
Ectopic Spacer Acquisition in Streptococcus thermophilus CRISPR3 Array

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Phage and Bacterial Strains
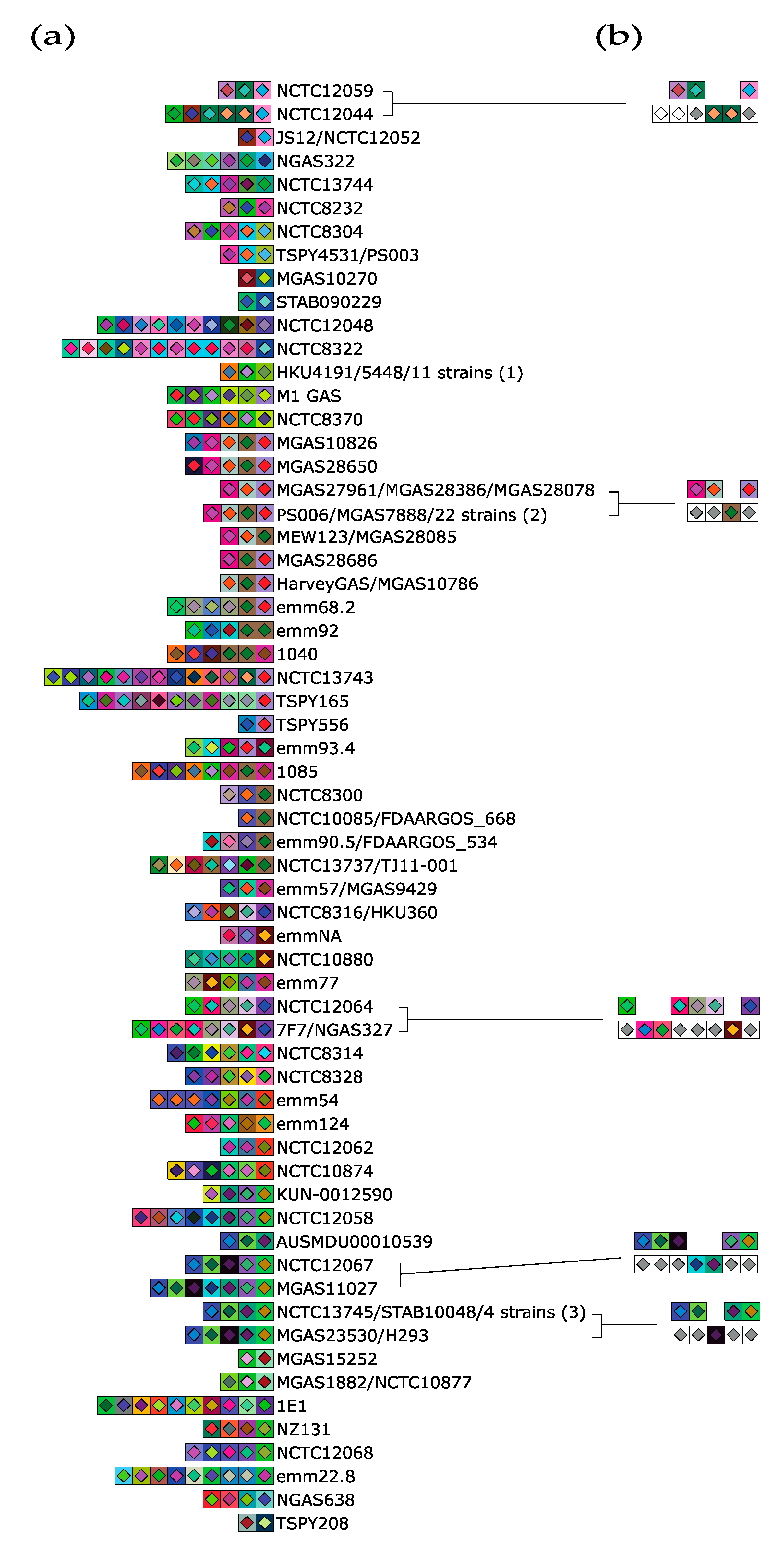
2.2. S. thermophilus and S. pyogenes CRISPR Loci Analysis
2.3. Bacteriophage Insensitive Mutants (BIMs)
3. Results
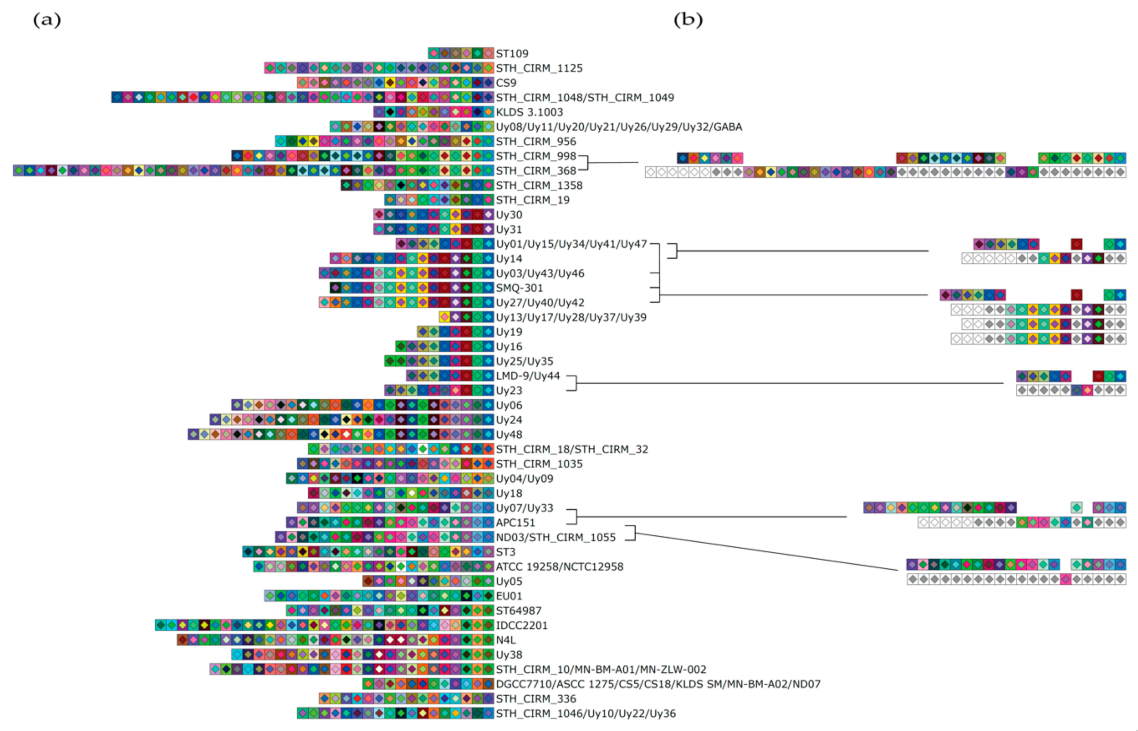
3.1. Evidence of Ectopic Acquisition Events in CRISPR3
3.2. Evaluation of Spacer Acquisition
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hols, P.; Hancy, F.; Fontaine, L.; Grossiord, B.; Prozzi, D.; Leblond-Bourget, N.; Decaris, B.; Bolotin, A.; Delorme, C.; Ehrlich, S.D.; et al. New insights in the molecular biology and physiology of Streptococcus thermophilus revealed by comparative genomics. FEMS Microbiol. Rev. 2005, 29, 435–463. [Google Scholar] [CrossRef]
- Delorme, C. Safety assessment of dairy microorganisms: Streptococcus thermophilus. Int. J. Food Microbiol. 2008, 126, 274–277. [Google Scholar] [CrossRef] [PubMed]
- Iyer, R.; Tomar, S.K.; Uma Maheswari, T.; Singh, R. Streptococcus thermophilus strains: Multifunctional lactic acid bacteria. Int. Dairy J. 2010, 20, 133–141. [Google Scholar] [CrossRef]
- Uriot, O.; Denis, S.; Junjua, M.; Roussel, Y.; Dary-Mourot, A.; Blanquet-Diot, S. Streptococcus thermophilus: From yogurt starter to a new promising probiotic candidate? J. Funct. Foods 2017, 37, 74–89. [Google Scholar] [CrossRef]
- Garneau, J.E.; Moineau, S. Bacteriophages of lactic acid bacteria and their impact on milk fermentations. Microb. Cell Fact. 2011, 10, S20. [Google Scholar] [CrossRef]
- Mahony, J.; Cambillau, C.; van Sinderen, D. Host recognition by lactic acid bacterial phages. FEMS Microbiol. Rev. 2017, 41, S16–S26. [Google Scholar] [CrossRef] [PubMed]
- Lavelle, K.; Martinez, I.; Neve, H.; Lugli, G.A.; Franz, C.M.A.P.; Ventura, M.; Bello, F.D.; van Sinderen, D.; Mahony, J. Biodiversity of Streptococcus thermophilus phages in global dairy fermentations. Viruses 2018, 10, 577. [Google Scholar] [CrossRef]
- Romero, D.A.; Magill, D.; Millen, A.; Horvath, P.; Fremaux, C. Dairy lactococcal and streptococcal phage–host interactions: An industrial perspective in an evolving phage landscape. FEMS Microbiol. Rev. 2020, 44, 909–932. [Google Scholar] [CrossRef]
- Barrangou, R.; Fremaux, C.; Deveau, H.; Richards, M.; Boyaval, P.; Moineau, S.; Romero, D.A.; Horvath, P. CRISPR provides acquired resistance against viruses in prokaryotes. Science 2007, 315, 1709–1712. [Google Scholar] [CrossRef]
- Deveau, H.; Barrangou, R.; Garneau, J.E.; Labonté, J.; Fremaux, C.; Boyaval, P.; Romero, D.A.; Horvath, P.; Moineau, S. Phage response to CRISPR-encoded resistance in Streptococcus thermophilus. J. Bacteriol. 2008, 190, 1390–1400. [Google Scholar] [CrossRef]
- Mills, S.; Griffin, C.; Coffey, A.; Meijer, W.C.; Hafkamp, B.; Ross, R.P. CRISPR analysis of bacteriophage-insensitive mutants (BIMs) of industrial Streptococcus thermophilus– implications for starter design. J. Appl. Microbiol. 2010, 108, 945–955. [Google Scholar] [CrossRef] [PubMed]
- Paez-Espino, D.; Sharon, I.; Morovic, W.; Stahl, B.; Thomas, B.C.; Barrangou, R.; Banfield, J.F. CRISPR immunity drives rapid phage genome evolution in Streptococcus thermophilus. mBio 2015, 6, e00262-15. [Google Scholar] [CrossRef]
- Hao, M.; Cui, Y.; Qu, X. Analysis of CRISPR-Cas system in Streptococcus thermophilus and its application. Front. Microbiol. 2018, 9, 257. [Google Scholar] [CrossRef] [PubMed]
- Naumenko, O.; Skrypkina, I.; Zhukova, Y.; Vakulenko, M.; Kigel, N. Selection and analysis of bacteriophage-insensitive mutants of Streptococcus thermophilus isolated in Ukraine. Int. J. Dairy Technol. 2019, 72, 515–523. [Google Scholar] [CrossRef]
- Cañez, C.; Selle, K.; Goh, Y.J.; Barrangou, R. Outcomes and characterization of chromosomal self-targeting by native CRISPR-Cas systems in Streptococcus thermophilus. FEMS Microbiol. Lett. 2019, 366, fnz105. [Google Scholar] [CrossRef] [PubMed]
- Bolotin, A.; Quinquis, B.; Sorokin, A.; Dusko Ehrlich, S. Clustered regularly interspaced short palindrome repeats (CRISPRs) have spacers of extrachromosomal origin. Microbiology 2005, 151, 2551–2561. [Google Scholar] [CrossRef] [PubMed]
- Pourcel, C.; Salvignol, G.; Vergnaud, G. CRISPR elements in Yersinia pestis acquire new repeats by preferential uptake of bacteriophage DNA, and provide additional tools for evolutionary studies. Microbiology 2005, 151, 653–663. [Google Scholar] [CrossRef] [PubMed]
- Hille, F.; Richter, H.; Wong, S.P.; Bratovič, M.; Ressel, S.; Charpentier, E. The biology of CRISPR-Cas: Backward and forward. Cell 2018, 172, 1239–1259. [Google Scholar] [CrossRef] [PubMed]
- Brouns, S.J.J.; Jore, M.M.; Lundgren, M.; Westra, E.R.; Slijkhuis, R.J.H.; Snijders, A.P.L.; Dickman, M.J.; Makarova, K.S.; Koonin, E.V.; van der Oost, J. Small CRISPR RNAs guide antiviral defense in prokaryotes. Science 2008, 321, 960–964. [Google Scholar] [CrossRef]
- Deltcheva, E.; Chylinski, K.; Sharma, C.M.; Gonzales, K.; Chao, Y.; Pirzada, Z.A.; Eckert, M.R.; Vogel, J.; Charpentier, E. CRISPR RNA maturation by trans-encoded small RNA and host factor RNase III. Nature 2011, 471, 602–607. [Google Scholar] [CrossRef]
- Garneau, J.E.; Dupuis, M.È.; Villion, M.; Romero, D.A.; Barrangou, R.; Boyaval, P.; Fremaux, C.; Horvath, P.; Magadán, A.H.; Moineau, S. The CRISPR/cas bacterial immune system cleaves bacteriophage and plasmid DNA. Nature 2010, 468, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Magadán, A.H.; Dupuis, M.È.; Villion, M.; Moineau, S. Cleavage of phage DNA by the Streptococcus thermophilus CRISPR3-Cas system. PLoS ONE 2012, 7, 40913. [Google Scholar] [CrossRef]
- Gasiunas, G.; Sinkunas, T.; Siksnys, V. Molecular mechanisms of CRISPR-mediated microbial immunity. Cell. Mol. Life Sci. 2014, 71, 449–465. [Google Scholar] [CrossRef] [PubMed]
- Makarova, K.S.; Wolf, Y.I.; Iranzo, J.; Shmakov, S.A.; Alkhnbashi, O.S.; Brouns, S.J.J.; Charpentier, E.; Cheng, D.; Haft, D.H.; Horvath, P.; et al. Evolutionary classification of CRISPR–Cas systems: A burst of class 2 and derived variants. Nat. Rev. Microbiol. 2020, 18, 67–83. [Google Scholar] [CrossRef]
- Nuñez, J.K.; Kranzusch, P.J.; Noeske, J.; Wright, A.V.; Davies, C.W.; Doudna, J.A. Cas1-Cas2 complex formation mediates spacer acquisition during CRISPR-Cas adaptive immunity. Nat. Struct. Mol. Biol. 2014, 21, 528–534. [Google Scholar] [CrossRef]
- Amitai, G.; Sorek, R. CRISPR-Cas adaptation: Insights into the mechanism of action. Nat. Rev. Microbiol. 2016, 14, 67–76. [Google Scholar] [CrossRef]
- Horvath, P.; Romero, D.A.; Coûté-Monvoisin, A.C.; Richards, M.; Deveau, H.; Moineau, S.; Boyaval, P.; Fremaux, C.; Barrangou, R. Diversity, activity, and evolution of CRISPR loci in Streptococcus thermophilus. J. Bacteriol. 2008, 190, 1401–1412. [Google Scholar] [CrossRef] [PubMed]
- Carte, J.; Christopher, R.T.; Smith, J.T.; Olson, S.; Barrangou, R.; Moineau, S.; Glover 3rd, C.V.C.; Graveley, B.R.; Terns, R.M.; Terns, M.P. The three major types of CRISPR-Cas systems function independently in CRISPR RNA biogenesis in Streptococcus thermophilus. Mol. Microbiol. 2014, 93, 98–112. [Google Scholar] [CrossRef]
- Mosterd, C.; Rousseau, G.M.; Moineau, S. A short overview of the CRISPR-Cas adaptation stage. Can. J. Microbiol. 2020, 12, 1–12. [Google Scholar] [CrossRef]
- Makarova, K.S.; Wolf, Y.I.; Alkhnbashi, O.S.; Costa, F.; Shah, S.A.; Saunders, S.J.; Barrangou, R.; Brouns, S.J.J.; Charpentier, E.; Haft, D.H.; et al. An updated evolutionary classification of CRISPR-Cas systems. Nat. Rev. Microbiol. 2015, 13, 722–736. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, M.; Drabavicius, G.; Silanskas, A.; Gasiunas, G.; Siksnys, V.; Wigley, D.B. Structure of the DNA-Bound Spacer Capture Complex of a Type II CRISPR-Cas System. Mol. Cell 2019, 75, 90–101. [Google Scholar] [CrossRef] [PubMed]
- McGinn, J.; Marraffini, L.A. Molecular mechanisms of CRISPR–Cas spacer acquisition. Nat. Rev. Microbiol. 2019, 17, 7–12. [Google Scholar] [CrossRef] [PubMed]
- McGinn, J.; Marraffini, L.A. CRISPR-Cas systems optimize their immune response by specifying the site of spacer integration. Mol. Cell 2016, 64, 616–623. [Google Scholar] [CrossRef]
- Wright, A.V.; Doudna, J.A. Protecting genome integrity during CRISPR immune adaptation. Nat. Struct. Mol. Biol. 2016, 23, 876–883. [Google Scholar] [CrossRef]
- Wright, A.V.; Liu, J.J.; Knott, G.J.; Doxzen, K.W.; Nogales, E.; Doudna, J.A. Structures of the CRISPR genome integration complex. Science 2017, 357, 1113–1118. [Google Scholar] [CrossRef]
- Xiao, Y.; Ng, S.; Hyun Nam, K.; Ke, A. How type II CRISPR-Cas establish immunity through Cas1-Cas2-mediated spacer integration. Nature 2017, 550, 137–141. [Google Scholar] [CrossRef]
- Mosterd, C.; Moineau, S. Characterization of a Type II-A CRISPR-Cas system in Streptococcus mutans. mSphere 2020, 5, e00235-20. [Google Scholar] [CrossRef]
- Achigar, R.; Magadán, A.H.; Tremblay, D.M.; Julia Pianzzola, M.; Moineau, S. Phage-host interactions in Streptococcus thermophilus: Genome analysis of phages isolated in Uruguay and ectopic spacer acquisition in CRISPR array. Sci. Rep. 2017, 7, 43438. [Google Scholar] [CrossRef]
- Hynes, A.P.; Villion, M.; Moineau, S. Adaptation in bacterial CRISPR-Cas immunity can be driven by defective phages. Nat. Commun. 2014, 5, 4399. [Google Scholar] [CrossRef] [PubMed]
- Biswas, A.; Staals, R.H.J.; Morales, S.E.; Fineran, P.C.; Brown, C.M. CRISPRDetect: A flexible algorithm to define CRISPR arrays. BMC Genomics 2016, 17, 356. [Google Scholar] [CrossRef]
- Dion, M.B.; Labrie, S.J.; Shah, S.A.; Moineau, S. Crisprstudio: A user-friendly software for rapid crispr array visualization. Viruses 2018, 10, 602. [Google Scholar] [CrossRef]
- Wei, Y.; Chesne, M.T.; Terns, R.M.; Terns, M.P. Sequences spanning the leader-repeat junction mediate CRISPR adaptation to phage in Streptococcus thermophilus. Nucleic Acids Res. 2015, 43, 1749–1758. [Google Scholar] [CrossRef] [PubMed]
- Hynes, A.P.; Rousseau, G.M.; Lemay, M.-L.; Horvath, P.; Romero, D.A.; Fremaux, C.; Moineau, S. An anti-CRISPR from a virulent streptococcal phage inhibits Streptococcus pyogenes Cas9. Nat. Microbiol. 2017, 2, 1374–1380. [Google Scholar] [CrossRef]
- Landsberger, M.; Gandon, S.; Meaden, S.; Rollie, C.; Chevallereau, A.; Chabas, H.; Buckling, A.; Westra, E.R.; van Houte, S. Anti-CRISPR phages cooperate to overcome CRISPR-Cas immunity. Cell 2018, 174, 908–916. [Google Scholar] [CrossRef] [PubMed]
- Stanley, S.Y.; Maxwell, K.L. Phage-Encoded Anti-CRISPR Defenses. Annu. Rev. Genet. 2018, 52, 445–464. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BIM | Sequence of New Spacer (5′-3′) | Position in CRISPR3 | Protospacer Position in Phage 53 Genome | PAM NGGNG |
|---|---|---|---|---|
| BIM1 | TGATAGTAAAATATTGTCATCATTGAATAC | 6 | None * | None |
| CATTACAGACACAGGAGAAGGCGGCTATTA | 4 | 22837-22866, orf22 | TGGTG | |
| BIM2 | TTATGCAAACGGTGGCCTAGTCCACAAGAA | 4 | 14360-14389, orf14 | CGGCG |
| BIM3 | AGTTGATGGTAAAACGGTGGAATGACCATA | 4 | 31076-31105, orf34 | TGGCG |
| BIM4 | AAACGTCAAAAAAGCTGGTAGTAAGGTCAA | 4 | 11789-11818, orf14 | TGGCG |
| BIM5 | TGTTCAGTATCGTCGACTTCATTCCCCAAA | 4 | 10537-10508, orf14 | CGGCG |
| BIM6 | TGTTCAGTATCGTCGACTTCATTCCCCAAA | 4 | 10537-10508, orf14 | CGGCG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Achigar, R.; Scarrone, M.; Rousseau, G.M.; Philippe, C.; Machado, F.; Duvós, V.; Campot, M.P.; Dion, M.B.; Shao, Y.; Pianzzola, M.J.; et al. Ectopic Spacer Acquisition in Streptococcus thermophilus CRISPR3 Array. Microorganisms 2021, 9, 512. https://doi.org/10.3390/microorganisms9030512
Achigar R, Scarrone M, Rousseau GM, Philippe C, Machado F, Duvós V, Campot MP, Dion MB, Shao Y, Pianzzola MJ, et al. Ectopic Spacer Acquisition in Streptococcus thermophilus CRISPR3 Array. Microorganisms. 2021; 9(3):512. https://doi.org/10.3390/microorganisms9030512
Chicago/Turabian StyleAchigar, Rodrigo, Martina Scarrone, Geneviève M. Rousseau, Cécile Philippe, Felipe Machado, Valentina Duvós, María Pía Campot, Moïra B. Dion, Yuyu Shao, María Julia Pianzzola, and et al. 2021. "Ectopic Spacer Acquisition in Streptococcus thermophilus CRISPR3 Array" Microorganisms 9, no. 3: 512. https://doi.org/10.3390/microorganisms9030512
APA StyleAchigar, R., Scarrone, M., Rousseau, G. M., Philippe, C., Machado, F., Duvós, V., Campot, M. P., Dion, M. B., Shao, Y., Pianzzola, M. J., & Moineau, S. (2021). Ectopic Spacer Acquisition in Streptococcus thermophilus CRISPR3 Array. Microorganisms, 9(3), 512. https://doi.org/10.3390/microorganisms9030512