
BipD of Burkholderia pseudomallei: Structure, Functions, and Detection Methods

 ,
,  ,
,
Abstract
1. Introduction
2. Content
2.1. Type III Secretion Systems (T3SSs) of B. pseudomallei
2.2. BipD of Bsa
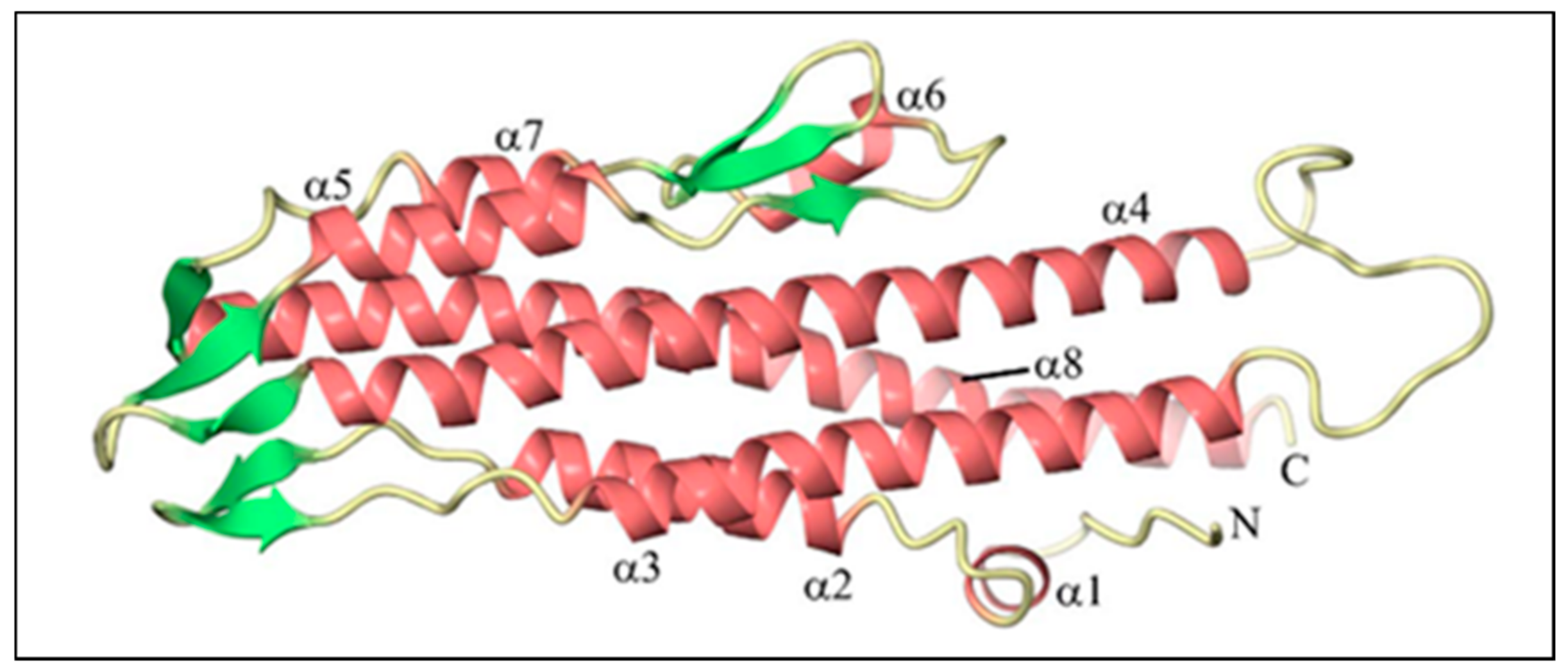
2.3. The Three-Dimensional (3D) Structure of BipD
2.4. Functions of BipD in the Pathogenesis of B. pseudomallei
2.5. Detection of BipD-Specific Antibodies for Diagnosis
Serological Tests
3. Conclusions
4. Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sprague, L.; Neubauer, H. Melioidosis in animals: A review on epizootiology, diagnosis and clinical presentation. J. Vet. Med. Ser. B 2004, 51, 305–320. [Google Scholar] [CrossRef] [PubMed]
- Limmathurotsakul, D.; Wongratanacheewin, S.; Teerawattanasook, N.; Wongsuvan, G.; Chaisuksant, S.; Chetchotisakd, P.; Chaowagul, W.; Day, N.P.; Peacock, S.J. Increasing incidence of human melioidosis in Northeast Thailand. Am. J. Trop. Med. Hyg. 2010, 82, 1113–1117. [Google Scholar] [CrossRef]
- Stewart, T.; Engelthaler, D.M.; Blaney, D.D.; Tuanyok, A.; Wangsness, E.; Smith, T.L.; Pearson, T.; Komatsu, K.K.; Keim, P.; Currie, B.J. Epidemiology and investigation of melioidosis, Southern Arizona. Emerg. Infect. Dis. 2011, 17, 1286. [Google Scholar] [CrossRef] [PubMed]
- Le Tohic, S.; Montana, M.; Koch, L.; Curti, C.; Vanelle, P. A review of melioidosis cases imported into Europe. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 1395–1408. [Google Scholar] [CrossRef] [PubMed]
- Limmathurotsakul, D.; Golding, N.; Dance, D.A.; Messina, J.P.; Pigott, D.M.; Moyes, C.L.; Rolim, D.B.; Bertherat, E.; Day, N.P.; Peacock, S.J. Predicted global distribution of Burkholderia pseudomallei and burden of melioidosis. Nat. Microbiol. 2016, 1, 15008. [Google Scholar] [CrossRef]
- Nathan, S.; Chieng, S.; Kingsley, P.V.; Mohan, A.; Podin, Y.; Ooi, M.-H.; Mariappan, V.; Vellasamy, K.M.; Vadivelu, J.; Daim, S. Melioidosis in Malaysia: Incidence, clinical challenges, and advances in understanding pathogenesis. Trop. Med. Infect. Dis. 2018, 3, 25. [Google Scholar] [CrossRef]
- Wiersinga, W.J.; Van der Poll, T.; White, N.J.; Day, N.P.; Peacock, S.J. Melioidosis: Insights into the pathogenicity of Burkholderia pseudomallei. Nat. Rev. Microbiol. 2006, 4, 272–282. [Google Scholar] [CrossRef] [PubMed]
- Wiersinga, W.J.; Virk, H.S.; Torres, A.G.; Currie, B.J.; Peacock, S.J.; Dance, D.A.B.; Limmathurotsakul, D. Melioidosis. Nat. Rev. Dis. Primers 2018, 4, 17107. [Google Scholar] [CrossRef] [PubMed]
- Limmathurotsakul, D.; Kanoksil, M.; Wuthiekanun, V.; Kitphati, R.; deStavola, B.; Day, N.P.; Peacock, S.J. Activities of daily living associated with acquisition of melioidosis in northeast Thailand: A matched case-control study. PLoS Negl. Trop. Dis. 2013, 7, e2072. [Google Scholar] [CrossRef]
- Cheng, A.C.; Currie, B.J. Melioidosis: Epidemiology, pathophysiology, and management. Clin. Microbiol. Rev. 2005, 18, 383–416. [Google Scholar] [CrossRef] [PubMed]
- Choy, J.L.; Mayo, M.; Janmaat, A.; Currie, B.J. Animal melioidosis in Australia. Acta Trop. 2000, 74, 153–158. [Google Scholar] [CrossRef]
- Chandni, R. Melioidosis: The great mimicker. Med. Update 2013, 23, 14–18. [Google Scholar]
- Churuangsuk, C.; Chusri, S.; Hortiwakul, T.; Charernmak, B.; Silpapojakul, K. Characteristics, clinical outcomes and factors influencing mortality of patients with melioidosis in southern Thailand: A 10-year retrospective study. Asian Pac. J. Trop. Med. 2016, 9, 256–260. [Google Scholar] [CrossRef]
- Currie, B.J.; Ward, L.; Cheng, A.C. The epidemiology and clinical spectrum of melioidosis: 540 cases from the 20 year Darwin prospective study. PLoS Negl. Trop. Dis. 2010, 4, e900. [Google Scholar] [CrossRef]
- Currie, B.J.; Jacups, S.P.; Cheng, A.C.; Fisher, D.A.; Anstey, N.M.; Huffam, S.E.; Krause, V.L. Melioidosis epidemiology and risk factors from a prospective whole-population study in northern Australia. Trop. Med. Int. Health 2004, 9, 1167–1174. [Google Scholar] [CrossRef] [PubMed]
- Moreno, R.M.; Jimenez, V.; Monroy, F.P. Impact of Binge Alcohol Intoxication on the Humoral Immune Response during Burkholderia spp. Infections. Microorganisms 2019, 7, 125. [Google Scholar] [CrossRef]
- Ngauy, V.; Lemeshev, Y.; Sadkowski, L.; Crawford, G. Cutaneous melioidosis in a man who was taken as a prisoner of war by the Japanese during World War II. J. Clin. Microbiol. 2005, 43, 970–972. [Google Scholar] [CrossRef] [PubMed]
- Russo, R.; Chae, R.; Mukherjee, S.; Singleton, E.J.; Occi, J.L.; Kadouri, D.E.; Connell, N.D. Susceptibility of select agents to predation by predatory bacteria. Microorganisms 2015, 3, 903–912. [Google Scholar] [CrossRef]
- Chetchotisakd, P.; Chierakul, W.; Chaowagul, W.; Anunnatsiri, S.; Phimda, K.; Mootsikapun, P.; Chaisuksant, S.; Pilaikul, J.; Thinkhamrop, B.; Phiphitaporn, S. Trimethoprim-sulfamethoxazole versus trimethoprim-sulfamethoxazole plus doxycycline as oral eradicative treatment for melioidosis (MERTH): A multicentre, double-blind, non-inferiority, randomised controlled trial. Lancet 2014, 383, 807–814. [Google Scholar] [CrossRef]
- Ahmad, N.; Hashim, R.; Mohd Noor, A. The in vitro antibiotic susceptibility of Malaysian isolates of Burkholderia pseudomallei. Int. J. Microbiol. 2013, 2013. [Google Scholar] [CrossRef]
- Limmathurotsakul, D.; Funnell, S.G.; Torres, A.G.; Morici, L.A.; Brett, P.J.; Dunachie, S.; Atkins, T.; Altmann, D.M.; Bancroft, G.; Peacock, S.J. Consensus on the development of vaccines against naturally acquired melioidosis. Emerg. Infect. Dis. 2015, 21, e141480. [Google Scholar] [CrossRef] [PubMed]
- Tellapragada, C.; Shaw, T.; D’Souza, A.; Eshwara, V.K.; Mukhopadhyay, C. Improved detection of Burkholderia pseudomallei from non-blood clinical specimens using enrichment culture and PCR: Narrowing diagnostic gap in resource-constrained settings. Trop. Med. Int. Health 2017, 22, 866–870. [Google Scholar] [CrossRef] [PubMed]
- Premaratne, K.; Karunaratne, G.; Dias, R.; Lamahewage, A.; Samarasinghe, M.; Corea, E.; Gunawardena, R. Melioidosis presenting as parotid abscess in children: Two consecutive cases. Sri Lankan J. Infect. Dis. 2017, 7, 116–122. [Google Scholar] [CrossRef]
- Chen, P.; Gates-Hollingsworth, M.; Pandit, S.; Park, A.; Montgomery, D.; AuCoin, D.; Gu, J.; Zenhausern, F. based Vertical Flow Immunoassay (VFI) for detection of bio-threat pathogens. Talanta 2019, 191, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Woods, K.L.; Boutthasavong, L.; NicFhogartaigh, C.; Lee, S.J.; Davong, V.; AuCoin, D.P.; Dance, D.A. Evaluation of a rapid diagnostic test for detection of Burkholderia pseudomallei in the Lao People’s Democratic Republic. J. Clin. Microbiol. 2018, 56. [Google Scholar] [CrossRef]
- Pumpuang, A.; Dunachie, S.J.; Phokrai, P.; Jenjaroen, K.; Sintiprungrat, K.; Boonsilp, S.; Brett, P.J.; Burtnick, M.N.; Chantratita, N. Comparison of O-polysaccharide and hemolysin co-regulated protein as target antigens for serodiagnosis of melioidosis. PLoS Negl. Trop. Dis. 2017, 11, e0005499. [Google Scholar] [CrossRef] [PubMed]
- Chaichana, P.; Jenjaroen, K.; Amornchai, P.; Chumseng, S.; Langla, S.; Rongkard, P.; Sumonwiriya, M.; Jeeyapant, A.; Chantratita, N.; Teparrukkul, P. Antibodies in melioidosis: The role of the indirect hemagglutination assay in evaluating patients and exposed populations. Am. J. Trop. Med. Hyg. 2018, 99, 1378–1385. [Google Scholar] [CrossRef] [PubMed]
- Kohler, C.; Dunachie, S.J.; Müller, E.; Kohler, A.; Jenjaroen, K.; Teparrukkul, P.; Baier, V.; Ehricht, R.; Steinmetz, I. Rapid and sensitive multiplex detection of Burkholderia pseudomallei-specific antibodies in melioidosis patients based on a protein microarray approach. PLoS Negl. Trop. Dis. 2016, 10, e0004847. [Google Scholar] [CrossRef]
- Dulsuk, A.; Paksanont, S.; Sangchankoom, A.; Ekchariyawat, P.; Phunpang, R.; Jutrakul, Y.; Chantratita, N.; West, T.E. Validation of a monoclonal antibody-based immunofluorescent assay to detect Burkholderia pseudomallei in blood cultures. Trans. R. Soc. Trop. Med. Hyg. 2016, 110, 670–672. [Google Scholar] [CrossRef]
- Peeters, M.; Chung, P.; Lin, H.; Mortelmans, K.; Phe, C.; San, C.; Kuijpers, L.M.F.; Teav, S.; Phe, T.; Jacobs, J. Diagnostic accuracy of the InBiOS AMD rapid diagnostic test for the detection of Burkholderia pseudomallei antigen in grown blood culture broth. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 1169–1177. [Google Scholar] [CrossRef]
- Barua, A.; Sathyaseelan, K. Identification of New PCR Targets and its Validation for Development. Def. Life. Sci. J. 2016, 1, 18. [Google Scholar]
- Lowe, C.-W.; Satterfield, B.A.; Nelson, D.B.; Thiriot, J.D.; Heder, M.J.; March, J.K.; Drake, D.S.; Lew, C.S.; Bunnell, A.J.; Moore, E.S. A Quadruplex Real-Time PCR Assay for the Rapid Detection and Differentiation of the Most Relevant Members of the B. pseudomallei Complex: B. mallei, B. pseudomallei, and B. thailandensis. PLoS ONE 2016, 11, e0164006. [Google Scholar] [CrossRef]
- Sangwichian, O.; Whistler, T.; Nithichanon, A.; Kewcharoenwong, C.; Sein, M.M.; Arayanuphum, C.; Chantratita, N.; Lertmemongkolchai, G. Adapting microarray gene expression signatures for early melioidosis diagnosis. J. Clin. Microbiol. 2020. [Google Scholar] [CrossRef]
- Rizzi, M.C.; Rattanavong, S.; Bouthasavong, L.; Seubsanith, A.; Vongsouvath, M.; Davong, V.; De Silvestri, A.; Manciulli, T.; Newton, P.N.; Dance, D.A. Evaluation of the Active Melioidosis Detect™ test as a point-of-care tool for the early diagnosis of melioidosis: A comparison with culture in Laos. Trans. R. Soc. Trop. Med. Hyg. 2019, 113, 757–763. [Google Scholar] [CrossRef] [PubMed]
- Suttisunhakul, V.; Wuthiekanun, V.; Brett, P.J.; Khusmith, S.; Day, N.P.; Burtnick, M.N.; Limmathurotsakul, D.; Chantratita, N. Development of rapid enzyme-linked immunosorbent assays for detection of antibodies to Burkholderia pseudomallei. J. Clin. Microbiol. 2016, 54, 1259–1268. [Google Scholar] [CrossRef]
- Felgner, P.L.; Kayala, M.A.; Vigil, A.; Burk, C.; Nakajima-Sasaki, R.; Pablo, J.; Molina, D.M.; Hirst, S.; Chew, J.S.; Wang, D. A Burkholderia pseudomallei protein microarray reveals serodiagnostic and cross-reactive antigens. Proc. Natl. Acad. Sci. USA 2009, 106, 13499–13504. [Google Scholar] [CrossRef] [PubMed]
- Kritsiriwuthinan, K.; Wajanarogana, S.; Choosang, K.; Homsian, J.; Rerkthanom, S. Production and evaluation of recombinant Burkholderia pseudomallei GroEL and OmpA proteins for serodiagnosis of melioidosis. Acta Trop. 2018, 178, 333–339. [Google Scholar] [CrossRef]
- Abby, S.S.; Rocha, E.P. The non-flagellar type III secretion system evolved from the bacterial flagellum and diversified into host-cell adapted systems. PLoS Genet. 2012, 8, e1002983. [Google Scholar] [CrossRef]
- Holden, M.T.; Titball, R.W.; Peacock, S.J.; Cerdeño-Tárraga, A.M.; Atkins, T.; Crossman, L.C.; Pitt, T.; Churcher, C.; Mungall, K.; Bentley, S.D. Genomic plasticity of the causative agent of melioidosis, Burkholderia pseudomallei. Proc. Natl. Acad. Sci. USA 2004, 101, 14240–14245. [Google Scholar] [CrossRef] [PubMed]
- Stevens, M.P.; Wood, M.W.; Taylor, L.A.; Monaghan, P.; Hawes, P.; Jones, P.W.; Wallis, T.S.; Galyov, E.E. An Inv/Mxi-Spa-like type III protein secretion system in Burkholderia pseudomallei modulates intracellular behaviour of the pathogen. Mol. Microbiol. 2002, 46, 649–659. [Google Scholar] [CrossRef]
- Zilkenat, S.; Franz-Wachtel, M.; Stierhof, Y.-D.; Galán, J.E.; Macek, B.; Wagner, S. Determination of the stoichiometry of the complete bacterial type III secretion needle complex using a combined quantitative proteomic approach. Mol. Cell. Proteom. 2016, 15, 1598–1609. [Google Scholar] [CrossRef] [PubMed]
- Vander Broek, C.W.; Stevens, J.M. Type III secretion in the melioidosis pathogen Burkholderia pseudomallei. Front. Cell. Infect. Microbiol. 2017, 7, 255. [Google Scholar] [CrossRef] [PubMed]
- Bajunaid, W.; Haidar-Ahmad, N.; Kottarampatel, A.H.; Ourida Manigat, F.; Silué, N.F.; Tchagang, C.; Tomaro, K.; Campbell-Valois, F.-X. The T3SS of Shigella: Expression, Structure, Function, and Role in Vacuole Escape. Microorganisms 2020, 8, 1933. [Google Scholar] [CrossRef] [PubMed]
- Erskine, P.; Knight, M.; Ruaux, A.; Mikolajek, H.; Sang, N.W.F.; Withers, J.; Gill, R.; Wood, S.; Wood, M.; Fox, G. High resolution structure of BipD: An invasion protein associated with the type III secretion system of Burkholderia pseudomallei. J. Mol. Biol. 2006, 363, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Roehrich, A.D.; Bordignon, E.; Mode, S.; Shen, D.-K.; Liu, X.; Pain, M.; Murillo, I.; Martinez-Argudo, I.; Sessions, R.B.; Blocker, A.J. Steps for Shigella gatekeeper protein MxiC function in hierarchical type III secretion regulation. J. Biol. Chem. 2017, 292, 1705–1723. [Google Scholar] [CrossRef]
- Stensrud, K.F.; Adam, P.R.; La Mar, C.D.; Olive, A.J.; Lushington, G.H.; Sudharsan, R.; Shelton, N.L.; Givens, R.S.; Picking, W.L.; Picking, W.D. Deoxycholate interacts with IpaD of Shigella flexneri in inducing the recruitment of IpaB to the type III secretion apparatus needle tip. J. Biol. Chem. 2008, 283, 18646–18654. [Google Scholar] [CrossRef]
- Wang, Y.; Nordhues, B.A.; Zhong, D.; De Guzman, R.N. NMR characterization of the interaction of the Salmonella type III secretion system protein SipD and bile salts. Biochemistry 2010, 49, 4220–4226. [Google Scholar] [CrossRef][Green Version]
- Knight, M.; Ruaux, A.; Mikolajek, H.; Erskine, P.; Gill, R.; Wood, S.; Wood, M.; Cooper, J. Crystallization and preliminary X-ray diffraction analysis of BipD, a virulence factor from Burkholderia pseudomallei. Acta Crystallogr. Sect. F. Struct. Biol. Cryst. Commun. 2006, 62, 761–764. [Google Scholar] [CrossRef]
- Pal, M.; Erskine, P.; Gill, R.; Wood, S.; Cooper, J. Near-atomic resolution analysis of BipD, a component of the type III secretion system of Burkholderia pseudomallei. Acta Crystallogr. Sect. F. Struct. Biol. Cryst. Commun. 2010, 66, 990–993. [Google Scholar] [CrossRef]
- Johnson, S.; Roversi, P.; Espina, M.; Olive, A.; Deane, J.E.; Birket, S.; Field, T.; Picking, W.D.; Blocker, A.J.; Galyov, E.E. Self-chaperoning of the type III secretion system needle tip proteins IpaD and BipD. J. Biol. Chem. 2007, 282, 4035–4044. [Google Scholar] [CrossRef]
- Pilatz, S.; Breitbach, K.; Hein, N.; Fehlhaber, B.; Schulze, J.; Brenneke, B.; Eberl, L.; Steinmetz, I. Identification of Burkholderia pseudomallei genes required for the intracellular life cycle and in vivo virulence. Infect. Immun. 2006, 74, 3576–3586. [Google Scholar] [CrossRef]
- Moule, M.G.; Spink, N.; Willcocks, S.; Lim, J.; Guerra-Assunção, J.A.; Cia, F.; Champion, O.L.; Senior, N.J.; Atkins, H.S.; Clark, T. Characterization of new virulence factors involved in the intracellular growth and survival of Burkholderia pseudomallei. Infect. Immun. 2016, 84, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Kespichayawattana, W.; Rattanachetkul, S.; Wanun, T.; Utaisincharoen, P.; Sirisinha, S. Burkholderia pseudomallei induces cell fusion and actin-associated membrane protrusion: A possible mechanism for cell-to-cell spreading. Infect. Immun. 2000, 68, 5377–5384. [Google Scholar] [CrossRef]
- Stockton, J.L.; Torres, A.G. Multinucleated Giant Cell Formation as a Portal to Chronic Bacterial Infections. Microorganisms 2020, 8, 1637. [Google Scholar] [CrossRef]
- Gong, L.; Cullinane, M.; Treerat, P.; Ramm, G.; Prescott, M.; Adler, B.; Boyce, J.D.; Devenish, R.J. The Burkholderia pseudomallei type III secretion system and BopA are required for evasion of LC3-associated phagocytosis. PLoS ONE 2011, 6, e17852. [Google Scholar] [CrossRef]
- Stevens, M.P.; Haque, A.; Atkins, T.; Hill, J.; Wood, M.W.; Easton, A.; Nelson, M.; Underwood-Fowler, C.; Titball, R.W.; Bancroft, G.J. Attenuated virulence and protective efficacy of a Burkholderia pseudomallei bsa type III secretion mutant in murine models of melioidosis. Microbiology 2004, 150, 2669–2676. [Google Scholar] [CrossRef] [PubMed]
- Stevens, M.P.; Friebel, A.; Taylor, L.A.; Wood, M.W.; Brown, P.J.; Hardt, W.-D.; Galyov, E.E. A Burkholderia pseudomallei type III secreted protein, BopE, facilitates bacterial invasion of epithelial cells and exhibits guanine nucleotide exchange factor activity. J. Bacteriol. 2003, 185, 4992–4996. [Google Scholar] [CrossRef]
- Burtnick, M.N.; Brett, P.J.; Nair, V.; Warawa, J.M.; Woods, D.E.; Gherardini, F.C. Burkholderia pseudomallei type III secretion system mutants exhibit delayed vacuolar escape phenotypes in RAW 264.7 murine macrophages. Infect. Immun. 2008, 76, 2991–3000. [Google Scholar] [CrossRef]
- Chantratita, N.; Wuthiekanun, V.; Thanwisai, A.; Limmathurotsakul, D.; Cheng, A.C.; Chierakul, W.; Day, N.P.; Peacock, S.J. Accuracy of enzyme-linked immunosorbent assay using crude and purified antigens for serodiagnosis of melioidosis. Clin. Vaccine Immunol. 2007, 14, 110–113. [Google Scholar] [CrossRef] [PubMed]
- Druar, C.; Yu, F.; Barnes, J.L.; Okinaka, R.T.; Chantratita, N.; Beg, S.; Stratilo, C.W.; Olive, A.J.; Soltes, G.; Russell, M.L. Evaluating Burkholderia pseudomallei Bip proteins as vaccines and Bip antibodies as detection agents. FEMS Immunol. Med. Microbiol. 2008, 52, 78–87. [Google Scholar] [CrossRef]
- Allwood, E.M.; Logue, C.-A.; Hafner, G.J.; Ketheesan, N.; Norton, R.E.; Peak, I.R.; Beacham, I.R. Evaluation of recombinant antigens for diagnosis of melioidosis. FEMS Immunol. Med. Microbiol. 2008, 54, 144–153. [Google Scholar] [CrossRef][Green Version]
- Chuah, S.C.; Gilmore, G.; Norton, R.E. Rapid serological diagnosis of melioidosis: An evaluation of a prototype immunochromatographic test. Pathology 2005, 37, 169–171. [Google Scholar] [CrossRef]
- Hii, S.Y.F.; Ali, N.A.; Ahmad, N.; Amran, F. Comparison of in-house IgM and IgG ELISAs for the serodiagnosis of melioidosis in Malaysia. J. Med. Microbiol. 2017, 66, 1623. [Google Scholar] [CrossRef] [PubMed]
- Visutthi, M.; Jitsurong, S.; Chotigeat, W. Production and purification of Burkholderia pseudomallei BipD protein. Southeast. Asian J. Trop. Med. Public Health 2008, 39, 109. [Google Scholar]
- Castillo, J.; Gáspár, S.; Leth, S.; Niculescu, M.; Mortari, A.; Bontidean, I.; Soukharev, V.; Dorneanu, S.; Ryabov, A.; Csöregi, E. Biosensors for life quality: Design, development and applications. Sens. Actuators B Chem. 2004, 102, 179–194. [Google Scholar] [CrossRef]
- Dawan, S.; Kanatharana, P.; Chotigeat, W.; Jitsurong, S.; Thavarungkul, P. Surface plasmon resonance immunosensor for rapid and specific diagnosis of melioidosis antibody. Southeast. Asian J. Trop. Med. Public Health 2011, 42, 1168. [Google Scholar]
- Ellington, A.D.; Szostak, J.W. In vitro selection of RNA molecules that bind specific ligands. Nature 1990, 346, 818–822. [Google Scholar] [CrossRef]
- Darmostuk, M.; Rimpelova, S.; Gbelcova, H.; Ruml, T. Current approaches in SELEX: An update to aptamer selection technology. Biotechnol. Adv. 2015, 33, 1141–1161. [Google Scholar] [CrossRef]
- Siddiqui, S.; Yuan, J. Binding Characteristics Study of DNA based Aptamers for E. coli O157: H7. Molecules 2021, 26, 204. [Google Scholar] [CrossRef]
- Nosaz, Z.; Rasoulinejad, S.; Mousavi Gargari, S. Development of a DNA aptamer to detect Brucella abortus and Brucella melitensis through cell SELEX. Iran J. Vet. Res. 2020, 21, 294–300. [Google Scholar]
- Su, C.-H.; Tsai, M.-H.; Lin, C.-Y.; Ma, Y.-D.; Wang, C.-H.; Chung, Y.-D.; Lee, G.-B. Dual aptamer assay for detection of Acinetobacter baumannii on an electromagnetically-driven microfluidic platform. Biosens. Bioelectron. 2020, 159, 112148. [Google Scholar] [CrossRef]
- Amraee, M.; Oloomi, M.; Yavari, A.; Bouzari, S. DNA aptamer identification and characterization for E. coli O157 detection using cell based SELEX method. Anal. Biochem. 2017, 536, 36–44. [Google Scholar] [CrossRef]
- Zhu, Z.; Song, Y.; Li, C.; Zou, Y.; Zhu, L.; An, Y.; Yang, C.J. Monoclonal surface display SELEX for simple, rapid, efficient, and cost-effective aptamer enrichment and identification. Anal. Chem. 2014, 86, 5881–5888. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Wang, L.; Li, J.; Fan, C.; Zhao, J. Aptamer-based biosensors. TrAC Trends Anal. Chem. 2008, 27, 108–117. [Google Scholar] [CrossRef]
- Yoo, H.; Jo, H.; Oh, S.S. Detection and beyond: Challenges and advances in aptamer-based biosensors. Mater. Adv. 2020, 1, 2663–2687. [Google Scholar] [CrossRef]
- Alizadeh, N.; Memar, M.Y.; Moaddab, S.R.; Kafil, H.S. Aptamer-assisted novel technologies for detecting bacterial pathogens. Biomed. Pharmacother. 2017, 93, 737–745. [Google Scholar] [CrossRef]
- Majdinasab, M.; Hayat, A.; Marty, J.L. Aptamer-based assays and aptasensors for detection of pathogenic bacteria in food samples. TrAC Trends Anal. Chem. 2018, 107, 60–77. [Google Scholar] [CrossRef]
- Yi-Xian, W.; Zun-Zhong, Y.; Cheng-Yan, S.; Yi-Bin, Y. Application of aptamer based biosensors for detection of pathogenic microorganisms. Chin. J. Anal. Chem. 2012, 40, 634–642. [Google Scholar]
- Gnanam, A.J.; Hall, B.; Shen, X.; Piasecki, S.; Vernados, A.; Galyov, E.E.; Smither, S.J.; Kitto, G.B.; Titball, R.W.; Ellington, A.D.; et al. Development of aptamers specific for potential diagnostic targets in Burkholderia pseudomallei. Trans. R. Soc. Trop. Med. Hyg. 2008, 102, S55–S57. [Google Scholar] [CrossRef]
- Zhu, Q.; Liu, G.; Kai, M.J.M. DNA aptamers in the diagnosis and treatment of human diseases. Molecules 2015, 20, 20979–20997. [Google Scholar] [CrossRef]
- Zhu, Q.; Shibata, T.; Kabashima, T.; Kai, M. Inhibition of HIV-1 protease expression in T cells owing to DNA aptamer-mediated specific delivery of siRNA. Eur. J. Med. Chem. 2012, 56, 396–399. [Google Scholar] [CrossRef] [PubMed]
- Buglak, A.A.; Samokhvalov, A.V.; Zherdev, A.V.; Dzantiev, B.B. Methods and Applications of In Silico Aptamer Design and Modeling. Int. J. Mol. Sci. 2020, 21, 8420. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Residues of BipD | Structural Domains |
|---|---|
| 36–43 | α-helix (helix 1) |
| 47–63 | α-helix (helix 2) |
| 64–81 | β-hairpin (β1 and β2) |
| 82–111 | α-helix (helix 3) runs anti-parallel to helix 2 |
| 128–170 | α-helix (helix 4) runs anti-parallel to helix 3 |
| 171–183 | β-hairpin (β3 and β4) |
| 184–196 | α-helix (helix 5) runs anti-parallel to helix 4 |
| 197–203 | β-hairpin (β5), last three residues are part of a three-stranded β-sheet with β6 and β7 |
| 209–216 | α-helix (helix 6) |
| 220–230 | β-hairpin (β6 and β7) |
| 233–241 | α-helix (helix 7) runs anti-parallel to helix 5 |
| 246–250 | β-hairpin (β8) forms a three-stranded β-sheet with β3 and β4 |
| 251–301 | α-helix (helix 8) |
| Wild-Type Strain of BipD Mutant | In Vitro Growth Rate in LB at 37 °C | Escape from the Phagosomes of Macrophage Cells | Intracellular Replication | Invasion of Cultured HeLa Cells | Induced Membrane Protrusions and Actin Tails in Macrophage Cells | Association of Intracellular Bacteria with LAMP-1-Containing Vacuoles in Macrophage Cells | Refs. |
|---|---|---|---|---|---|---|---|
| 10276 | No effect | ND | Marked reduction in intracellular replication | The invasion occurred at a low frequency | No protrusions or actin rearrangements | Increased association with LAMP-1-containing vacuoles | [40] |
| 10276 | ND a | ND | ND | Highly significant reduction in invasion | ND | ND | [57] |
| 576 | No effect | ND | Low replication of the bacteria in the liver and spleen | ND | ND | ND | [56] |
| K96243 | ND | Unable to escape from phagosomes and showed a high level of co-localization with LC3 | ND | ND | No formation of actin tails | Increased levels of co-localization with LAMP-1 | [55] |
| Methods | BipD | Population | Sensitivity (%) | Specificity (%) | Refs. |
|---|---|---|---|---|---|
| ELISA | Histidine-BipD (His–BipD) | Malaysia, Thailand, and Australia | 42 | 100 | [61] |
| ELISA | His–BipD | Thailand and Australia | 63–75 | 61–64 | [60] |
| Immunoblot | Glutathione S-transferases-BipD (GST–BipD) BipD | Thailand | 78 100 | 90 91.1 | [64] |
| Immunoblot | GST–BipD | ND a | ND | ND | [40] |
| Surface plasmon resonance (SPR) | BipD | Thailand | 100 | 100 | [66] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Selvam, K.; Khalid, M.F.; Mustaffa, K.M.F.; Harun, A.; Aziah, I. BipD of Burkholderia pseudomallei: Structure, Functions, and Detection Methods. Microorganisms 2021, 9, 711. https://doi.org/10.3390/microorganisms9040711
Selvam K, Khalid MF, Mustaffa KMF, Harun A, Aziah I. BipD of Burkholderia pseudomallei: Structure, Functions, and Detection Methods. Microorganisms. 2021; 9(4):711. https://doi.org/10.3390/microorganisms9040711
Chicago/Turabian StyleSelvam, Kasturi, Muhammad Fazli Khalid, Khairul Mohd Fadzli Mustaffa, Azian Harun, and Ismail Aziah. 2021. "BipD of Burkholderia pseudomallei: Structure, Functions, and Detection Methods" Microorganisms 9, no. 4: 711. https://doi.org/10.3390/microorganisms9040711
APA StyleSelvam, K., Khalid, M. F., Mustaffa, K. M. F., Harun, A., & Aziah, I. (2021). BipD of Burkholderia pseudomallei: Structure, Functions, and Detection Methods. Microorganisms, 9(4), 711. https://doi.org/10.3390/microorganisms9040711