
The Small RNA RyhB Homologs from Salmonella Typhimurium Restrain the Intracellular Growth and Modulate the SPI-1 Gene Expression within RAW264.7 Macrophages

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Relevant Characteristic (s) | Reference/Source | |
|---|---|---|
| Strain | ||
| WT | Wild type strain of S. Typhimurium SL1344 | [20] |
| ∆ryhB-1 | S. Typhimurium SL1344 lacking ryhB-1 gene | This study |
| ∆ryhB-2 | S. Typhimurium SL1344 lacking ryhB-2 gene | This study |
| ∆ryhB-1 ∆ryhB-2 | S. Typhimurium SL1344 lacking ryhB-1 and ryhB-2 genes | This study |
| Escherichia coli JM109 | Strain used for heterologous expression analyzes by two-plasmid systems | Promega® |
| Plasmid | ||
| pBR322 | ApR, TcR, ColEl Ori | New England Biolabs® |
| pPromRyhB1 | ryhB-1 region of S. Typhimurium cloned into pBR322 | This study |
| pPromRyhB2 | ryhB-2 region of S. Typhimurium cloned into pBR322 | This study |
| pDiGc | bla GFP pBAD DsRed ori f1 AmpR | [19] |
| pBAD-His-Myc A | pBAD expression plasmid, ApR, pBR322 Ori | Invitrogen® |
| pRyhB1 | pBAD-RyhB1 vector, arabinose inducible | This study |
| pRyhB1MUT1 | pBAD-RyhB1 with ryhB-1 mutated at positions 43–48 | This study |
| pRyhB1MUT2 | pBAD-RyhB1 with ryhB-1 mutated at positions 52–55 | This study |
| pRyhB2 | pBAD-RyhB2 vector, arabinose inducible | This study |
| pRyhB2MUT1 | pBAD-RyhB2 with ryhB-2 mutated at positions 44–49 | This study |
| pRyhB2MUT2 | pBAD-RyhB2 with ryhB-2 mutated at positions 53–56 | This study |
| pSF-p15A | pSF-CMV-p15A Ori vector, KmR, p15A Ori | Sigma® |
| pSFp15A-rpsL | pSF-p15A vector with the rpsL promoter | This study |
| pRtsB | pSFp15A-rpsL vector with rtsB (from −20, relative to AUG) under the control of rpsL promoter (30S ribosomal RNA) for its constitutive expression | This study |
| pRtsBMUT | pRtsB vector with rtsB mutated at positions -1 and -5, relative to AUG | This study |
| pSicA | pSFp15A-rpsL vector with sicA (from its transcriptional start site) under the control of rpsL promoter (30S ribosomal RNA) for its constitutive expression | This study |
| pSicAMUT | pSicA vector with sicA mutated at positions -1 and -4, relative to AUG | This study |
3. Results
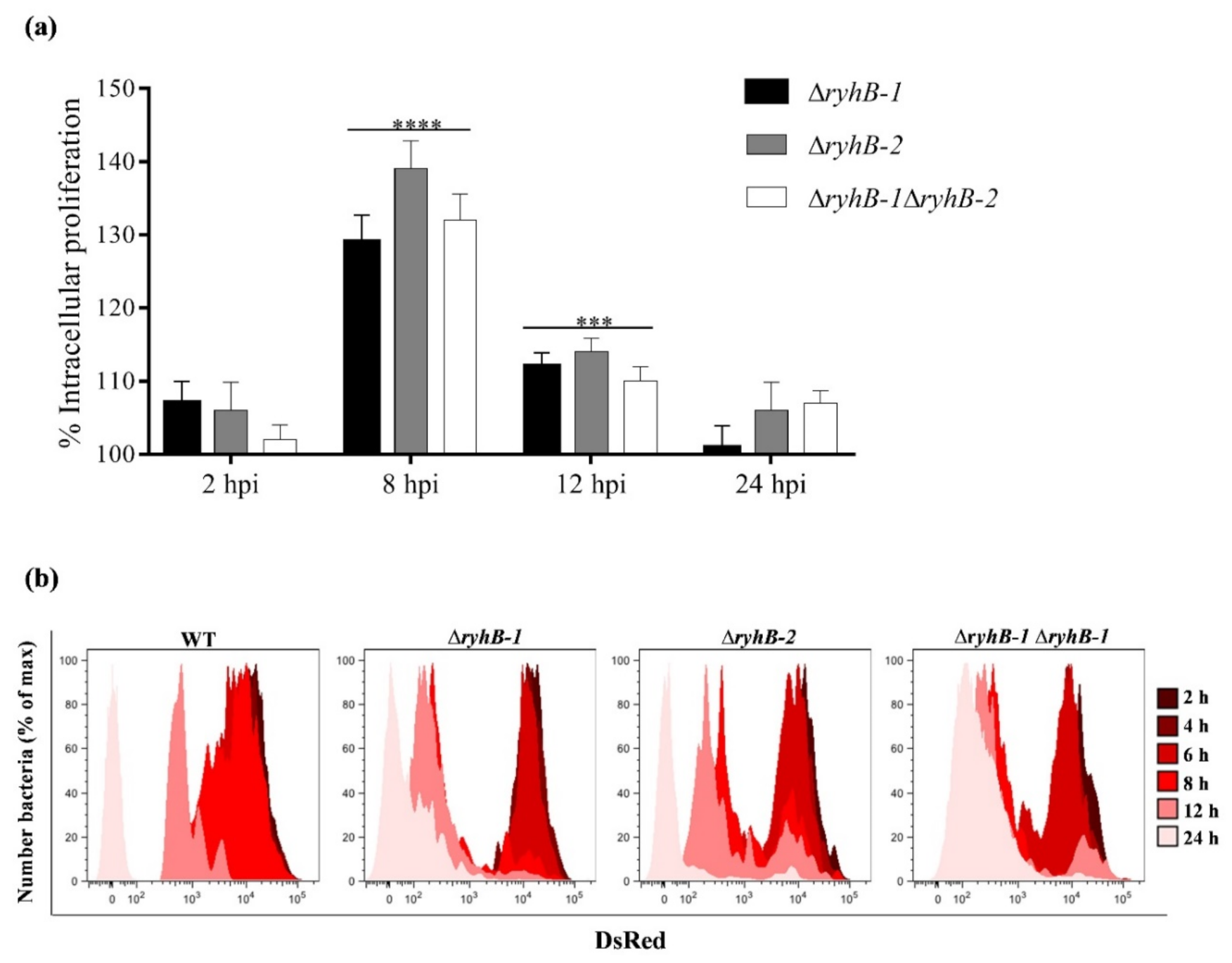
3.1. S. Typhimurium sRNAs RyhB-1 and RyhB-2 Regulate Intracellular Proliferation in Murine Macrophages
3.2. RyhB-1 and RyhB-2 Deletions Result in Metabolically More Active Intracellular Bacteria
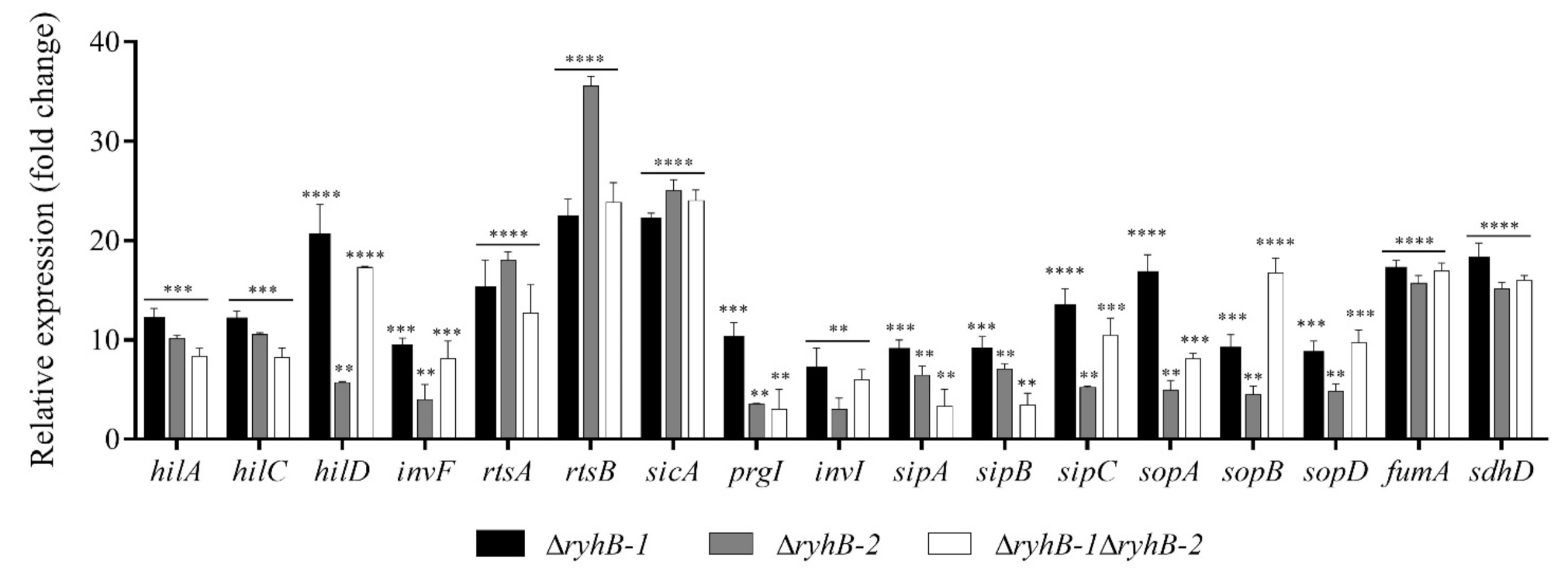
3.3. RyhB-1 and RyhB-2 from S. Typhimurium Affect the Expression of Genes Related to SPI-1 and Metabolism during Infection of RAW264.7 Macrophages
3.4. The RyhB sRNAs Directly Interact with the sicA and rtsB mRNAs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kuhle, V.; Hensel, M. Cellular microbiology of intracellular Salmonella enterica: Functions of the type III secretion system encoded by Salmonella pathogenicity island 2. Cell. Mol. Life Sci. 2004, 61, 2812–2826. [Google Scholar] [CrossRef]
- Haraga, A.; Ohlson, M.B.; Miller, S.I. Salmonellae interplay with host cells. Nat. Rev. Microbiol. 2008, 6, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Heurlier, K.; Williams, F.; Heeb, S.; Dormond, C.; Pessi, G.; Singer, D.; Cámara, M.; Williams, P.; Haas, D. Positive control of swarming, rhamnolipid synthesis, and lipase production by the posttranscriptional RsmA/RsmZ system in Pseudomonas aeruginosa PAO1. J. Bacteriol. 2004, 186, 2936–2945. [Google Scholar] [CrossRef] [PubMed]
- Boisset, S.; Geissmann, T.; Huntzinger, E.; Fechter, P.; Bendridi, N.; Possedko, M.; Chevalier, C.; Helfer, A.C.; Benito, Y.; Jacquier, A.; et al. Staphylococcus aureus RNAIII coordinately represses the synthesis of virulence factors and the transcription regulator Rot by an antisense mechanism. Genes Dev. 2007, 21, 1353–1366. [Google Scholar] [CrossRef] [PubMed]
- Mraheil, M.A.; Billion, A.; Kuenne, C.; Pischimarov, J.; Kreikemeyer, B.; Engelmann, S.; Hartke, A.; Giard, J.C.; Rupnik, M.; Vorwerk, S.; et al. Comparative genome-wide analysis of small RNAs of major Gram-positive pathogens: From identification to application. Microb. Biotechnol. 2010, 3, 658–676. [Google Scholar] [CrossRef] [PubMed]
- Michaux, C.; Verneuil, N.; Hartke, A.; Giard, J.C. Physiological roles of small RNA molecules. Microbiology 2014, 160, 1007–1019. [Google Scholar] [CrossRef]
- Srikumar, S.; Kröger, C.; Hébrard, M.; Colgan, A.; Owen, S.V.; Sivasankaran, S.K.; Cameron, A.D.; Hokamp, K.; Hinton, J.C. RNA-seq Brings New Insights to the Intra-Macrophage Transcriptome of Salmonella Typhimurium. PLoS Pathog. 2015, 11, e1005262. [Google Scholar] [CrossRef]
- Padalon-Brauch, G.; Hershberg, R.; Elgrably-Weiss, M.; Baruch, K.; Rosenshine, I.; Margalit, H.; Altuvia, S. Small RNAs encoded within genetic islands of Salmonella typhimurium show host-induced expression and role in virulence. Nucleic Acids. Res. 2008, 36, 1913–1927. [Google Scholar] [CrossRef]
- Ortega, A.D.; Gonzalo-Asensio, J.; García-del Portillo, F. Dynamics of Salmonella small RNA expression in non-growing bacteria located inside eukaryotic cells. RNA Biol. 2012, 9, 469–488. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.N.; Kwon, Y.M. Genetic and phenotypic characterization of the RyhB regulon in Salmonella Typhimurium. Microbiol. Res. 2013, 168, 41–49. [Google Scholar] [CrossRef]
- Leclerc, J.M.; Dozois, C.M.; Daigle, F. Role of the Salmonella enterica serovar Typhi Fur regulator and small RNAs RfrA and RfrB in iron homeostasis and interaction with host cells. Microbiology 2013, 159, 591–602. [Google Scholar] [CrossRef]
- Calderón, I.L.; Morales, E.H.; Collao, B.; Calderón, P.F.; Chahuán, C.A.; Acuña, L.G.; Gil, F.; Saavedra, C.P. Role of Salmonella Typhimurium small RNAs RyhB-1 and RyhB-2 in the oxidative stress response. Res. Microbiol. 2014, 165, 30–40. [Google Scholar] [CrossRef]
- Calderón, P.F.; Morales, E.H.; Acuña, L.G.; Fuentes, D.N.; Gil, F.; Porwollik, S.; McClelland, M.; Saavedra, C.P.; Calderón, I.L. The small RNA RyhB homologs from Salmonella Typhimurium participate in the response to S-nitrosoglutathione-induced stress. Biochem. Biophys. Res. Commun. 2014, 45, 641–645. [Google Scholar] [CrossRef]
- Datsenko, K.A.; Wanner, B.L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640–6645. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, D.N.; Calderón, P.F.; Acuña, L.G.; Rodas, P.I.; Paredes-Sabja, D.; Fuentes, J.A.; Gil, F.; Calderón, I.L. Motility modulation by the small non-coding RNA SroC in Salmonella Typhimurium. FEMS Microbiol Lett. 2015, 362, fnv135. [Google Scholar] [CrossRef]
- Papenfort, K.; Pfeiffer, V.; Mika, F.; Lucchini, S.; Hinton, J.C.; Vogel, J. SigmaE-dependent small RNAs of Salmonella respond to membrane stress by accelerating global omp mRNA decay. Mol. Microbiol. 2006, 62, 1674–1688. [Google Scholar] [CrossRef]
- Acuña, L.G.; Barros, M.J.; Peñaloza, D.; Rodas, P.I.; Paredes-Sabja, D.; Fuentes, J.A.; Gil, F.; Calderón, I.L. A feed-forward loop between SroC and MgrR small RNAs modulates the expression of eptB and the susceptibility to polymyxin B in Salmonella Typhimurium. Microbiology. 2016, 162, 1996–2004. [Google Scholar] [CrossRef]
- Acuña, L.G.; Barros, M.J.; Montt, F.; Peñaloza, D.; Núñez, P.; Valdés, I.; Gil, F.; Fuentes, J.A.; Calderón, I.L. Participation of two sRNA RyhB homologs from the fish pathogen Yersinia ruckeri in bacterial physiology. Microbiol. Res. 2021, 242, 126629. [Google Scholar] [CrossRef] [PubMed]
- Helaine, S.; Thompson, J.A.; Watson, K.G.; Liu, M.; Boyle, C.; Holden, D.W. Dynamics of intracellular bacterial replication at the single cell level. Proc. Natl. Acad. Sci. USA 2010, 107, 3746–3751. [Google Scholar] [CrossRef] [PubMed]
- Acuña, L.G.; Barros, M.J.; Nuñez, P.; Peñaloza, D.; Montt, F.; Pedraza, D.; Crossley, K.; Gil, F.; Fuentes, J.A.; Calderón, I.L. The cis-encoded antisense RNA IsrA from Salmonella Typhimurium represses the expression of STM0294.1n (iasE), an SOS-induced gene coding for an endoribonuclease activity. Biochem. Biophys. Res. Commun. 2020, 526, 706–712. [Google Scholar] [CrossRef] [PubMed]
- Lou, L.; Zhang, P.; Piao, R.; Wang, Y. Salmonella Pathogenicity Island 1 (SPI-1) and Its Complex Regulatory Network. Front. Cell. Infect. Microbiol. 2019, 9, 270. [Google Scholar] [CrossRef]
- Busch, A.; Richter, A.S.; Backofen, R. IntaRNA: Efficient prediction of bacterial sRNA targets incorporating target site accessibility and seed regions. Bioinformatics 2008, 24, 2849–2856. [Google Scholar] [CrossRef] [PubMed]
- Wright, P.R.; Georg, J.; Mann, M.; Sorescu, D.A.; Richter, A.S.; Lott, S.; Kleinkauf, R.; Hess, W.R.; Backofen, R. CopraRNA and IntaRNA: Predicting small RNA targets, networks and interaction domains. Nucleic. Acids. Res. 2014, 42, W119–W123. [Google Scholar] [CrossRef] [PubMed]
- Mann, M.; Wright, P.R.; Backofen, R. IntaRNA 2.0: Enhanced and customizable prediction of RNA-RNA interactions. Nucleic. Acids. Res. 2017, 45, W435–W439. [Google Scholar] [CrossRef]
- Papenfort, K.; Vogel, J. Multiple target regulation by small noncoding RNAs rewires gene expression at the post-transcriptional level. Res. Microbiol. 2009, 160, 278–287. [Google Scholar] [CrossRef]
- Hébrard, M.; Kröger, C.; Srikumar, S.; Colgan, A.; Händler, K.; Hinton, J.C. sRNAs and the virulence of Salmonella enterica serovar Typhimurium. RNA Biol. 2012, 9, 437–445. [Google Scholar] [CrossRef]
- Chakravarty, S.; Massé, E. RNA-Dependent Regulation of Virulence in Pathogenic Bacteria. Front. Cell. Infect. Microbiol. 2019, 9, 337. [Google Scholar] [CrossRef] [PubMed]
- Gonzalo-Asensio, J.; Ortega, A.D.; Rico-Pérez, G.; Pucciarelli, M.G.; García-Del Portillo, F. A novel antisense RNA from the Salmonella virulence plasmid pSLT expressed by non-growing bacteria inside eukaryotic cells. PLoS ONE 2013, 8, e77939. [Google Scholar] [CrossRef] [PubMed]
- Núñez-Hernández, C.; Tierrez, A.; Ortega, A.D.; Pucciarelli, M.G.; Godoy, M.; Eisman, B.; Casadesús, J.; García-del Portillo, F. Genome expression analysis of nonproliferating intracellular Salmonella enterica serovar Typhimurium unravels an acid pH-dependent PhoP-PhoQ response essential for dormancy. Infect. Immun. 2013, 81, 154–165. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, G.; Chen, C.; Portnoy, D.A. Strategies Used by Bacteria to Grow in Macrophages. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef]
- Leung, K.Y.; Finlay, B.B. Intracellular replication is essential for the virulence of Salmonella Typhimurium. Proc. Natl. Acad. Sci. USA 1991, 88, 11470–11474. [Google Scholar] [CrossRef] [PubMed]
- Tierrez, A.; García-del Portillo, F. New concepts in Salmonella virulence: The importance of reducing the intracellular growth rate in the host. Cell. Microbiol. 2005, 7, 901–909. [Google Scholar] [CrossRef]
- Albaghdadi, H.; Robinson, N.; Finlay, B.; Krishnan, L.; Sad, S. Selectively reduced intracellular proliferation of Salmonella enterica serovar Typhimurium within APCs limits antigen presentation and development of a rapid CD8 T cell response. J. Immunol. 2009, 183, 3778–3787. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, S.; Björkman, J.; Borg, S.; Syk, A.; Pettersson, S.; Andersson, D.I.; Rhen, M. Salmonella typhimurium mutants that downregulate phagocyte nitric oxide production. Cell. Microbiol. 2000, 2, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Kröger, C.; Colgan, A.; Srikumar, S.; Händler, K.; Sivasankaran, S.K.; Hammarlöf, D.L.; Canals, R.; Grissom, J.E.; Conway, T.; Hokamp, K.; et al. An infection-relevant transcriptomic compendium for Salmonella enterica serovar Typhimurium. Cell. Host. Microbe. 2013, 14, 683–695. [Google Scholar] [CrossRef]
- Ellis, M.J.; Tsai, C.N.; Johnson, J.W.; French, S.; Elhenawy, W.; Porwollik, S.; Andrews-Polymenis, H.; McClelland, M.; Magolan, J.; Coombes, B.K.; et al. A macrophage-based screen identifies antibacterial compounds selective for intracellular Salmonella Typhimurium. Nat. Commun. 2019, 10, 197. [Google Scholar] [CrossRef]
- Massé, E.; Gottesman, S. A small RNA regulates the expression of genes involved in iron metabolism in Escherichia coli. Proc. Natl. Acad. Sci. USA 2002, 99, 4620–4625. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, S.; Wu, N.; Yuan, Y.; Zhang, W.; Zhang, Y. Small Non-coding RNA RyhB Mediates Persistence to Multiple Antibiotics and Stresses in Uropathogenic Escherichia coli by Reducing Cellular Metabolism. Front. Microbiol. 2018, 9, 136. [Google Scholar] [CrossRef]
- Eriksson, S.; Lucchini, S.; Thompson, A.; Rhen, M.; Hinton, J.C. Unravelling the biology of macrophage infection by gene expression profiling of intracellular Salmonella enterica. Mol. Microbiol. 2003, 47, 103–118. [Google Scholar] [CrossRef]
- Vazquez-Torres, A.; Fang, F.C. Oxygen-dependent anti-Salmonella activity of macrophages. Trends. Microbiol. 2001, 9, 29–33. [Google Scholar] [CrossRef]
- Benjamin, J.A.; Desnoyers, G.; Morissette, A.; Salvail, H.; Massé, E. Dealing with oxidative stress and iron starvation in microorganisms: An overview. Can. J. Physiol. Pharmacol. 2010, 88, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Imlay, J.A. The molecular mechanisms and physiological consequences of oxidative stress: Lessons from a model bacterium. Nat. Rev. Microbiol. 2013, 11, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Ortega, A.D.; Quereda, J.J.; Pucciarelli, M.G.; García-del Portillo, F. Non-coding RNA regulation in pathogenic bacteria located inside eukaryotic cells. Front. Cell. Infect. Microbiol. 2014, 4, 162. [Google Scholar] [CrossRef]
- Hurley, D.; McCusker, M.P.; Fanning, S.; Martins, M. Salmonella-host interactions-modulation of the host innate immune system. Front. Immunol. 2014, 5, 481. [Google Scholar] [CrossRef]
- Toledo-Arana, A.; Dussurget, O.; Nikitas, G.; Sesto, N.; Guet-Revillet, H.; Balestrino, D.; Loh, E.; Gripenland, J.; Tiensuu, T.; Vaitkevicius, K.; et al. The Listeria transcriptional landscape from saprophytism to virulence. Nature 2009, 459, 950–956. [Google Scholar] [CrossRef]
- Wurtzel, O.; Sesto, N.; Mellin, J.R.; Karunker, I.; Edelheit, S.; Bécavin, C.; Archambaud, C.; Cossart, P.; Sorek, R. Comparative transcriptomics of pathogenic and non-pathogenic Listeria species. Mol. Syst. Biol. 2012, 8, 583. [Google Scholar] [CrossRef]
- Lin, H.H.; Chen, H.L.; Weng, C.C.; Janapatla, R.P.; Chen, C.L.; Chiu, C.H. Activation of apoptosis by Salmonella pathogenicity island-1 effectors through both intrinsic and extrinsic pathways in Salmonella-infected macrophages. J. Microbiol. Immunol. Infect. 2020, 26. [Google Scholar] [CrossRef]
- Brodsky, I.E.; Medzhitov, R. Pyroptosis: Macrophage suicide exposes hidden invaders. Curr. Biol. 2011, 21, R72–R75. [Google Scholar] [CrossRef]
- Cirillo, D.M.; Valdivia, R.H.; Monack, D.M.; Falkow, S. Macrophage-dependent induction of the Salmonella pathogenicity island 2 type III secretion system and its role in intracellular survival. Mol. Microbiol. 1998, 30, 175–188. [Google Scholar] [CrossRef]
- Hansen-Wester, I.; Hensel, M. Salmonella pathogenicity islands encoding type III secretion systems. Microbes. Infect. 2001, 3, 549–559. [Google Scholar] [CrossRef]
- Monack, D.M.; Navarre, W.W.; Falkow, S. Salmonella-induced macrophage death: The role of caspase-1 in death and inflammation. Microbes. Infect. 2001, 3, 1201–1212. [Google Scholar] [CrossRef] [PubMed]
- Takaya, A.; Suzuki, A.; Kikuchi, Y.; Eguchi, M.; Isogai, E.; Tomoyasu, T.; Yamamoto, T. Derepression of Salmonella pathogenicity island 1 genes within macrophages leads to rapid apoptosis via caspase-1- and caspase-3-dependent pathways. Cell Microbiol. 2005, 7, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Boddicker, J.D.; Jones, B.D. Lon protease activity causes down-regulation of Salmonella pathogenicity island 1 invasion gene expression after infection of epithelial cells. Infect. Immun. 2004, 72, 2002–2013. [Google Scholar] [CrossRef] [PubMed]
- Caswell, C.C.; Oglesby-Sherrouse, A.G.; Murphy, E.R. Sibling rivalry: Related bacterial small RNAs and their redundant and non-redundant roles. Front. Cell Infect. Microbiol. 2014, 4, 151. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.N.; Kwon, Y.M. Identification of target transcripts regulated by small RNA RyhB homologs in Salmonella: RyhB-2 regulates motility phenotype. Microbiol. Res. 2013, 168, 621–629. [Google Scholar] [CrossRef]
- Darwin, K.H.; Miller, V.L. The putative invasion protein chaperone SicA acts together with InvF to activate the expression of Salmonella typhimurium virulence genes. Mol. Microbiol. 2000, 35, 949–960. [Google Scholar] [CrossRef]
- Ellermeier, C.D.; Slauch, J.M. RtsA and RtsB coordinately regulate expression of the invasion and flagellar genes in Salmonella enterica serovar Typhimurium. J. Bacteriol. 2003, 185, 5096–5108. [Google Scholar] [CrossRef] [PubMed]
- Sano, G.; Takada, Y.; Goto, S.; Maruyama, K.; Shindo, Y.; Oka, K.; Matsui, H.; Matsuo, K. Flagella facilitate escape of Salmonella from oncotic macrophages. J. Bacteriol. 2007, 189, 8224–8232. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peñaloza, D.; Acuña, L.G.; Barros, M.J.; Núñez, P.; Montt, F.; Gil, F.; Fuentes, J.A.; Calderón, I.L. The Small RNA RyhB Homologs from Salmonella Typhimurium Restrain the Intracellular Growth and Modulate the SPI-1 Gene Expression within RAW264.7 Macrophages. Microorganisms 2021, 9, 635. https://doi.org/10.3390/microorganisms9030635
Peñaloza D, Acuña LG, Barros MJ, Núñez P, Montt F, Gil F, Fuentes JA, Calderón IL. The Small RNA RyhB Homologs from Salmonella Typhimurium Restrain the Intracellular Growth and Modulate the SPI-1 Gene Expression within RAW264.7 Macrophages. Microorganisms. 2021; 9(3):635. https://doi.org/10.3390/microorganisms9030635
Chicago/Turabian StylePeñaloza, Diego, Lillian G. Acuña, M. José Barros, Paula Núñez, Fernanda Montt, Fernando Gil, Juan A. Fuentes, and Iván L. Calderón. 2021. "The Small RNA RyhB Homologs from Salmonella Typhimurium Restrain the Intracellular Growth and Modulate the SPI-1 Gene Expression within RAW264.7 Macrophages" Microorganisms 9, no. 3: 635. https://doi.org/10.3390/microorganisms9030635
APA StylePeñaloza, D., Acuña, L. G., Barros, M. J., Núñez, P., Montt, F., Gil, F., Fuentes, J. A., & Calderón, I. L. (2021). The Small RNA RyhB Homologs from Salmonella Typhimurium Restrain the Intracellular Growth and Modulate the SPI-1 Gene Expression within RAW264.7 Macrophages. Microorganisms, 9(3), 635. https://doi.org/10.3390/microorganisms9030635