
Comparison of Common Enrichment Broths Used in Diagnostic Laboratories for Shiga Toxin—Producing Escherichia coli



Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Isolate Selection and Enrichment
2.2. Spiking of Negative Stool
2.3. DNA Extraction and qPCR of Broth Enrichment
2.4. Statistics
3. Results
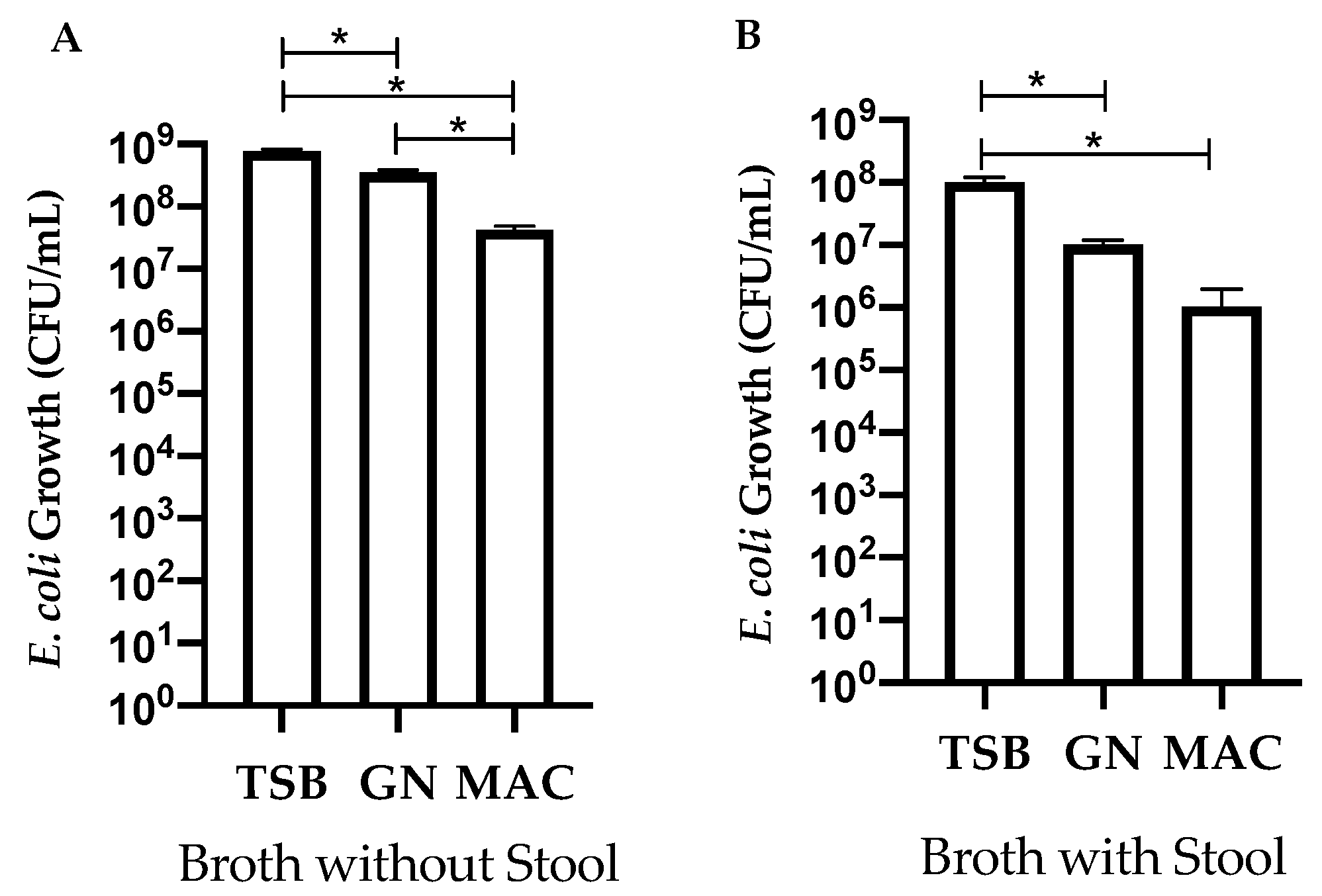
3.1. Comparing Enrichment of STEC in Different Broths with and without the Presence of Stool
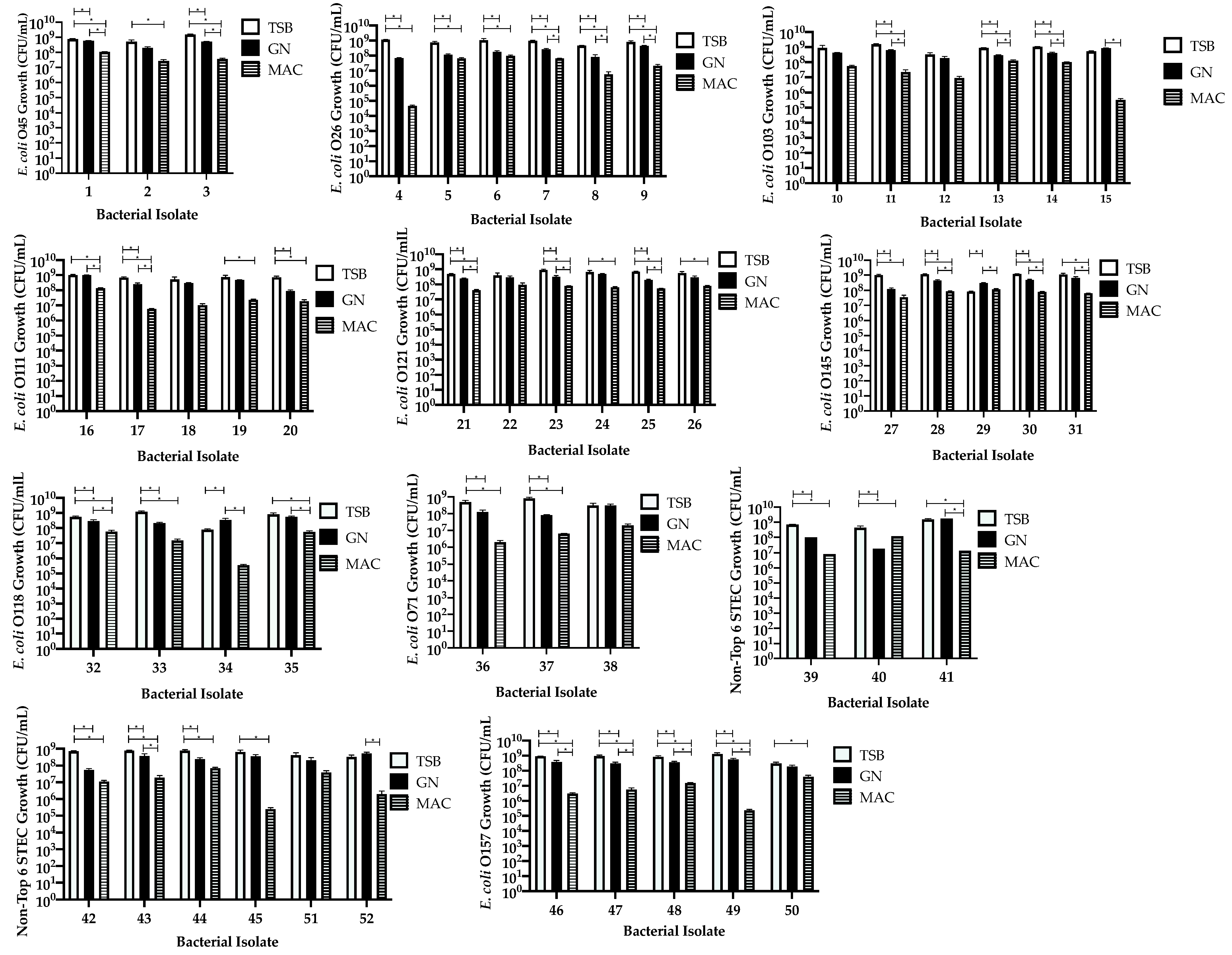
3.2. Comparison of Isolates within Each Serotype Group Independent of Stool
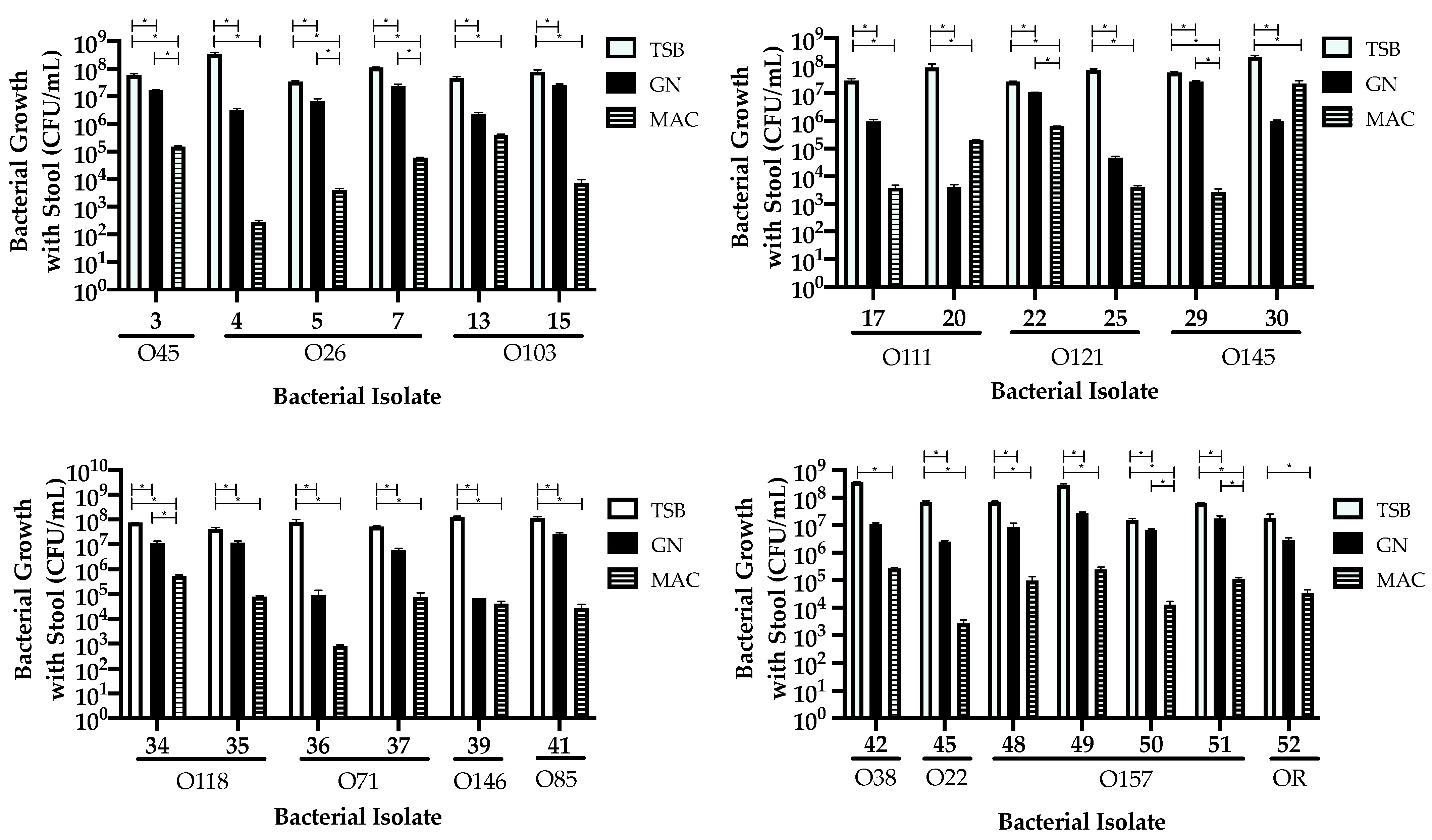
3.3. Comparing Isolate Growth in the Presence of Stool
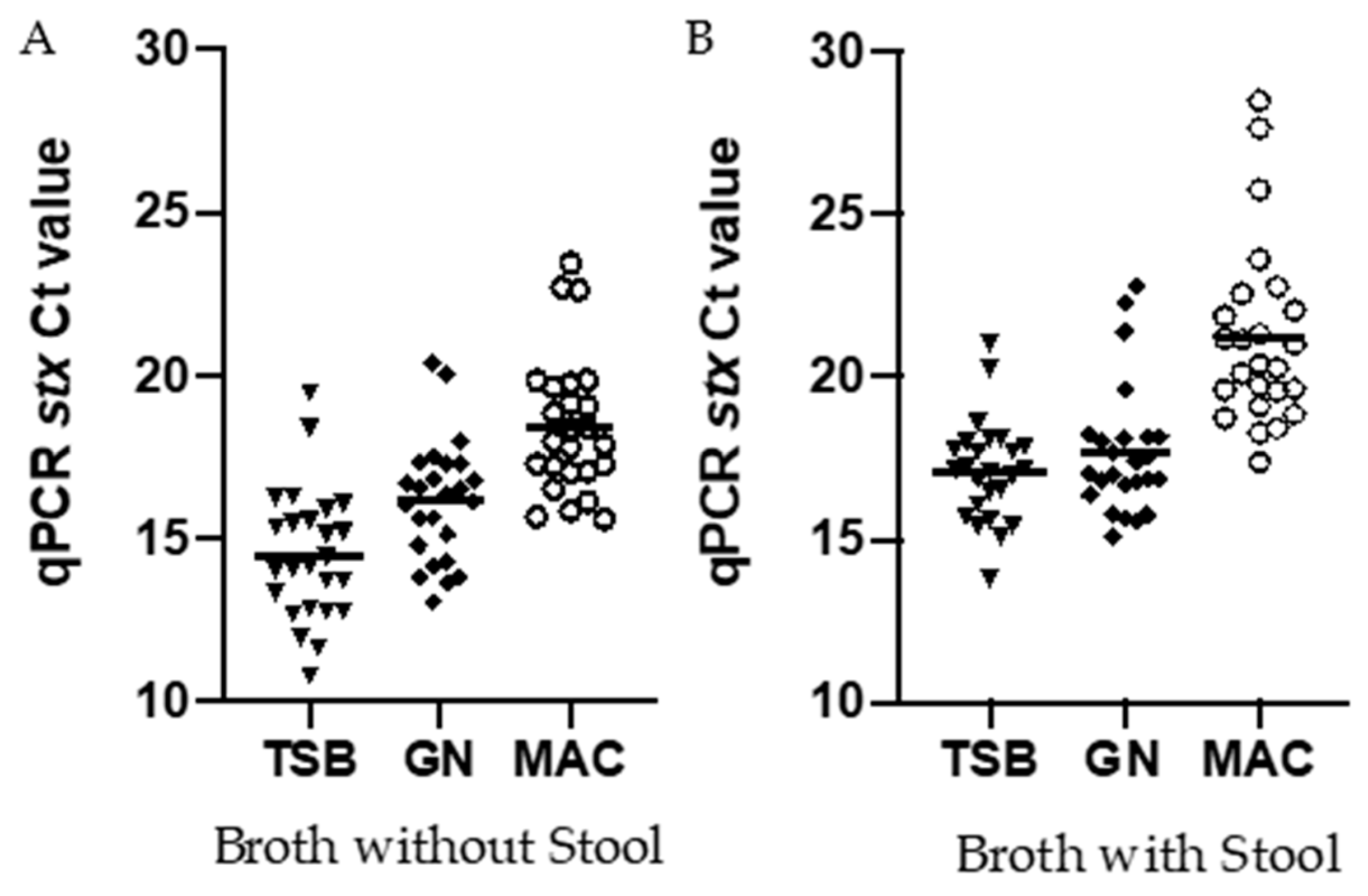
3.4. Comparing Ct Values Targeting the Stx Gene for Isolates Inoculated into Broth with and without Stool
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thomas, M.K.; Murray, R.; Flockhart, L.; Pintar, K.; Fazil, A.; Nesbitt, A.; Marshall, B.; Tataryn, J.; Pollari, F. Estimates of foodborne illness-related hospitalizations and deaths in Canada for 30 specified pathogens and unspecified agents. Foodborne Pathog. Dis. 2015, 12, 820–827. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.; Lefevre, S.; Donguy, M.P.; Nisavanh, A.; Terpant, G.; Fougere, E.; Vaissiere, E.; Guinard, A.; Mailles, A.; de Valk, H.; et al. Outbreak of Shiga toxin-producing Escherichia coli (STEC) O26 paediatric haemolytic uraemic syndrome (HUS) cases associated with the consumption of soft raw cow’s milk cheeses, France, March to May 2019. Eur. Surveill 2019, 24, 1900305. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, C.; Byrne, L.; Vishram, B.; Sawyer, C.; Balasegaram, S.; Ahyow, L.; Johnson, S. Shiga toxin-producing Escherichia coli haemolytic uraemic syndrome (STEC-HUS): Diagnosis, surveillance and public-health management in England. J. Med. Microbiol. 2020, 69, 1034–1036. [Google Scholar] [CrossRef]
- Tarr, G.A.M.; Stokowski, T.; Shringi, S.; Tarr, P.I.; Freedman, S.B.; Oltean, H.N.; Rabinowitz, P.M.; Chui, L. Contribution and Interaction of Shiga Toxin Genes to Escherichia coli O157:H7 Virulence. Toxins 2019, 11, 607. [Google Scholar] [CrossRef] [PubMed]
- Menge, C. Molecular Biology of Escherichia Coli Shiga Toxins’ Effects on Mammalian Cells. Toxins 2020, 12, 345. [Google Scholar] [CrossRef] [PubMed]
- Soledad-Cadona, J.; Bustamante, A.V.; Gonzalez, J.; Mariel-Sanso, A. Pathogenicity Islands Distribution in Non-O157 Shiga Toxin-Producing Escherichia coli (STEC). Genes 2018, 9, 81. [Google Scholar] [CrossRef]
- Beutin, L.; Krause, G.; Zimmermann, S.; Kaulfuss, S.; Gleier, K. Characterization of Shiga toxin-producing Escherichia coli strains isolated from human patients in Germany over a 3-year period. J. Clin. Microbiol. 2004, 42, 1099–1108. [Google Scholar] [CrossRef]
- Gould, L.H.; Mody, R.K.; Ong, K.L.; Clogher, P.; Cronquist, A.B.; Garman, K.N.; Lathrop, S.; Medus, C.; Spina, N.L.; Webb, T.H.; et al. Increased recognition of non-O157 Shiga toxin-producing Escherichia coli infections in the United States during 2000-2010: Epidemiologic features and comparison with E. coli O157 infections. Foodborne Pathog. Dis. 2013, 10, 453–460. [Google Scholar] [CrossRef]
- Collis, R.M.; Biggs, P.J.; Midwinter, A.C.; Browne, A.S.; Wilkinson, D.A.; Irshad, H.; French, N.P.; Brightwell, G.; Cookson, A.L. Genomic epidemiology and carbon metabolism of Escherichia coli serogroup O145 reflect contrasting phylogenies. PLoS ONE 2020, 15, e0235066. [Google Scholar] [CrossRef]
- Zelyas, N.; Poon, A.; Patterson-Fortin, L.; Johnson, R.P.; Lee, W.; Chui, L. Assessment of commercial chromogenic solid media for the detection of non-O157 Shiga toxin-producing Escherichia coli (STEC). Diagn. Microbiol. Infect. Dis. 2016, 85, 302–308. [Google Scholar] [CrossRef]
- Gould, L.H.; Bopp, C.; Strockbine, N.; Atkinson, R.; Baselski, V.; Body, B.; Carey, R.; Crandall, C.; Hurd, S.; Kaplan, R.; et al. Centers for Disease, Prevention, Recommendations for diagnosis of shiga toxin--producing Escherichia coli infections by clinical laboratories. MMWR Recomm. Rep. 2009, 58, 1–14. [Google Scholar]
- Humphries, R.M.; Linscott, A.J. Practical Guidance for Clinical Microbiology Laboratories: Diagnosis of Bacterial Gastroenteritis. Clin. Microbiol. Rev. 2015, 28, 3–31. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, C.; Perry, N.T.; Godbole, G.; Gharbia, S. Evaluation of chromogenic selective agar (CHROMagar STEC) for the direct detection of Shiga toxin-producing Escherichia coli from faecal specimens. J. Med. Microbiol. 2020, 69, 487–491. [Google Scholar] [CrossRef]
- Gouali, M.; Ruckly, C.; Carle, I.; Lejay-Collin, M.; Weill, F.X. Evaluation of CHROMagar STEC and STEC O104 chromogenic agar media for detection of Shiga Toxin-producing Escherichia coli in stool specimens. J. Clin. Microbiol. 2013, 51, 894–900. [Google Scholar] [CrossRef] [PubMed]
- Perelle, S.; Dilasser, F.; Grout, J.; Fach, P. Detection by 5’-nuclease PCR of Shiga-toxin producing Escherichia coli O26, O55, O91, O103, O111, O113, O145 and O157:H7, associated with the world’s most frequent clinical cases. Mol. Cell Probes. 2004, 18, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Morton, V.; Cheng, J.M.; Sharma, D.; Kearney, A. An outbreak of Shiga toxin-producing Escherichia coli O121 infections associated with flour-Canada 2016-2017(dagger). Can. Commun. Dis. Rep. 2017, 43, 154–155. [Google Scholar] [CrossRef] [PubMed]
- Brooks, J.T.; Sowers, E.G.; Wells, J.G.; Greene, K.D.; Griffin, P.M.; Hoekstra, R.M.; Strockbine, N.A. Non-O157 Shiga toxin-producing Escherichia coli infections in the United States 1983-2002. J. Infect. Dis. 2005, 192, 1422–1429. [Google Scholar] [CrossRef] [PubMed]
- Hussein, H.S.; Bollinger, L.M.; Hall, M.R. Growth and enrichment medium for detection and isolation of Shiga toxin-producing Escherichia coli in cattle feces. J. Food Prot. 2008, 71, 927–933. [Google Scholar] [CrossRef]
- Kanki, M.; Seto, K.; Harada, T.; Yonogi, S.; Kumeda, Y. Comparison of four enrichment broths for the detection of non-O157 Shiga-toxin-producing Escherichia coli O91, O103, O111, O119, O121, O145 and O165 from pure culture and food samples. Lett. Appl. Microbiol. 2011, 53, 167–173. [Google Scholar] [CrossRef]
- Dallman, T.; Cross, L.; Bishop, C.; Perry, N.; Olesen, B.; Grant, K.A.; Jenkins, C. Whole genome sequencing of an unusual serotype of Shiga toxin-producing Escherichia coli. Emerg. Infect. Dis. 2013, 19, 1302–1304. [Google Scholar] [CrossRef]
- Berger, P.; Kouzel, I.U.; Berger, M.; Haarmann, N.; Dobrindt, U.; Koudelka, G.B.; Mellmann, A. Carriage of Shiga toxin phage profoundly affects Escherichia coli gene expression and carbon source utilization. BMC Genom. 2019, 20, 504. [Google Scholar] [CrossRef] [PubMed]
- Martinson, J.N.V.; Pinkham, N.V.; Peters, G.W.; Cho, H.; Heng, J.; Rauch, M.; Broadaway, S.C.; Walk, S.T. Rethinking gut microbiome residency and the Enterobacteriaceae in healthy human adults. ISME J. 2019, 13, 2306–2318. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference Gene, Primer/Probe | Sequence 5′-3′ |
|---|---|
| stx1-F | TTT GTY ACT GTS ACA GCW GAA GCY TTA CG |
| stx1-R | CCC CAG TTC ARW GTR AGR TCM ACR TC |
| stx1-P | CTG GAT GAT CTC AGT GGG CGT TCT TAT GTA A |
| stx2-F | TTT GTY ACT GTS ACA GCW GAA GCY TTA CG |
| stx2-R | CCC CAG TTC ARW GTR AGR TCM ACR TC |
| stx2 -P | TCG TCA GGC ACT GTC TGA AAC TGC TCC |
| In the sequences: Y is (C, T), S is (C, G), W is (A, T), R is (A, G), M is (A, C). | |
| Broth | TSB | GN | Mac |
|---|---|---|---|
| TSB | NA | 38 (71%) | 42 (81%) |
| GN | 2 (3%) | NA | 28 (54%) |
| Mac | 0 | 0 | NA |
| No Significance | 6 (12%) | ||
| Broth | TSB | GN | Mac |
|---|---|---|---|
| TSB | NA | 23 (92%) | 25 (100%) |
| GN | 0 | NA | 8 (32%) |
| Mac | 0 | 0 | NA |
| No Significance | 0 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bording-Jorgensen, M.; Tyrrell, H.; Lloyd, C.; Chui, L. Comparison of Common Enrichment Broths Used in Diagnostic Laboratories for Shiga Toxin—Producing Escherichia coli. Microorganisms 2021, 9, 503. https://doi.org/10.3390/microorganisms9030503
Bording-Jorgensen M, Tyrrell H, Lloyd C, Chui L. Comparison of Common Enrichment Broths Used in Diagnostic Laboratories for Shiga Toxin—Producing Escherichia coli. Microorganisms. 2021; 9(3):503. https://doi.org/10.3390/microorganisms9030503
Chicago/Turabian StyleBording-Jorgensen, Michael, Hannah Tyrrell, Colin Lloyd, and Linda Chui. 2021. "Comparison of Common Enrichment Broths Used in Diagnostic Laboratories for Shiga Toxin—Producing Escherichia coli" Microorganisms 9, no. 3: 503. https://doi.org/10.3390/microorganisms9030503
APA StyleBording-Jorgensen, M., Tyrrell, H., Lloyd, C., & Chui, L. (2021). Comparison of Common Enrichment Broths Used in Diagnostic Laboratories for Shiga Toxin—Producing Escherichia coli. Microorganisms, 9(3), 503. https://doi.org/10.3390/microorganisms9030503