fungi;p__Mortierellomycota;c__Mortierellomycetes;o__Mortierellales;f__Mortierellaceae;g__Mortierella;s__Mortierella_zonata; | 12 | 0.1551 | 2.3156 | 0.9845 | NS | 0.0366 | NS | 0.0413 | |
| unidentified | k__Fungi;p__Ascomycota;c__Eurotiomycetes;o__Chaetothyriales;f__Herpotrichiellaceae;g__Phialophora;s__unidentified; | 12 | 0.1551 | 2.3156 | 0.9845 | NS | 0.0366 | NS | 0.0413 | |
| Microascus_albonigrescens | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Microascales;f__Microascaceae;g__Microascus;s__Microascus_albonigrescens; | 12 | 0.1551 | 2.3156 | 0.9845 | NS | 0.0366 | NS | 0.0413 | |
| Coniophora_olivacea | k__Fungi;p__Basidiomycota;c__Agaricomycetes;o__Boletales;f__Coniophoraceae;g__Coniophora;s__Coniophora_olivacea; | 12 | 0.1522 | 2.3083 | 0.9868 | NS | 0.0374 | NS | 0.0420 | |
| Mortierella_amoeboidea | k__Fungi;p__Mortierellomycota;c__Mortierellomycetes;o__Mortierellales;f__Mortierellaceae;g__Mortierella;s__Mortierella_amoeboidea; | 12 | 0.3633 | −1.2981 | 0.5824 | NS | 0.0457 | NS | 0.0549 | |
| unidentified | k__Fungi;p__Mortierellomycota;c__Mortierellomycetes;o__Mortierellales;f__Mortierellaceae;g__Mortierella;s__unidentified; | 12 | 0.3202 | −1.1002 | 0.5135 | NS | 0.0534 | NS | 0.0140 | |
| unidentified | k__Fungi;p__Basidiomycota;c__Wallemiomycetes;o__Wallemiales;f__Wallemiales_fam_Incertae_sedis;g__Wallemia;s__unidentified; | 12 | 0.6577 | −1.1381 | 0.5338 | NS | 0.0544 | NS | NS | |
| unidentified | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Branch06;f__unidentified;g__unidentified;s__unidentified; | 11 | 0.2450 | −1.9076 | 0.8332 | NS | NS | NS | 0.0428 | |
| unidentified | k__Fungi;p__Ascomycota;c__unidentified;o__unidentified;f__unidentified;g__unidentified;s__unidentified; | 11 | 0.2480 | −1.7915 | 0.7848 | NS | NS | NS | 0.0433 | |
| Plectosphaerella_cucumerina | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Glomerellales;f__Plectosphaerellaceae;g__Plectosphaerella;s__Plectosphaerella_cucumerina; | 11 | 0.1903 | −1.3601 | 0.6277 | NS | NS | NS | 0.0531 | |
| unidentified | k__Fungi;p__Ascomycota;c__Dothideomycetes;o__Pleosporales;f__Phaeosphaeriaceae;g__Chaetosphaeronema;s__unidentified; | 11 | 0.0922 | 2.4429 | 1.0354 | NS | NS | NS | 0.0379 | |
References
- Hand, T.W.; Vujkovic-Cvijin, I.; Ridaura, V.K.; Belkaid, Y. Linking the Microbiota, Chronic Disease, and the Immune System. Trends Endocrinol. Metab. 2016, 27, 831–843. [Google Scholar] [CrossRef]
- Sun, J.; Chang, E.B. Exploring gut microbes in human health and disease: Pushing the envelope. Genes Dis. 2014, 1, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The Microbiota-Gut-Brain Axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef]
- Petersen, C.; Round, J.L. Defining dysbiosis and its influence on host immunity and disease. Cell. Microbiol. 2014, 16, 1024–1033. [Google Scholar] [CrossRef]
- Lee, M.; Krishnamurthy, J.; Susi, A.; Sullivan, C.; Gorman, G.H.; Hisle-Gorman, E.; Erdie-Lalena, C.R.; Nylund, C.M. Association of Autism Spectrum Disorders and Inflammatory Bowel Disease. J. Autism Dev. Disord. 2018, 48, 1523–1529. [Google Scholar] [CrossRef] [PubMed]
- Garrett, V.D.; Brantley, P.J.; Jones, G.N.; McKnight, G.T. The relation between daily stress and Crohn’s disease. J. Behav. Med. 1991, 14, 87–96. [Google Scholar] [CrossRef]
- Neufeld, K.M.; Kang, N.; Bienenstock, J.; Foster, J.A. Reduced anxiety-like behavior and central neurochemical change in germ-free mice. Neurogastroenterol. Motil. 2010, 23, 255-e119. [Google Scholar] [CrossRef]
- Cryan, J.F. Stress and the Microbiota-Gut-Brain Axis. Can. J. Psychiatry 2016, 61, 201–203. [Google Scholar] [CrossRef] [PubMed]
- Fung, T.C.; A Olson, C.; Hsiao, T.C.F.C.A.O.E.Y. Interactions between the microbiota, immune and nervous systems in health and disease. Nat. Neurosci. 2017, 20, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, E.Y.; McBride, S.W.; Hsien, S.; Sharon, G.; Hyde, E.R.; McCue, T.; Codelli, J.A.; Chow, J.; Reisman, S.E.; Petrosino, J.F.; et al. Microbiota Modulate Behavioral and Physiological Abnormalities Associated with Neurodevelopmental Disorders. Cell 2013, 155, 1451–1463. [Google Scholar] [CrossRef]
- Vendrik, K.E.W.; Ooijevaar, R.E.; De Jong, P.R.C.; Laman, J.D.; Van Oosten, B.W.; Van Hilten, J.J.; Ducarmon, Q.R.; Keller, J.J.; Kuijper, E.J.; Contarino, M.F. Fecal Microbiota Transplantation in Neurological Disorders. Front. Cell. Infect. Microbiol. 2020, 10, 98. [Google Scholar] [CrossRef] [PubMed]
- Vuong, H.E.; Hsiao, E.Y. Emerging Roles for the Gut Microbiome in Autism Spectrum Disorder. Biol. Psychiatry 2017, 81, 411–423. [Google Scholar] [CrossRef] [PubMed]
- Moos, W.H.; Faller, U.V.; Harpp, D.N.; Kanara, I.; Pernokas, J.; Powers, W.R.; Steliou, K. Microbiota and Neurological Disorders: A Gut Feeling. BioResearch Open Access 2016, 5, 137–145. [Google Scholar] [CrossRef]
- Pascal, V.; Pozuelo, M.; Borruel, N.; Casellas, F.; Campos, D.; Santiago, A.; Martinez, X.; Varela, E.; Sarrabayrouse, G.; Machiels, K.; et al. A microbial signature for Crohn’s disease. Gut 2017, 66, 813–822. [Google Scholar] [CrossRef]
- Ghaisas, S.; Maher, J.; Kanthasamy, A. Gut microbiome in health and disease: Linking the microbiome-gut-brain axis and environmental factors in. Crime Justice Am. 2012, 417–421. [Google Scholar] [CrossRef]
- Enaud, R.; Vandenborght, L.-E.; Coron, N.; Bazin, T.; Prevel, R.; Schaeverbeke, T.; Berger, P.; Fayon, M.; Lamireau, T.; Delhaes, L. The Mycobiome: A Neglected Component in the Microbiota-Gut-Brain Axis. Microorg. 2018, 6, 22. [Google Scholar] [CrossRef]
- Limon, J.J.; Skalski, J.H.; Underhill, D.M. Commensal Fungi in Health and Disease. Cell Host Microbe 2017, 22, 156–165. [Google Scholar] [CrossRef]
- Nash, A.K.; Auchtung, T.A.; Wong, M.C.; Smith, D.P.; Gesell, J.R.; Ross, M.C.; Stewart, C.J.; Metcalf, G.A.; Muzny, D.M.; Gibbs, R.A.; et al. The gut mycobiome of the Human Microbiome Project healthy cohort. Microbiome 2017, 5, 153. [Google Scholar] [CrossRef]
- Li, C.; Li, Y.; Ding, C. The Role of Copper Homeostasis at the Host-Pathogen Axis: From Bacteria to Fungi. Int. J. Mol. Sci. 2019, 20, 175. [Google Scholar] [CrossRef] [PubMed]
- Oever, J.T.; Netea, M.G. The bacteriome-mycobiome interaction and antifungal host defense. Eur. J. Immunol. 2014, 44, 3182–3191. [Google Scholar] [CrossRef] [PubMed]
- Al Theyab, A.; Almutairi, T.; Al-Suwaidi, A.M.; Bendriss, G.; McVeigh, C.; Chaari, A. Epigenetic Effects of Gut Metabolites: Exploring the Path of Dietary Prevention of Type 1 Diabetes. Front. Nutr. 2020, 7. [Google Scholar] [CrossRef] [PubMed]
- Dalton, A.; Mermier, C.; Zuhl, M. Exercise influence on the microbiome–gut–brain axis. Gut Microbes 2019, 10, 555–568. [Google Scholar] [CrossRef] [PubMed]
- El Aidy, S.; Stilling, R.; Dinan, T.G.; Cryan, J.F. Microbiome to Brain: Unravelling the Multidirectional Axes of Communication. Adv. Exp. Med. Biol. 2016, 874, 301–336. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.P.; Easson, C.; Lyle, S.M.; Kapoor, R.; Donnelly, C.P.; Davidson, E.J.; Parikh, E.; Lopez, J.V.; Tartar, J.L. Gut microbiome diversity is associated with sleep physiology in humans. PLOS ONE 2019, 14, e0222394. [Google Scholar] [CrossRef]
- Haartsen, R.; Jones, E.J.; Johnson, M.H. Human brain development over the early years. Curr. Opin. Behav. Sci. 2016, 10, 149–154. [Google Scholar] [CrossRef]
- Yang, I.; Corwin, E.J.; Brennan, P.A.; Jordan, S.; Murphy, J.R.; Dunlop, A. The Infant Microbiome. Nurs. Res. 2016, 65, 76–88. [Google Scholar] [CrossRef]
- Rook, G.A.; Lowry, C.A.; Raison, C.L. Hygiene and other early childhood influences on the subsequent function of the immune system. Brain Res. 2015, 1617, 47–62. [Google Scholar] [CrossRef]
- Derrien, M.; Alvarez, A.-S.; de Vos, W.M. The Gut Microbiota in the First Decade of Life. Trends Microbiol. 2019, 27, 997–1010. [Google Scholar] [CrossRef]
- Bressan, P.; Kramer, P. Bread and Other Edible Agents of Mental Disease. Front. Hum. Neurosci. 2016, 10, 130. [Google Scholar] [CrossRef]
- Kok, C.R.; Hutkins, R. Yogurt and other fermented foods as sources of health-promoting bacteria. Nutr. Rev. 2018, 76, 4–15. [Google Scholar] [CrossRef]
- Abdellatif, B.; McVeigh, C.; Bendriss, G.; Chaari, A. The Promising Role of Probiotics in Managing the Altered Gut in Autism Spectrum Disorders. Int. J. Mol. Sci. 2020, 21, 4159. [Google Scholar] [CrossRef]
- Pais, P.; Almeida, V.; Yılmaz, M.; Teixeira, M.C. Saccharomyces boulardii: What Makes It Tick as Successful Probiotic? J. Fungi 2020, 6, 78. [Google Scholar] [CrossRef]
- Zou, R.; Wang, Y.; Duan, M.; Guo, M.; Zhang, Q.; Zheng, H. Dysbiosis of Gut Fungal Microbiota in Children with Autism Spectrum Disorders. J. Autism Dev. Disord. 2021, 51, 267–275. [Google Scholar] [CrossRef]
- Łukasik, J.; Patro-Gołąb, B.; Horvath, A.; Baron, R.; Szajewska, H. Sawanti Working Group Early Life Exposure to Antibiotics and Autism Spectrum Disorders: A Systematic Review. J. Autism Dev. Disord. 2019, 49, 3866–3876. [Google Scholar] [CrossRef]
- A Slob, E.M.; Brew, B.K.; Vijverberg, S.J.H.; Dijs, T.; Beijsterveldt, C.E.M.V.; Koppelman, G.H.; Bartels, M.; Dolan, C.V.; Larsson, H.; Lundström, S.; et al. Early-life antibiotic use and risk of attention-deficit hyperactivity disorder and autism spectrum disorder: Results of a discordant twin study. Int. J. Epidemiol. 2020, dyaa168. [Google Scholar] [CrossRef]
- Heijtz, R.D. Fetal, neonatal, and infant microbiome: Perturbations and subsequent effects on brain development and behavior. Semin. Fetal Neonatal Med. 2016, 21, 410–417. [Google Scholar] [CrossRef]
- Hamad, A.F.; Alessi-Severini, S.; Mahmud, S.M.; Brownell, M.; Kuo, I.F. Early childhood antibiotics use and autism spectrum disorders: A population-based cohort study. Int. J. Epidemiol. 2018, 47, 1497–1506. [Google Scholar] [CrossRef]
- Jaureguiberry, M.S.; Venturino, A. Nutritional and environmental contributions to Autism Spectrum Disorders: Focus on nutrigenomics as complementary therapy. Int. J. Vitam. Nutr. Res. 2020, 1–19. [Google Scholar] [CrossRef]
- Alshaban, F. Prevalence of Autism Spectrum Disorders in Qatar. Qatar Found. Annu. Res. Forum Proc. 2011, 2011, BMP64. [Google Scholar] [CrossRef]
- Bendriss, G.; Al-Ali, D.; Shafiq, A.; Laswi, I.; Mhaimeed, N.; Salameh, M.; Burney, Z.; Pillai, K.; Chaari, A.; Zakaria, D.; et al. Targeting the gut microbiome: A brief report on the awareness, practice, and readiness to engage in clinical interventions in Qatar. Qatar Med. J. 2021, 2020, 47. [Google Scholar] [CrossRef]
- Kuehbacher, T.; Rehman, A.; Lepage, P.; Hellmig, S.; Fölsch, U.R.; Schreiber, S.; Ott, S.J. Intestinal TM7 bacterial phylogenies in active inflammatory bowel disease. J. Med. Microbiol. 2008, 57, 1569–1576. [Google Scholar] [CrossRef]
- Fattorusso, A.; Di Genova, L.; Dell’Isola, G.B.; Mencaroni, E.; Esposito, S. Autism Spectrum Disorders and the Gut Microbiota. Nutr. 2019, 11, 521. [Google Scholar] [CrossRef]
- Ma, B.; Liang, J.; Dai, M.; Wang, J.; Luo, J.; Zhang, Z.; Jing, J. Altered Gut Microbiota in Chinese Children With Autism Spectrum Disorders. Front. Cell. Infect. Microbiol. 2019, 9, 40. [Google Scholar] [CrossRef]
- Gong, D.; Gong, X.; Wang, L.; Yu, X.; Dong, Q. Involvement of Reduced Microbial Diversity in Inflammatory Bowel Disease. Gastroenterol. Res. Pr. 2016, 2016, 6951091. [Google Scholar] [CrossRef]
- Liang, X.; Li, H.; Tian, G.; Li, S. Dynamic microbe and molecule networks in a mouse model of colitis-associated colorectal cancer. Sci. Rep. 2014, 4, 4985. [Google Scholar] [CrossRef]
- Xu, M.; Xu, X.; Li, J.; Li, F. Association Between Gut Microbiota and Autism Spectrum Disorder: A Systematic Review and Meta-Analysis. Front. Psychiatry 2019, 10, 473. [Google Scholar] [CrossRef]
- Torres, J.; Hu, J.; Seki, A.; Eisele, C.; Nair, N.; Huang, R.; Tarassishin, L.; Jharap, B.; Cote-Daigneault, J.; Mao, Q.; et al. Infants born to mothers with IBD present with altered gut microbiome that transfers abnormalities of the adaptive immune system to germ-free mice. Gut 2019, 69, 42–51. [Google Scholar] [CrossRef]
- Russo, A.; Krigsman, A.; Jepson, B.; Wakefield, A. Generalized Autoimmunity of ANCA and ASCA Related to Severity of Disease in Autistic Children with GI Disease. Immunol. Immunogenet. Insights 2009, 1, III.S3073. [Google Scholar] [CrossRef]
- Brock, P.M.; Döring, H.; Bidartondo, M.I. How to know unknown fungi: The role of a herbarium. New Phytol. 2008, 181, 719–724. [Google Scholar] [CrossRef]
- Dollive, S.; Chen, Y.-Y.; Grunberg, S.; Bittinger, K.; Hoffmann, C.; Vandivier, L.; Cuff, C.; Lewis, J.D.; Wu, G.D.; Bushman, F.D. Fungi of the Murine Gut: Episodic Variation and Proliferation during Antibiotic Treatment. PLOS ONE 2013, 8, e71806. [Google Scholar] [CrossRef]
- Noverr, M.C.; Noggle, R.M.; Toews, G.B.; Huffnagle, G.B. Role of Antibiotics and Fungal Microbiota in Driving Pulmonary Allergic Responses. Infect. Immun. 2004, 72, 4996–5003. [Google Scholar] [CrossRef]
- Rapid Range Shifts of Species Associated with High Levels of Climate Warming—European Environment Agency. 2011. Available online: https://value.eea.europa.eu/data-and-maps/indicators/distribution-of-animal-species-1/chen-et-al.-2011-rapid (accessed on 27 February 2021).
- Gouba, N.; Raoult, D.; Drancourt, M. Plant and Fungal Diversity in Gut Microbiota as Revealed by Molecular and Culture Investigations. PLOS ONE 2013, 8, e59474. [Google Scholar] [CrossRef] [PubMed]
- Morton, J.T.; Marotz, C.; Washburne, A.; Silverman, J.; Zaramela, L.S.; Edlund, A.; Zengler, K.; Knight, R. Establishing microbial composition measurement standards with reference frames. Nat. Commun. 2019, 10, 1–11. [Google Scholar] [CrossRef]
- Laue, H.E.; Korrick, S.A.; Baker, E.R.; Karagas, M.R.; Madan, J.C. Prospective associations of the infant gut microbiome and microbial function with social behaviors related to autism at age 3 years. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Iebba, V.; Aloi, M.; Civitelli, F.; Cucchiara, S. Gut Microbiota and Pediatric Disease. Dig. Dis. 2011, 29, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Christophersen, C.T.; Sorich, M.J.; Gerber, J.P.; Angley, M.T.; Conlon, M.A. Low Relative Abundances of the Mucolytic Bacterium Akkermansia muciniphila and Bifidobacterium spp. in Feces of Children with Autism. Appl. Environ. Microbiol. 2011, 77, 6718–6721. [Google Scholar] [CrossRef]
- Bäckhed, F.; Fraser, C.M.; Ringel, Y.; Sanders, M.E.; Sartor, R.B.; Sherman, P.M.; Versalovic, J.; Young, V.; Finlay, B.B. Defining a Healthy Human Gut Microbiome: Current Concepts, Future Directions, and Clinical Applications. Cell Host Microbe 2012, 12, 611–622. [Google Scholar] [CrossRef]
- Koenig, J.E.; Spor, A.; Scalfone, N.; Fricker, A.D.; Stombaugh, J.; Knight, R.; Angenent, L.T.; Ley, R.E. Succession of microbial consortia in the developing infant gut microbiome. Proc. Natl. Acad. Sci. USA 2010, 108, 4578–4585. [Google Scholar] [CrossRef] [PubMed]
- Yatsunenko, T.; Rey, F.E.; Manary, M.J.; Trehan, I.; Dominguez-Bello, M.G.; Contreras, M.; Magris, M.; Hidalgo, G.; Baldassano, R.N.; Anokhin, A.P.; et al. Human gut microbiome viewed across age and geography. Nature 2012, 486, 222–227. [Google Scholar] [CrossRef]
- Nagpal, R.; Mainali, R.; Ahmadi, S.; Wang, S.; Singh, R.; Kavanagh, K.; Kitzman, D.W.; Kushugulova, A.; Marotta, F.; Yadav, H. Gut microbiome and aging: Physiological and mechanistic insights. Nutr. Health Aging 2018, 4, 267–285. [Google Scholar] [CrossRef]
- DeJong, E.N.; Surette, M.G.; Bowdish, D.M. The Gut Microbiota and Unhealthy Aging: Disentangling Cause from Consequence. Cell Host Microbe 2020, 28, 180–189. [Google Scholar] [CrossRef]
- Shen, J.; Zuo, Z.-X.; Mao, A.-P. Effect of Probiotics on Inducing Remission and Maintaining Therapy in Ulcerative Colitis, Crohnʼs Disease, and Pouchitis. Inflamm. Bowel Dis. 2014, 20, 21–35. [Google Scholar] [CrossRef]
- Wong, J.M.W.; De Souza, R.; Kendall, C.W.C.; Emam, A.; Jenkins, D.J.A. Colonic Health: Fermentation and Short Chain Fatty Acids. J. Clin. Gastroenterol. 2006, 40, 235–243. [Google Scholar] [CrossRef]
- Asghari, G.; Farhadnejad, H.; Teymoori, F.; Mirmiran, P.; Tohidi, M.; Azizi, F. High dietary intake of branched-chain amino acids is associated with an increased risk of insulin resistance in adults. J. Diabetes 2017, 10, 357–364. [Google Scholar] [CrossRef]
- Kandeel, W.A.; Meguid, N.A.; Bjørklund, G.; Eid, E.M.; Farid, M.; Mohamed, S.K.; Wakeel, K.E.; Chirumbolo, S.; Elsaeid, A.; Hammad, D.Y. Impact of Clostridium Bacteria in Children with Autism Spectrum Disorder and Their Anthropometric Measurements. J. Mol. Neurosci. 2020, 70, 897–907. [Google Scholar] [CrossRef]
- Suez, J.; Korem, T.; Zeevi, D.; Zilberman-Schapira, G.; Thaiss, C.A.; Maza, O.; Israeli, D.; Zmora, N.; Gilad, S.; Weinberger, A.; et al. Artificial sweeteners induce glucose intolerance by altering the gut microbiota. Nat. Cell Biol. 2014, 514, 181–186. [Google Scholar] [CrossRef]
- Zmora, N.; Suez, J.; Elinav, E. You are what you eat: Diet, health and the gut microbiota. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 35–56. [Google Scholar] [CrossRef]
- Miclotte, L.; Van De Wiele, T. Food processing, gut microbiota and the globesity problem. Crit. Rev. Food Sci. Nutr. 2019, 60, 1769–1782. [Google Scholar] [CrossRef]
- Blum, W.E.; Zechmeister-Boltenstern, S.; Keiblinger, K.M. Does Soil Contribute to the Human Gut Microbiome? Microorganisms 2019, 7, 287. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, F.; Ding, X.; Wu, G.; Lam, Y.Y.; Wang, X.; Fu, H.; Xue, X.; Lu, C.; Ma, J.; et al. Gut bacteria selectively promoted by dietary fibers alleviate type 2 diabetes. Science 2018, 359, 1151–1156. [Google Scholar] [CrossRef]
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef] [PubMed]
- Peleg, A.Y.; Hogan, D.A.; Mylonakis, E. Medically important bacterial–fungal interactions. Nat. Rev. Genet. 2010, 8, 340–349. [Google Scholar] [CrossRef] [PubMed]
- Gaziano, R.; Sabbatini, S.; Roselletti, E.; Perito, S.; Monari, C. Saccharomyces cerevisiae-Based Probiotics as Novel Antimicrobial Agents to Prevent and Treat Vaginal Infections. Front. Microbiol. 2020, 11, 718. [Google Scholar] [CrossRef] [PubMed]
- Martins, F.S.; Nardi, R.M.D.; Arantes, R.M.E.; Rosa, C.A.; Neves, M.J.; Nicoli, J.R. Screening of yeasts as probiotic based on capacities to colonize the gastrointestinal tract and to protect against enteropathogen challenge in mice. J. Gen. Appl. Microbiol. 2005, 51, 83–92. [Google Scholar] [CrossRef]
- Kühle, A.V.D.A.; Jespersen, L. The Taxonomic Position of Saccharomyces boulardii as Evaluated by Sequence Analysis of the D1/D2 Domain of 26S rDNA, the ITS1-5.8S rDNA-ITS2 Region and the Mitochondrial Cytochrome-c Oxidase II Gene. Syst. Appl. Microbiol. 2003, 26, 564–571. [Google Scholar] [CrossRef]
- Krajmalnik-Brown, R.; Lozupone, C.; Kang, D.-W.; Adams, J.B. Gut bacteria in children with autism spectrum disorders: Challenges and promise of studying how a complex community influences a complex disease. Microb. Ecol. Health Dis. 2015, 26, 26914. [Google Scholar] [CrossRef]
- Ercolini, D.; Francavilla, R.; Vannini, L.; De Filippis, F.; Capriati, T.; Di Cagno, R.; Iacono, G.; De Angelis, M.; Gobbetti, M. From an imbalance to a new imbalance: Italian-style gluten-free diet alters the salivary microbiota and metabolome of African celiac children. Sci. Rep. 2015, 5, 18571. [Google Scholar] [CrossRef]
- Son, J.S.; Zheng, L.J.; Rowehl, L.M.; Tian, X.; Zhang, Y.; Zhu, W.; Litcher-Kelly, L.; Gadow, K.D.; Gathungu, G.; Robertson, C.E.; et al. Comparison of Fecal Microbiota in Children with Autism Spectrum Disorders and Neurotypical Siblings in the Simons Simplex Collection. PLOS ONE 2015, 10, e0137725. [Google Scholar] [CrossRef]
- Williams, B.A.; Grant, L.J.; Gidley, M.J.; Mikkelsen, D. Gut Fermentation of Dietary Fibres: Physico-Chemistry of Plant Cell Walls and Implications for Health. Int. J. Mol. Sci. 2017, 18, 2203. [Google Scholar] [CrossRef]
- Kang, D.-W.; Park, J.G.; Ilhan, Z.E.; Wallstrom, G.; LaBaer, J.; Adams, J.B.; Krajmalnik-Brown, R. Reduced Incidence of Prevotella and Other Fermenters in Intestinal Microflora of Autistic Children. PLOS ONE 2013, 8, e68322. [Google Scholar] [CrossRef]
- Esaiassen, E.; Fjalstad, J.W.; Juvet, L.K.; Anker, J.N.V.D.; Klingenberg, C. Antibiotic exposure in neonates and early adverse outcomes: A systematic review and meta-analysis. J. Antimicrob. Chemother. 2017, 72, 1858–1870. [Google Scholar] [CrossRef]
- E Dawson-Hahn, E.; Mickan, S.; Onakpoya, I.; Roberts, N.; Kronman, M.; Butler, C.C.; Thompson, M.J. Short-course versus long-course oral antibiotic treatment for infections treated in outpatient settings: A review of systematic reviews. Fam. Pr. 2017, 34, 511–519. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Chung, J.; Battaglia, T.; Henderson, N.; Jay, M.; Li, H.; Lieber, A.D.; Wu, F.; Perez-Perez, G.I.; Chen, Y.; et al. Antibiotics, birth mode, and diet shape microbiome maturation during early life. Sci. Transl. Med. 2016, 8, 343ra82. [Google Scholar] [CrossRef]
- Schwartz, D.J.; Langdon, A.E.; Dantas, G. Understanding the impact of antibiotic perturbation on the human microbiome. Genome Med. 2020, 12, 1–12. [Google Scholar] [CrossRef]
- Liu, F.; Li, J.; Wu, F.; Zheng, H.; Peng, Q.; Zhou, H. Altered composition and function of intestinal microbiota in autism spectrum disorders: A systematic review. Transl. Psychiatry 2019, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Stojanov, S.; Berlec, A.; Štrukelj, B. The Influence of Probiotics on the Firmicutes/Bacteroidetes Ratio in the Treatment of Obesity and Inflammatory Bowel disease. Microorganisms 2020, 8, 1715. [Google Scholar] [CrossRef] [PubMed]
- Strati, F.; Cavalieri, D.; Albanese, D.; De Felice, C.; Donati, C.; Hayek, J.; Jousson, O.; Leoncini, S.; Renzi, D.; Calabrò, A.; et al. New evidences on the altered gut microbiota in autism spectrum disorders. Microbiome 2017, 5, 1–11. [Google Scholar] [CrossRef]
- Finegold, S.M.; Dowd, S.E.; Gontcharova, V.; Liu, C.; Henley, K.E.; Wolcott, R.D.; Youn, E.; Summanen, P.H.; Granpeesheh, D.; Dixon, D.; et al. Pyrosequencing study of fecal microflora of autistic and control children. Anaerobe 2010, 16, 444–453. [Google Scholar] [CrossRef]
- Li, Q.; Han, Y.; Dy, A.B.C.; Hagerman, R.J. The Gut Microbiota and Autism Spectrum Disorders. Front. Cell. Neurosci. 2017, 11, 120. [Google Scholar] [CrossRef] [PubMed]
- Fernández, J.; Redondo-Blanco, S.; Gutiérrez-Del-Río, I.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Colon microbiota fermentation of dietary prebiotics towards short-chain fatty acids and their roles as anti-inflammatory and antitumour agents: A review. J. Funct. Foods 2016, 25, 511–522. [Google Scholar] [CrossRef]
- Topping, D.L.; Clifton, P.M. Short-Chain Fatty Acids and Human Colonic Function: Roles of Resistant Starch and Nonstarch Polysaccharides. Physiol. Rev. 2001, 81, 1031–1064. [Google Scholar] [CrossRef]
- Dalile, B.; Van Oudenhove, L.; Vervliet, B.; Verbeke, K. The role of short-chain fatty acids in microbiota—Gut—Brain communication. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 461–478. [Google Scholar] [CrossRef]
- Macfabe, D.F. Short-chain fatty acid fermentation products of the gut microbiome: Implications in autism spectrum disorders. Microb. Ecol. Heal. Dis. 2012, 23, 23. [Google Scholar] [CrossRef]
- Binda, C.; Lopetuso, L.R.; Rizzatti, G.; Gibiino, G.; Cennamo, V.; Gasbarrini, A. Actinobacteria: A relevant minority for the maintenance of gut homeostasis. Dig. Liver Dis. 2018, 50, 421–428. [Google Scholar] [CrossRef]
- Cross, K.L.; Campbell, J.H.; Balachandran, M.; Campbell, A.G.; Cooper, S.J.; Griffen, A.; Heaton, M.; Joshi, S.; Klingeman, D.; Leys, E.; et al. Targeted isolation and cultivation of uncultivated bacteria by reverse genomics. Nat. Biotechnol. 2019, 37, 1314–1321. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Sun, T.; Wu, F.; Li, F.; Liu, Y.; Li, W.; Dai, N.; Tan, L.; Li, T.; Song, Y. Correlation of gut microbiota and neurotransmitters in a rat model of post-traumatic stress disorder. J. Tradit. Chin. Med Sci. 2020, 7, 375–385. [Google Scholar] [CrossRef]
- Bojović, K.; Bajić, S.S.; Milutinović, D.V.; Tomić, M.; Golić, N.; Tolinački, M. Gut Microbiota Dysbiosis Associated With Altered Production of Short Chain Fatty Acids in Children with Neurodevelopmental Disorders. Front. Cell. Infect. Microbiol. 2020, 10, 223. [Google Scholar] [CrossRef]
- Valles-Colomer, M.; Falony, G.; Darzi, Y.; Tigchelaar, E.F.; Wang, J.; Tito, R.Y.; Schiweck, C.; Kurilshikov, A.; Joossens, M.; Wijmenga, C.; et al. The neuroactive potential of the human gut microbiota in quality of life and depression. Nat. Microbiol. 2019, 4, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Kobliner, V.; Mumper, E.; Baker, S.M. Reduction in obsessive compulsive disorder and self-injurious behavior with sac-charomyces boulardii in a child with autism: A case report. Integr. Med. 2018, 17, 38–41. [Google Scholar]
- Pandey, K.R.; Naik, S.R.; Vakil, B.V. Probiotics, prebiotics and synbiotics—A review. J. Food Sci. Technol. 2015, 52, 7577–7587. [Google Scholar] [CrossRef] [PubMed]
- Backhed, F. Host-Bacterial Mutualism in the Human Intestine. Science 2005, 307, 1915–1920. [Google Scholar] [CrossRef]
- Atarashi, K.; Tanoue, T.; Shima, T.; Imaoka, A.; Kuwahara, T.; Momose, Y.; Cheng, G.; Yamasaki, S.; Saito, T.; Ohba, Y.; et al. Induction of colonic regulatory T cells by indigenous clostridium species. Science 2011, 331, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.J.; Boushey, H.A. The microbiome in asthma. J. Allergy Clin. Immunol. 2015, 135, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Iliev, I.D.; Leonardi, I.D.I.I. Fungal dysbiosis: Immunity and interactions at mucosal barriers. Nat. Rev. Immunol. 2017, 17, 635–646. [Google Scholar] [CrossRef] [PubMed]
- Pascal, M.; Perez-Gordo, M.; Caballero, T.; Escribese, M.M.; Longo, M.N.L.; Luengo, O.; Manso, L.; Matheu, V.; Seoane, E.; Zamorano, M.; et al. Microbiome and Allergic Diseases. Front. Immunol. 2018, 9, 1584. [Google Scholar] [CrossRef] [PubMed]
- Demirci, M.; Tokman, H.; Uysal, H.; Demiryas, S.; Karakullukcu, A.; Saribas, S.; Cokugras, H.; Kocazeybek, B. Reduced Akkermansia muciniphila and Faecalibacterium prausnitzii levels in the gut microbiota of children with allergic asthma. Allergol. Immunopathol. 2019, 47, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Enstrom, A.M.; A Van De Water, J.; Ashwood, P. Autoimmunity in autism. Curr. Opin. Investig. Drugs 2009, 10, 463–473. [Google Scholar]
- McLean, M.H.; Dieguez, D.; Miller, L.M.; A Young, H. Does the microbiota play a role in the pathogenesis of autoimmune diseases? Gut 2015, 64, 332–341. [Google Scholar] [CrossRef] [PubMed]




| CTRL | NDD | GID | TOTAL | |
|---|---|---|---|---|
| Gender N (%) | ||||
| Male | 10 (37) | 11 (73) | 5 (38) | 26 (47) |
| Female | 17 (63) | 4 (27) | 8 (62) | 29 (52) |
| Total | 27 (100) | 15 (100) | 13 (100) | 55 (100) |
| Age N (%) | ||||
| 3 to 18 years | 12 (44) | 14 (93) | 3 (23) | 29 (52) |
| 19–60 years | 15 (56) | 1 (7) | 10 (77) | 26 (47) |
| Total | 27 (100) | 15 (100) | 13 (100) | 55 (100) |
| Ethnicity N (%) | ||||
| Middle Eastern | 8 (30) | 6 (40) | 1 (8) | 15 (27) |
| Caucasian | 9 (33) | 2 (13) | 9 (69) | 20 (11) |
| Indian | 1 (4) | 3 (20) | 1 (8) | 5 (9) |
| Asian | 2 (7) | 2 (13) | 0 (0) | 4 (7) |
| North African | 2 (7) | 0 (0) | 2 (15) | 4 (7) |
| Black African | 1 (4) | 1 (7) | 0 (0) | 2 (3) |
| Body mass index N (%) | ||||
| BMI ≥ 30 | 4 (15) | 0 (0) | 2 (15) | 6 (11) |
| BMI < 30 | 22 (81) | 11 (73) | 11 (85) | 44 (80) |
| Probiotic intake N (%) | ||||
| Regular intake | 12 (44) | 6 (40) | 7 (54) | 23 (42) |
| No regular intake | 15 (56) | 9 (60) | 6 (46) | 30 (55) |
| Diet in processed carbohydrates N (%) | ||||
| Medium to high | 22 (81) | 15 (100) | 9 (69) | 46 (84) |
| Low | 5 (19) | 0 | 4 (31) | 9 (16) |
| Recurrent antibiotic use before 3Y N (%) | ||||
| Yes | 7 | 10 | 6 | 23 (42) |
| No | 20 | 5 | 5 | 30 (55) |
| Phyla Ratio | Basic Covariates + Probiotics and Diet | ||||||
|---|---|---|---|---|---|---|---|
| DF | R-Squared | Beta | SEbeta | p-Value | |||
| NDD compared to CTRL | Proteobacteria/Cyanobacteria | ↑ | 19 | 0.222 | 3413.585 | 1182.735 | 0.009 |
| Firmicutes/Actinobacteria | ↓ | 29 | 0.253 | −170.525 | 74.769 | 0.030 | |
| Firmicutes/Bacteroidetes | ↓ | 29 | 0.094 | −0.721 | 0.345 | 0.045 | |
| Proteobacteria/Verrucomicrobia | ↑ | 27 | 0.179 | 848.553 | 404.793 | 0.046 | |
| GID compared to CTRL | Firmicutes/Verrucomicrobia | ↑ | 28 | 0.182 | 9628.261 | 3295.069 | 0.007 |
| Cyanobacteria/Verrucomicrobia | ↑ | 28 | 0.159 | 7.397 | 2.630 | 0.009 | |
| Actinobacteria/Verrucomicrobia | ↑ | 28 | 0.030 | 1278.024 | 576.498 | 0.035 | |
| TM7/Actinobacteria | ↓ | 31 | 0.113 | −0.009 | 0.004 | 0.041 | |
| TM7/Firmicutes | ↓ | 31 | 0.066 | 0.000 | 0.000 | 0.044 | |
| NDD compared to GID | TM7/Actinobacteria | ↓ | 42 | 0.125 | −0.008 | 0.003 | 0.018 |
| Firmicutes/Verrucomicrobia | ↑ | 38 | 0.109 | 6035.704 | 2508.904 | 0.021 | |
| Proteobacteria/Verrucomicrobia | ↑ | 38 | 0.193 | 756.313 | 320.375 | 0.023 | |
| Firmicutes/Actinobacteria | ↓ | 42 | 0.229 | −106.975 | 49.164 | 0.035 | |
| Cyanobacteria/Verrucomicrobia | ↑ | 38 | 0.070 | 4.239 | 1.995 | 0.040 | |
| Level | Name | Basic Covariates | Adding Probiotics | Adding Diet | Adding Probiotics and Diet | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| DF | R-Squared | Beta | SEbeta | p-Value | p-Value 1 | p-Value 2 | p-Value 3 | |||
| Order | ↑ | Enterobacteriales | 32 | 0.170983 | 1.43611 | 0.447029 | 0.002996 | 0.002745 | 0.004335 | 0.003967 |
| ↑ | RF32 | 32 | 0.100373 | 0.257666 | 0.097869 | 0.012931 | 0.017538 | 0.017723 | 0.023618 | |
| Family | ↑ | Enterobacteriaceae | 32 | 0.170983 | 1.43611 | 0.447029 | 0.002996 | 0.002745 | 0.004335 | 0.012903 |
| ↑ | Enterococcaceae | 32 | 0.142559 | 1.225977 | 0.450581 | 0.010441 | 0.00933 | 0.014408 | NS | |
| Genus | ↑ | SMB53 | 32 | 0.298415 | 0.75418 | 0.21188 | 0.001185 | 0.001722 | 0.001206 | 0.001785 |
| ↑ | Escherichia | 32 | 0.122884 | 1.231856 | 0.429424 | 0.007242 | 0.008244 | 0.009616 | 0.010944 | |
| ↑ | Clostridium | 32 | 0.193332 | 1.244652 | 0.434901 | 0.007365 | 0.009685 | 0.010454 | 0.013531 | |
| ↑ | Butyricicoccus | 32 | 0.137802 | 1.193238 | 0.431852 | 0.009413 | 0.009851 | 0.01305 | 0.013604 | |
| ↑ | Enterococcus | 32 | 0.142559 | 1.225977 | 0.450581 | 0.010441 | 0.00933 | 0.014408 | 0.012903 | |
| ↑ | Lactococcus | 32 | 0.138711 | 0.12109 | 0.049673 | 0.020514 | 0.030686 | 0.019488 | 0.02914 | |
| ↑ | Veillonella | 32 | 0.066232 | 1.126486 | 0.464704 | 0.021175 | 0.029099 | 0.029006 | 0.039101 | |
| ↓ | Coprococcus | 32 | 0.10669 | −0.60313 | 0.29107 | 0.046392 | 0.046974 | NS | 0.003967 | |
| Species | ↑ | coli | 32 | 0.122884 | 1.231856 | 0.429424 | 0.007242 | 0.008244 | 0.009616 | 0.010944 |
| ↑ | pullicaecorum | 32 | 0.137802 | 1.193238 | 0.431852 | 0.009413 | 0.009851 | 0.01305 | 0.013604 | |
| ↑ | dispar | 32 | 0.050691 | 1.005875 | 0.471401 | 0.040622 | NS | NS | NS | |
| ↑ | bifidum | 32 | 0.034055 | 0.899072 | 0.439749 | 0.049203 | NS | NS | NS | |
| Level | Name | Basic Covariates | Adding Probiotics | Adding Diet | Adding Probiotics and Diet | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| DF | R-Squared | Beta | SEbeta | p-Value | p-Value 1 | p-Value 2 | p-Value 3 | |||
| Phylum | ↓ | Firmicutes | 32 | 0.077031 | −0.95357 | 0.366404 | 0.013907 | 0.014174 | 0.007765 | 0.007917 |
| Order | ↑ | Enterobacteriales | 32 | 0.168988 | 1.447609 | 0.445448 | 0.002717 | 0.002453 | 0.003935 | 0.003554 |
| ↓ | Clostridiales | 32 | 0.115446 | −1.03299 | 0.356563 | 0.006743 | 0.007301 | 0.003770 | 0.004081 | |
| ↑ | RF32 | 32 | 0.052137 | 0.185216 | 0.090321 | 0.048567 | NS | NS | NS | |
| Family | ↑ | Enterobacteriaceae | 32 | 0.168988 | 1.447609 | 0.445448 | 0.002717 | 0.002453 | 0.003935 | 0.003554 |
| ↑ | Brevibacteriaceae | 32 | 0.133259 | 1.249522 | 0.433006 | 0.006939 | 0.009815 | 0.008963 | 0.012486 | |
| ↑ | Enterococcaceae | 32 | 0.144581 | 1.225499 | 0.447687 | 0.010027 | 0.009193 | 0.013925 | 0.012765 | |
| ↓ | Mogibacteriaceae | 32 | 0.065234 | −0.80553 | 0.385277 | 0.044577 | NS | NS | NS | |
| ↑ | Corynebacteriaceae | 32 | 0.02451 | −0.5357 | 0.3331 | NS | NS | NS | 0.041973 | |
| Genus | ↑ | SMB53 | 32 | 0.225957 | 0.647016 | 0.220494 | 0.006137 | 0.008254 | 0.006756 | 0.009169 |
| ↑ | Brevibacterium | 32 | 0.133259 | 1.249522 | 0.433006 | 0.006939 | 0.009815 | 0.008963 | 0.012486 | |
| ↑ | Enterococcus | 32 | 0.144581 | 1.225499 | 0.447687 | 0.010027 | 0.009193 | 0.013925 | 0.012765 | |
| ↑ | Clostridium | 32 | 0.181039 | 1.147432 | 0.433232 | 0.012448 | 0.016721 | 0.017419 | 0.023002 | |
| ↑ | Escherichia | 32 | 0.091324 | 1.123838 | 0.436121 | 0.014785 | 0.017222 | 0.018506 | 0.021551 | |
| ↑ | Butyricicoccus | 32 | 0.111908 | 1.060691 | 0.413567 | 0.015219 | 0.016210 | 0.020536 | 0.021816 | |
| ↑ | Veillonella | 32 | 0.055591 | 1.079019 | 0.461371 | 0.025758 | 0.035185 | 0.035173 | 0.047124 | |
| ↓ | Coprococcus | 32 | 0.112285 | −0.70915 | 0.316395 | 0.032068 | 0.032769 | 0.036046 | 0.036777 | |
| ↓ | Odoribacter | 32 | 0.1039 | −0.880 | 0.4329 | NS | 0.049563 | 0.047686 | 0.047292 | |
| Species | ↑ | aureum | 32 | 0.133259 | 1.249522 | 0.433006 | 0.006939 | 0.009815 | 0.008963 | 0.012486 |
| ↑ | coli | 32 | 0.091324 | 1.123838 | 0.436121 | 0.014785 | 0.017222 | 0.018506 | 0.021551 | |
| ↑ | pullicaecorum | 32 | 0.111908 | 1.060691 | 0.413567 | 0.015219 | 0.016210 | 0.020536 | 0.021816 | |
| ↓ | dolichum | 32 | 0.102217 | -0.11802 | 0.054746 | 0.038727 | NS | 0.043557 | NS | |
| ↑ | dispar | 32 | 0.045073 | 0.989858 | 0.465437 | 0.041250 | NS | NS | NS | |
| ↑ | faecis | 32 | 0.031296 | 0.477281 | 0.229260 | 0.045439 | 0.037505 | NS | 0.045605 | |
| ↑ | bifidum | 32 | 0.038356 | 0.897201 | 0.434263 | 0.047000 | NS | NS | NS | |
| Level | Name | Basic Covariates | Adding Probiotics | Adding Diet | Adding Probiotics and Diet | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| DF | R-Squared | Beta | Sebeta | p-Value | p-Value 1 | p-Value 2 | p-Value 3 | |||
| Family | ↓ | Odoribacteraceae | 34 | 0.04502157 | −0.7451711 | 0.35897333 | 0.04554079 | NS | 0.04884609 | NS |
| Genus | ↑ | Anaerotruncus | 34 | 0.18021437 | 0.54957278 | 0.1741761 | 0.00334826 | 0.00463763 | 0.00201844 | 0.00288391 |
| ↓ | Butyricimonas | 34 | 0.14848867 | −0.6716181 | 0.27245144 | 0.01890498 | 0.02584356 | 0.02047021 | 0.02777944 | |
| Species | ↓ | catus | 34 | 0.082374 | −0.8869353 | 0.35892387 | 0.01863917 | 0.01932181 | 0.01999275 | 0.0205811 |
| ↑ | luteciae | 34 | 0.07544368 | 0.22614282 | 0.11062371 | 0.04873506 | NS | NS | NS | |
| Level | Name | Basic Covariates | Adding Probiotics | Adding Diet | Adding Probiotics and Diet | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| DF | R-Squared | Beta | Sebeta | p-Value | p-Value 1 | p-Value 2 | p-Value 3 | |||
| no significant difference in any taxa abundances between NDD and GID at Phylum, Class, Family, Order levels across all 4 models | ||||||||||
| Genus | ↑ | SMB53 | 45 | 0.0566 | 0.6860 | 0.2891 | 0.0220 | 0.0297 | 0.0234 | 0.0317 |
| ↓ | Odoribacter | 45 | 0.1602 | −0.5429 | 0.2629 | 0.0447 | 0.0417 | 0.0473 | 0.0441 | |
| ↑ | Anaerotruncus | 45 | 0.0435 | 0.5814 | 0.2888 | 0.0501 | NS | 0.0534 | NS | |
| ↑ | Escherichia | 43 | 0.0048 | 0.6123 | 0.3051 | NS | 0.0510 | 0.0444 | 0.0548 | |
| Species | ↑ | coli | 45 | 0.0247 | 0.6246 | 0.2975 | 0.0414 | 0.0510 | 0.0444 | 0.0548 |
| Increased Absoulte Abundance | |
|---|---|
| Species | p-Value Basic Covariates + Probiotics + Diet |
| Unidentified/k__Fungi;p__Ascomycota;c__Leotiomycetes;o__unidentified;f__unidentified;g__unidentified;s__unidentified; | 0.0499 |
| Peziza_nivalis | 0.0260 |
| Chaetomium_erectum | 0.0176 |
| Melanophyllum_haematospermum | 0.0190 |
| Unidentified/k__Fungi;p__Ascomycota;c__Dothideomycetes;o__Capnodiales;f__Mycosphaerellaceae;g__unidentified;s__unidentified; | 0.0247 |
| Arnium_arizonense | 0.0370 |
| Unidentified/k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Microascales;f__Halosphaeriaceae;g__unidentified;s__unidentified; | 0.0246 |
| Phialophora_mustea | 0.0404 |
| Unidentified/k__Fungi;p__Ascomycota;c__Eurotiomycetes;o__Onygenales;f__unidentified;g__unidentified;s__unidentified; | 0.0266 |
| Exophiala_dermatitidis | 0.0413 |
| Dioszegia_fristingensis | 0.0413 |
| Pyrenochaeta_keratinophila | 0.0413 |
| Exophiala_oligosperma | 0.0413 |
| Alternaria_hungarica | 0.0413 |
| Peziza_buxea | 0.0413 |
| Tetragoniomyces_uliginosus | 0.0413 |
| Unidentified/k__Fungi;p__Basidiomycota;c__Agaricomycetes;o__Agaricales;f__Omphalotaceae;g__Marasmiellus;s__unidentified; | 0.0413 |
| Unidentified/k__Fungi;p__Ascomycota;c__Pezizomycetes;o__Pezizales;f__Pyronemataceae;g__Trichophaea;s__unidentified; | 0.0413 |
| Exophiala_phaeomuriformis | 0.0413 |
| Unidentified/k__Fungi;p__Ascomycota;c__Eurotiomycetes;o__Chaetothyriales;f__Herpotrichiellaceae;g__Phialophora;s__unidentified; | 0.0413 |
| Aspergillus_ochraceus | 0.0413 |
| Mortierella_zonata | 0.0413 |
| Unidentified/k__Fungi;p__Ascomycota;c__Dothideomycetes;o__Pleosporales;f__Leptosphaeriaceae;g__Leptosphaeria;s__unidentified; | 0.0413 |
| Microascus_albonigrescens | 0.0413 |
| Coniophora_olivacea | 0.0420 |
| Unidentified/k__Fungi;p__Ascomycota;c__Dothideomycetes;o__Pleosporales;f__Phaeosphaeriaceae;g__Chaetosphaeronema;s__unidentified; | 0.0379 |
| Decreased Absolute Abundance | |
| Species | p-Value Basic Covariates + Probiotics + Diet |
| Betamyces_americae-meridionalis | 0.0510 |
| Unidentified/k__Fungi;p__Chytridiomycota;c__Rhizophydiomycetes;o__Rhizophydiales;f__unidentified;g__unidentified;s__unidentified; | 0.0183 |
| Spizellomyces_dolichospermus | 0.0442 |
| Unidentified/k__Fungi;p__Ascomycota;c__Leotiomycetes;o__Helotiales;f__unidentified;g__unidentified;s__unidentified; | 0.0160 |
| Unidentified/k__Fungi;p__Basidiomycota;c__Agaricomycetes;o__Auriculariales;f__unidentified;g__unidentified;s__unidentified; | 0.0124 |
| Mortierella_amoeboidea | 0.0549 |
| Unidentified/k__Fungi;p__Mortierellomycota;c__Mortierellomycetes;o__Mortierellales;f__Mortierellaceae;g__Mortierella;s__unidentified; | 0.0140 |
| Unidentified/k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Branch06;f__unidentified;g__unidentified;s__unidentified; | 0.0428 |
| Unidentified/k__Fungi;p__Ascomycota;c__unidentified;o__unidentified;f__unidentified;g__unidentified;s__unidentified; | 0.0433 |
| Plectosphaerella_cucumerina | 0.0531 |
| DF | R-Squared | Beta | SEbeta | p Value 3 | ||||
|---|---|---|---|---|---|---|---|---|
| Phylum | Rozellomycota | k__Fungi;p__Rozellomycota; | ↓ | 21 | 0.1222 | −0.7622 | 0.3547 | 0.0434 |
| Class | Wallemiomycetes | k__Fungi;p__Basidiomycota;c__Wallemiomycetes; | ↓ | 21 | 0.3717 | −0.8819 | 0.2930 | 0.0067 |
| Spizellomycetes | k__Fungi;p__Chytridiomycota;c__Spizellomycetes; | ↓ | 21 | 0.1580 | −0.9972 | 0.3424 | 0.0083 | |
| Agaricomycetes | k__Fungi;p__Basidiomycota;c__Agaricomycetes; | ↓ | 21 | 0.3393 | −0.8713 | 0.3415 | 0.0186 | |
| Orbiliomycetes | k__Fungi;p__Ascomycota;c__Orbiliomycetes; | ↑ | 21 | 0.0222 | 0.7902 | 0.3669 | 0.0431 | |
| Order | Wallemiales | k__Fungi;p__Basidiomycota;c__Wallemiomycetes;o__Wallemiales; | ↓ | 21 | 0.3717 | −0.8819 | 0.2930 | 0.0067 |
| Spizellomycetales | k__Fungi;p__Chytridiomycota;c__Spizellomycetes;o__Spizellomycetales; | ↓ | 21 | 0.1580 | −0.9972 | 0.3424 | 0.0083 | |
| Branch06 | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Branch06; | ↓ | 21 | 0.1625 | −0.8069 | 0.3433 | 0.0286 | |
| Orbiliales | k__Fungi;p__Ascomycota;c__Orbiliomycetes;o__Orbiliales; | ↑ | 21 | −0.0004 | 0.8219 | 0.3895 | 0.0470 | |
| Family | Wallemiales_fam_Incertae_sedis | k__Fungi;p__Basidiomycota;c__Wallemiomycetes;o__Wallemiales;f__Wallemiales_fam_Incertae_sedis; | ↓ | 21 | 0.3717 | −0.8819 | 0.2930 | 0.0067 |
| Spizellomycetaceae | k__Fungi;p__Chytridiomycota;c__Spizellomycetes;o__Spizellomycetales;f__Spizellomycetaceae; | ↓ | 21 | 0.0651 | −0.9407 | 0.3653 | 0.0177 | |
| unidentified | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Branch06;f__unidentified; | ↓ | 21 | 0.1625 | −0.8069 | 0.3433 | 0.0286 | |
| Phaffomycetaceae | k__Fungi;p__Ascomycota;c__Saccharomycetes;o__Saccharomycetales;f__Phaffomycetaceae; | ↓ | 21 | 0.3319 | −0.7243 | 0.3141 | 0.0314 | |
| Leotiomycetes_fam_Incertae_sedis | k__Fungi;p__Ascomycota;c__Leotiomycetes;o__Leotiomycetes_ord_Incertae_sedis;f__Leotiomycetes_fam_Incertae_sedis; | ↓ | 21 | 0.1523 | −0.7956 | 0.3477 | 0.0326 | |
| Genus | Wallemia | k__Fungi;p__Basidiomycota;c__Wallemiomycetes;o__Wallemiales;f__Wallemiales_fam_Incertae_sedis;g__Wallemia; | ↓ | 21 | 0.3717 | −0.8819 | 0.2930 | 0.0067 |
| Saccharomyces | k__Fungi;p__Ascomycota;c__Saccharomycetes;o__Saccharomycetales;f__Saccharomycetaceae;g__Saccharomyces; | ↑ | 21 | 0.2563 | 0.9880 | 0.3354 | 0.0077 | |
| Spizellomyces | k__Fungi;p__Chytridiomycota;c__Spizellomycetes;o__Spizellomycetales;f__Spizellomycetaceae;g__Spizellomyces; | ↓ | 21 | 0.0691 | −0.9486 | 0.3713 | 0.0185 | |
| unidentified | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Branch06;f__unidentified;g__unidentified; | ↓ | 21 | 0.1625 | −0.8069 | 0.3433 | 0.0286 | |
| Cyberlindnera | k__Fungi;p__Ascomycota;c__Saccharomycetes;o__Saccharomycetales;f__Phaffomycetaceae;g__Cyberlindnera; | ↓ | 21 | 0.2069 | −0.8408 | 0.3647 | 0.0314 | |
| Scytalidium | k__Fungi;p__Ascomycota;c__Leotiomycetes;o__Leotiomycetes_ord_Incertae_sedis;f__Leotiomycetes_fam_Incertae_sedis;g__Scytalidium; | ↓ | 21 | 0.1523 | −0.7956 | 0.3477 | 0.0326 | |
| Plectosphaerella | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Glomerellales;f__Plectosphaerellaceae;g__Plectosphaerella; | ↓ | 21 | 0.1150 | −0.5678 | 0.2598 | 0.0403 | |
| unidentified | k__Fungi;p__Ascomycota;c__Dothideomycetes;o__Dothideomycetidae_ord_Incertae_sedis;f__Eremomycetaceae;g__unidentified; | ↑ | 21 | 0.0949 | 0.7849 | 0.3637 | 0.0426 | |
| Species | unidentified | k__Fungi;p__Basidiomycota;c__Wallemiomycetes;o__Wallemiales;f__Wallemiales_fam_Incertae_sedis;g__Wallemia;s__unidentified; | ↓ | 21 | 0.4107 | −0.9420 | 0.2880 | 0.0036 |
| Saccharomyces_cerevisiae | k__Fungi;p__Ascomycota;c__Saccharomycetes;o__Saccharomycetales;f__Saccharomycetaceae;g__Saccharomyces;s__Saccharomyces_cerevisiae; | ↑ | 21 | 0.2563 | 0.9880 | 0.3354 | 0.0077 | |
| Spizellomyces_lactosolyticus | k__Fungi;p__Chytridiomycota;c__Spizellomycetes;o__Spizellomycetales;f__Spizellomycetaceae;g__Spizellomyces;s__Spizellomyces_lactosolyticus; | ↓ | 21 | 0.0336 | −0.9039 | 0.3694 | 0.0233 | |
| unidentified | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Branch06;f__unidentified;g__unidentified;s__unidentified; | ↓ | 21 | 0.1625 | −0.8069 | 0.3433 | 0.0286 | |
| Oidiodendron_tenuissimum | k__Fungi;p__Ascomycota;c__Leotiomycetes;o__Helotiales;f__Myxotrichaceae;g__Oidiodendron;s__Oidiodendron_tenuissimum; | ↓ | 21 | 0.2239 | −0.8286 | 0.3600 | 0.0317 | |
| Scytalidium_circinatum | k__Fungi;p__Ascomycota;c__Leotiomycetes;o__Leotiomycetes_ord_Incertae_sedis;f__Leotiomycetes_fam_Incertae_sedis;g__Scytalidium;s__Scytalidium_circinatum; | ↓ | 21 | 0.1523 | −0.7956 | 0.3477 | 0.0326 | |
| Cyberlindnera_fabianii | k__Fungi;p__Ascomycota;c__Saccharomycetes;o__Saccharomycetales;f__Phaffomycetaceae;g__Cyberlindnera;s__Cyberlindnera_fabianii; | ↓ | 21 | 0.1991 | −0.8316 | 0.3662 | 0.0338 | |
| Plectosphaerella_cucumerina | k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Glomerellales;f__Plectosphaerellaceae;g__Plectosphaerella;s__Plectosphaerella_cucumerina; | ↓ | 21 | 0.1150 | −0.5678 | 0.2598 | 0.0403 | |
| unidentified | k__Fungi;p__Ascomycota;c__Dothideomycetes;o__Dothideomycetidae_ord_Incertae_sedis;f__Eremomycetaceae;g__unidentified;s__unidentified; | ↑ | 21 | 0.0949 | 0.7849 | 0.3637 | 0.0426 | |
| Calicium_viride | k__Fungi;p__Ascomycota;c__Lecanoromycetes;o__Caliciales;f__Caliciaceae;g__Calicium;s__Calicium_viride; | ↓ | 21 | 0.2079 | −0.5845 | 0.2745 | 0.0452 |
| Increased in GID vs. CTRL | Decreased in GID vs. CTRL | ||||
|---|---|---|---|---|---|
| Species | R-squared | beta | SEbeta | p-value basic covariates + probiotics + diet | no species significantly decreased in GID compared to CTRL |
| Saccharomyces_cerevisiae | 0.27665113 | 1.09894673 | 0.34372426 | 0.01138158 | |
| Candida_parapsilosis | 0.28455775 | 1.05552026 | 0.37516274 | 0.01592194 | |
| Metarhizium_anisopliae | 0.2610366 | 0.14201073 | 0.05109858 | 0.02205624 | |
| unidentified | 0.21517888 | 0.9223066 | 0.37481416 | 0.0260362 | |
| Humicola_phialophoroides | 0.10755173 | 0.81109936 | 0.35117492 | 0.04047386 | |
| Penicillium_pimiteouiense | 0.17719919 | 0.85081527 | 0.36915659 | 0.03106863 | |
| Fusarium_solani | 0.17912126 | 0.90662454 | 0.41162482 | 0.04566077 | |
| unidentified | 0.46101634 | −0.9334353 | 0.31142748 | 0.00943343 | |
| Phaeoacremonium_hungaricum | 0.19271259 | 0.48732341 | 0.22192725 | 0.04648654 | |
| Calicium_viride | 0.16255788 | −0.6647188 | 0.31299161 | 0.04865476 | |
| T-TEST ABX VS. NON-ABX_BACTERIA | ||||||||
|---|---|---|---|---|---|---|---|---|
| Name | Statistic | df | p-Value | CI | CI | Mean of +abx | Mean of −abx | |
| Phylum | TM7 | 2.0450 | 26 | 0.0509 | −0.0018 | 0.8286 | 0.5465 | 0.1331 |
| Class | TM7-3 | 2.0450 | 26 | 0.0509 | −0.0018 | 0.8286 | 0.5465 | 0.1331 |
| Family | Succinivibrionaceae | −2.3054 | 30 | 0.0281 | −0.0713 | −0.0043 | −0.0512 | −0.0134 |
| Genus | Butyricimonas | −3.3256 | 38 | 0.0020 | −0.8139 | −0.1979 | −0.2551 | 0.2508 |
| Dorea | −2.4678 | 49 | 0.0172 | −0.3432 | −0.0351 | −0.2667 | −0.0776 | |
| Eggerthella | 2.4743 | 21 | 0.0219 | 0.0794 | 0.9150 | 0.3092 | −0.1880 | |
| Succinivibrio | −2.3054 | 30 | 0.0281 | −0.0713 | −0.0043 | −0.0512 | −0.0134 | |
| Species | eggerthii | −2.3689 | 49 | 0.0218 | −0.1606 | −0.0132 | −0.0693 | 0.0176 |
| lenta | 2.4743 | 21 | 0.0219 | 0.0794 | 0.9150 | 0.3092 | −0.1880 | |
| T-TEST ABX VS. NON-ABX_FUNGI | ||||||||
| Name | Statistic | df | p-Value | CI | CI | Mean of +abx | Mean of −abx | |
| Phylum | Neocallimastigomycota | −2.4086 | 27 | 0.0231 | −0.2759 | −0.0221 | −0.1031 | 0.0459 |
| Class | Ascomycota_cls_Incertae_sedis | −2.5506 | 28 | 0.0165 | −0.7215 | −0.0787 | −0.0487 | 0.3514 |
| Neocallimastigomycetes | −2.4086 | 27 | 0.0231 | −0.2759 | −0.0221 | −0.1031 | 0.0459 | |
| Order | Ascomycota_ord_Incertae_sedis | −2.5506 | 28 | 0.0165 | −0.7215 | −0.0787 | −0.0487 | 0.3514 |
| Neocallimastigales | −2.4086 | 27 | 0.0231 | −0.2759 | −0.0221 | −0.1031 | 0.0459 | |
| Coniochaetales | −2.3188 | 25 | 0.0287 | −0.4096 | −0.0244 | −0.0277 | 0.1893 | |
| Caliciales | −2.0308 | 26 | 0.0526 | −0.4670 | 0.0029 | 0.0024 | 0.2345 | |
| Family | Ascomycota_fam_Incertae_sedis | −2.5506 | 28 | 0.0165 | −0.7215 | −0.0787 | −0.0487 | 0.3514 |
| unidentified/k__Fungi;p__Mortierellomycota;c__Mortierellomycetes;o__Mortierellales;f__unidentified; | 2.7003 | 13 | 0.0184 | 0.1502 | 1.3624 | 0.6390 | −0.1173 | |
| unidentified/k__Fungi;p__Ascomycota;c__Dothideomycetes;o__Pleosporales;f__unidentified; | −2.4067 | 29 | 0.0226 | −1.0320 | −0.0841 | −0.1434 | 0.4147 | |
| Neocallimastigaceae | −2.4086 | 27 | 0.0231 | −0.2759 | −0.0221 | −0.1031 | 0.0459 | |
| Cortinariaceae | −2.2683 | 24 | 0.0326 | −0.3772 | −0.0178 | −0.0234 | 0.1741 | |
| Caliciaceae | −2.0308 | 26 | 0.0526 | −0.4670 | 0.0029 | 0.0024 | 0.2345 | |
| Genus | unidentified/k__Fungi;p__Mortierellomycota;c__Mortierellomycetes;o__Mortierellales;f__unidentified;g__unidentified; | 2.7003 | 13 | 0.0184 | 0.1502 | 1.3624 | 0.6390 | −0.1173 |
| unidentified/k__Fungi;p__Ascomycota;c__Dothideomycetes;o__Pleosporales;f__unidentified;g__unidentified; | −2.4067 | 29 | 0.0226 | −1.0320 | −0.0841 | −0.1434 | 0.4147 | |
| unidentified/k__Fungi;p__Neocallimastigomycota;c__Neocallimastigomycetes;o__Neocallimastigales;f__Neocallimastigaceae;g__unidentified; | −2.4086 | 27 | 0.0231 | −0.2759 | −0.0221 | −0.1031 | 0.0459 | |
| unidentified/k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Glomerellales;f__Plectosphaerellaceae;g__unidentified; | 2.6718 | 9 | 0.0254 | 0.1292 | 1.5437 | 0.8394 | 0.0029 | |
| unidentified/k__Fungi;p__Ascomycota;c__Pezizomycetes;o__Pezizales;f__Pyronemataceae;g__unidentified; | −2.2488 | 28 | 0.0325 | −0.4571 | −0.0215 | −0.0731 | 0.1662 | |
| Calicium | −2.1900 | 28 | 0.0369 | −0.5191 | −0.0175 | −0.0296 | 0.2387 | |
| Neurospora | −2.1516 | 21 | 0.0432 | −0.6826 | −0.0116 | 0.0000 | 0.3471 | |
| unidentified/k__Fungi;p__Ascomycota;c__Dothideomycetes;o__Pleosporales;f__Pleosporaceae;g__unidentified; | −2.1190 | 22 | 0.0456 | −0.3204 | −0.0035 | −0.0124 | 0.1495 | |
| Didymella | −2.0777 | 28 | 0.0469 | −0.7601 | −0.0056 | −0.0625 | 0.3203 | |
| unidentified/k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Sordariales;f__Sordariaceae;g__unidentified; | −2.0729 | 21 | 0.0507 | −0.6871 | 0.0011 | 0.0000 | 0.3430 | |
| Specie | unidentified/k__Fungi;p__Mortierellomycota;c__Mortierellomycetes;o__Mortierellales;f__unidentified;g__unidentified;s__unidentified; | 2.7003 | 13 | 0.0184 | 0.1502 | 1.3624 | 0.6390 | −0.1173 |
| unidentified/k__Fungi;p__Ascomycota;c__Dothideomycetes;o__Pleosporales;f__unidentified;g__unidentified;s__unidentified; | −2.4067 | 29 | 0.0226 | −1.0320 | −0.0841 | −0.1434 | 0.4147 | |
| unidentified/k__Fungi;p__Neocallimastigomycota;c__Neocallimastigomycetes;o__Neocallimastigales;f__Neocallimastigaceae;g__unidentified;s__unidentified; | −2.4086 | 27 | 0.0231 | −0.2759 | −0.0221 | −0.1031 | 0.0459 | |
| unidentified/k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Glomerellales;f__Plectosphaerellaceae;g__unidentified;s__unidentified; | 2.6718 | 9 | 0.0254 | 0.1292 | 1.5437 | 0.8394 | 0.0029 | |
| unidentified/k__Fungi;p__Ascomycota;c__Pezizomycetes;o__Pezizales;f__Pyronemataceae;g__unidentified;s__unidentified; | −2.2488 | 28 | 0.0325 | −0.4571 | −0.0215 | −0.0731 | 0.1662 | |
| Oidiodendron_truncatum | −2.1978 | 20 | 0.0399 | −0.6164 | −0.0161 | 0.0007 | 0.3170 | |
| Neurospora_terricola | −2.1516 | 21 | 0.0432 | −0.6826 | −0.0116 | 0.0000 | 0.3471 | |
| Rhodotorula_mucilaginosa | −2.1079 | 27 | 0.0444 | −0.3907 | −0.0053 | −0.0745 | 0.1235 | |
| unidentified/k__Fungi;p__Ascomycota;c__Dothideomycetes;o__Pleosporales;f__Pleosporaceae;g__unidentified;s__unidentified; | −2.1190 | 22 | 0.0456 | −0.3204 | −0.0035 | −0.0124 | 0.1495 | |
| Didymella_calidophila | −2.0777 | 28 | 0.0469 | −0.7601 | −0.0056 | −0.0625 | 0.3203 | |
| unidentified/k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Sordariales;f__Sordariaceae;g__unidentified;s__unidentified; | −2.0729 | 21 | 0.0507 | −0.6871 | 0.0011 | 0.0000 | 0.3430 | |
| unidentified/k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Hypocreales;f__Cordycipitaceae;g__Lecanicillium;s__unidentified; | −2.0597 | 21 | 0.0520 | −0.2603 | 0.0013 | 0.0000 | 0.1295 | |
| BACTERIA_NDD_ABX | ||||||
|---|---|---|---|---|---|---|
| NAME | DF | R-Squared | Beta | SEbeta | p-Value | |
| Family | Campylobacteraceae | 29 | 0.1678 | −0.1755 | 0.0740 | 0.0246 |
| Corynebacteriaceae | 28 | 0.1823 | −0.1587 | 0.0735 | 0.0395 | |
| Genus | Rothia | 28 | 0.2196 | 0.3948 | 0.1658 | 0.0243 |
| Campylobacter | 29 | 0.1678 | −0.1755 | 0.0740 | 0.0246 | |
| WAL_1855D | 29 | 0.1869 | −0.4043 | 0.1749 | 0.0281 | |
| Mogibacterium | 29 | 0.2957 | 0.3538 | 0.1532 | 0.0283 | |
| Butyricimonas | 28 | 0.1855 | 0.6542 | 0.2915 | 0.0329 | |
| Adlercreutzia | 29 | 0.1526 | 0.5718 | 0.2629 | 0.0380 | |
| Corynebacterium | 28 | 0.1823 | −0.1587 | 0.0735 | 0.0395 | |
| Lactococcus | 30 | 0.2294 | 0.0998 | 0.0475 | 0.0440 | |
| Species | zeae | 29 | 0.2868 | 0.0120 | 0.0043 | 0.0092 |
| eggerthii | 30 | 0.0677 | 0.1520 | 0.0625 | 0.0213 | |
| FUNGI_NDD_ABX | ||||||
| DF | R-Squared | Beta | SEbeta | p-Value | ||
| Phylum | Olpidiomycota | 12 | 0.2202 | −1.2000 | 0.4241 | 0.0152 |
| Class | Olpidiomycetes | 12 | 0.2225 | −1.2098 | 0.4265 | 0.0150 |
| Order | Leotiomycetes_ord_Incertae_sedis | 12 | 0.3393 | −1.4495 | 0.4912 | 0.0121 |
| Olpidiales | 12 | 0.2225 | −1.2098 | 0.4265 | 0.0150 | |
| Chaetothyriales | 12 | 0.2304 | −0.3137 | 0.1328 | 0.0359 | |
| Sebacinales | 12 | 0.0227 | −0.6656 | 0.3112 | 0.0537 | |
| Family | Bolbitiaceae | 12 | 0.1570 | −0.5405 | 0.1897 | 0.0146 |
| Unidentified/k__Fungi;p__Olpidiomycota;c__Olpidiomycetes;o__Olpidiales;f__unidentified; | 12 | 0.1817 | −0.9807 | 0.3936 | 0.0283 | |
| Bulleraceae | 13 | 0.3745 | 0.6050 | 0.2638 | 0.0391 | |
| Cyphellophoraceae | 12 | 0.4351 | 0.0220 | 0.0100 | 0.0485 | |
| Genus | Spizellomyces | 11 | 0.6659 | −0.8910 | 0.2156 | 0.0017 |
| Wickerhamomyces | 13 | 0.5155 | −1.2559 | 0.3417 | 0.0028 | |
| Mycothermus | 12 | 0.4477 | −0.6108 | 0.1895 | 0.0073 | |
| Unidentified/k__Fungi;p__Olpidiomycota;c__Olpidiomycetes;o__Olpidiales;f__unidentified;g__unidentified; | 12 | 0.1817 | −0.9807 | 0.3936 | 0.0283 | |
| Chaetosphaeria | 12 | 0.4210 | 0.4878 | 0.2078 | 0.0369 | |
| Unidentified/ k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Glomerellales;f__Plectosphaerellaceae;g__unidentified; | 12 | 0.3705 | −0.5830 | 0.2507 | 0.0384 | |
| Bullera | 13 | 0.3745 | 0.6050 | 0.2638 | 0.0391 | |
| Rhodotorula | 12 | 0.2511 | 0.3827 | 0.1657 | 0.0395 | |
| Torulaspora | 13 | 0.4224 | 0.5985 | 0.2622 | 0.0399 | |
| Cyphellophora | 12 | 0.4351 | 0.0220 | 0.0100 | 0.0485 | |
| Cercosporella | 13 | 0.0391 | −0.4085 | 0.1930 | 0.0541 | |
| Specie | Mycothermus_thermophilus | 12 | 0.4477 | −0.6108 | 0.1895 | 0.0073 |
| Unidentified/k__Fungi;p__Olpidiomycota;c__Olpidiomycetes;o__Olpidiales;f__unidentified;g__unidentified;s__unidentified; | 12 | 0.1817 | −0.9807 | 0.3936 | 0.0283 | |
| Pichia_membranifaciens | 12 | 0.3877 | 0.3491 | 0.1427 | 0.0309 | |
| Chaetosphaeria_chloroconia | 12 | 0.4224 | 0.4856 | 0.2079 | 0.0376 | |
| Penicillium_pimiteouiense | 13 | 0.2521 | −0.5084 | 0.2202 | 0.0380 | |
| Unidentified/ k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Glomerellales;f__Plectosphaerellaceae;g__unidentified;s__unidentified; | 12 | 0.3705 | −0.5830 | 0.2507 | 0.0384 | |
| Bullera_unica | 13 | 0.3744 | 0.6056 | 0.2640 | 0.0391 | |
| Torulaspora_delbrueckii | 13 | 0.4223 | 0.5982 | 0.2622 | 0.0400 | |
| Cutaneotrichosporon_cyanovorans | 12 | 0.4212 | 0.6892 | 0.3091 | 0.0457 | |
| Candida_glaebosa | 12 | 0.4241 | −0.0525 | 0.0236 | 0.0461 | |
| Malassezia_sympodialis | 12 | 0.0533 | −0.1744 | 0.0788 | 0.0471 | |
| Unidentified/k__Fungi;p__Basidiomycota;c__Agaricomycetes;o__Agaricales;f__Amanitaceae;g__Amanita;s__unidentified; | 13 | 0.1741 | 0.0428 | 0.0200 | 0.0519 | |
| Cercosporella_tinosporae | 13 | 0.0391 | −0.4085 | 0.1930 | 0.0541 | |
| BACTERIA_GID_ABX | ||||||
|---|---|---|---|---|---|---|
| DF | R-Squared | Beta | SEbeta | p-Value | ||
| Genus | Atopobium | 28 | 0.0880 | 0.3233 | 0.1487 | 0.0383 |
| Allobaculum | 29 | 0.0745 | −0.0892 | 0.0436 | 0.0502 | |
| species | eggerthii | 30 | 0.0494 | 0.1431 | 0.0625 | 0.0292 |
| FUNGI_GID_ABX | ||||||
| NAME | DF | R-Squared | Beta | SEbeta | p-Value | |
| Phylum | Olpidiomycota | 17 | 0.1176 | −0.9239 | 0.3416 | 0.0150 |
| Neocallimastigomycota | 17 | 0.2300 | 0.2260 | 0.0970 | 0.0325 | |
| Class | Olpidiomycetes | 17 | 0.2028 | −0.9728 | 0.3292 | 0.0089 |
| Neocallimastigomycetes | 17 | 0.2300 | 0.2260 | 0.0970 | 0.0325 | |
| Ascomycota_cls_Incertae_sedis | 17 | 0.1328 | 0.6103 | 0.2704 | 0.0375 | |
| Order | Olpidiales | 17 | 0.2028 | −0.9728 | 0.3292 | 0.0089 |
| Neocallimastigales | 17 | 0.2300 | 0.2260 | 0.0970 | 0.0325 | |
| Ascomycota_ord_Incertae_sedis | 17 | 0.1328 | 0.6103 | 0.2704 | 0.0375 | |
| Family | Herpotrichiellaceae | 18 | 0.2303 | −0.1517 | 0.0522 | 0.0094 |
| Unidentified/k__Fungi;p__Olpidiomycota;c__Olpidiomycetes;o__Olpidiales;f__unidentified; | 17 | 0.1568 | −0.8076 | 0.3105 | 0.0186 | |
| Debaryomycetaceae | 17 | 0.2249 | −0.7173 | 0.2959 | 0.0268 | |
| Neocallimastigaceae | 17 | 0.2300 | 0.2260 | 0.0970 | 0.0325 | |
| Ascomycota_fam_Incertae_sedis | 17 | 0.1328 | 0.6103 | 0.2704 | 0.0375 | |
| Piskurozymaceae | 17 | 0.0240 | 0.0370 | 0.0178 | 0.0534 | |
| Genus | Spizellomyces | 16 | 0.4395 | −0.6295 | 0.1916 | 0.0047 |
| Mycothermus | 17 | 0.3457 | −0.6010 | 0.1907 | 0.0058 | |
| Wickerhamomyces | 17 | 0.3491 | −0.9463 | 0.3095 | 0.0071 | |
| Neonectria | 18 | 0.1834 | −0.6190 | 0.2156 | 0.0102 | |
| Unidentified/k__Fungi;p__Olpidiomycota;c__Olpidiomycetes;o__Olpidiales;f__unidentified;g__unidentified; | 17 | 0.1568 | −0.8076 | 0.3105 | 0.0186 | |
| Vishniacozyma | 17 | 0.1886 | −0.6668 | 0.2586 | 0.0195 | |
| Unidentified/k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Glomerellales;f__Plectosphaerellaceae;g__unidentified; | 17 | 0.2170 | −0.7604 | 0.3030 | 0.0225 | |
| Debaryomyces | 17 | 0.2666 | −0.6988 | 0.2822 | 0.0241 | |
| Penicillium | 16 | 0.1926 | −0.2792 | 0.1122 | 0.0242 | |
| Unidentified/k__Fungi;p__Neocallimastigomycota;c__Neocallimastigomycetes;o__Neocallimastigales;f__Neocallimastigaceae;g__unidentified; | 17 | 0.2300 | 0.2260 | 0.0970 | 0.0325 | |
| Specie | Mycothermus_thermophilus | 17 | 0.3457 | −0.6010 | 0.1907 | 0.0058 |
| Neonectria_candida | 18 | 0.1834 | −0.6190 | 0.2156 | 0.0102 | |
| Pichia_kluyveri | 16 | 0.3211 | −0.4299 | 0.1481 | 0.0104 | |
| Oidiodendron_truncatum | 17 | 0.3488 | 0.6676 | 0.2444 | 0.0142 | |
| Unidentified/k__Fungi;p__Olpidiomycota;c__Olpidiomycetes;o__Olpidiales;f__unidentified;g__unidentified;s__unidentified; | 17 | 0.1568 | −0.8076 | 0.3105 | 0.0186 | |
| Vishniacozyma_victoriae | 17 | 0.1889 | −0.6696 | 0.2599 | 0.0196 | |
| Penicillium_pimiteouiense | 17 | 0.3329 | −0.4693 | 0.1855 | 0.0216 | |
| Unidentified/k__Fungi;p__Ascomycota;c__Sordariomycetes;o__Glomerellales;f__Plectosphaerellaceae;g__unidentified;s__unidentified; | 17 | 0.2170 | −0.7604 | 0.3030 | 0.0225 | |
| Debaryomyces_udenii | 17 | 0.2666 | −0.6988 | 0.2822 | 0.0241 | |
| Unidentified/k__Fungi;p__Neocallimastigomycota;c__Neocallimastigomycetes;o__Neocallimastigales;f__Neocallimastigaceae;g__unidentified;s__unidentified; | 17 | 0.2300 | 0.2260 | 0.0970 | 0.0325 | |
| Malassezia_sympodialis | 17 | 0.0633 | −0.1761 | 0.0777 | 0.0368 | |
| Unidentified/k__Fungi;p__Basidiomycota;c__Agaricomycetes;o__Agaricales;f__Amanitaceae;g__Amanita;s__unidentified; | 17 | 0.2135 | 0.0476 | 0.0213 | 0.0396 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laswi, I.; Shafiq, A.; Al-Ali, D.; Burney, Z.; Pillai, K.; Salameh, M.; Mhaimeed, N.; Zakaria, D.; Chaari, A.; Yousri, N.A.; et al. A Comparative Pilot Study of Bacterial and Fungal Dysbiosis in Neurodevelopmental Disorders and Gastrointestinal Disorders: Commonalities, Specificities and Correlations with Lifestyle. Microorganisms 2021, 9, 741. https://doi.org/10.3390/microorganisms9040741
Laswi I, Shafiq A, Al-Ali D, Burney Z, Pillai K, Salameh M, Mhaimeed N, Zakaria D, Chaari A, Yousri NA, et al. A Comparative Pilot Study of Bacterial and Fungal Dysbiosis in Neurodevelopmental Disorders and Gastrointestinal Disorders: Commonalities, Specificities and Correlations with Lifestyle. Microorganisms. 2021; 9(4):741. https://doi.org/10.3390/microorganisms9040741
Chicago/Turabian StyleLaswi, Ibrahim, Ameena Shafiq, Dana Al-Ali, Zain Burney, Krishnadev Pillai, Mohammad Salameh, Nada Mhaimeed, Dalia Zakaria, Ali Chaari, Noha A. Yousri, and et al. 2021. "A Comparative Pilot Study of Bacterial and Fungal Dysbiosis in Neurodevelopmental Disorders and Gastrointestinal Disorders: Commonalities, Specificities and Correlations with Lifestyle" Microorganisms 9, no. 4: 741. https://doi.org/10.3390/microorganisms9040741
APA StyleLaswi, I., Shafiq, A., Al-Ali, D., Burney, Z., Pillai, K., Salameh, M., Mhaimeed, N., Zakaria, D., Chaari, A., Yousri, N. A., & Bendriss, G. (2021). A Comparative Pilot Study of Bacterial and Fungal Dysbiosis in Neurodevelopmental Disorders and Gastrointestinal Disorders: Commonalities, Specificities and Correlations with Lifestyle. Microorganisms, 9(4), 741. https://doi.org/10.3390/microorganisms9040741