
Interconnections between the Oral and Gut Microbiomes: Reversal of Microbial Dysbiosis and the Balance between Systemic Health and Disease




 ,
,  ,
, 


Abstract
1. Introduction
2. Composition of Oral Microbiome and Its Impact on Health and Disease
3. Microbial Dysbiosis and Development of Oral and Systemic Diseases
4. Gut Microbiome and Its Impact on Systemic Health and Disease
5. Microbial Dysbiosis in the Gastrointestinal System and Metabolic Triggers of Systemic Diseases
6. Microbial Dysbiosis in Nutritional and Gastrointestinal Disorders and Cancers
7. The Link between the Oral and Gut Microbiomes

{kind=link}
{kind=link}
| GI Health Status | GI Microbiota | Importance | Ref. |
|---|---|---|---|
| Health | Firmicutes, Bacteroidetes, Actinobacteria, Proteobacteria | A typical healthy GI microbiome contains 30–50% Firmicutes, 20–40% Bacteroidetes, 1–10% Actinobacteria, and a small percent of Proteobacteria. | [18] |
| Bacteroides spp., Parabacteroides spp., Prevotella spp., Desulfovibrio spp., Ruminococcus spp., Akkermansia spp. | Three enterotypes have been discovered depending on diet: enterotype I (Bacteroides spp., Parabacteroides spp.) with carbohydrate, fat, and protein-based diets, enterotype II (Prevotella spp., Desulfovibrio spp.) with high-fiber diets, and enterotype III (Ruminococcus spp., Akkermansia spp.) which is the most commonly observed. | [24] | |
| Dysbiosis | Enterobacteriaceae | These bacteria are commonly associated with oxidative stress within the gut and relative increases in the proportion of these microbes can be found in individuals with inflammatory bowel disease, especially colitis | [23] |
| Bacteroides spp. | In individuals with low-gene richness, Bacteroides spp. increase non-proportionally and have the genomic potential to secrete metabolites that negatively impact the host. | [25] | |
| Obesity | Bacteroides spp. | The dominance of Bacteroides spp. in enterotype 1 leads to low-gene richness within the microbiota, which in-turn correlates with obesity, increased inflammation, and significantly higher levels of weight gain. | [25] |
| Diabetes | Escherichia coli, Clostridium hathewayi, Clostridium ramosum, Clostridium symbiosum | These are examples of opportunistic pathogens which are linked to bacteremia and intra-abdominal infections as a result of diabetic dysbiosis. | [28] |
| Inflammatory Bowel Diseases | Escherichia coli (AIEC), Ruminococcus torques, Ruminococcus gnavus, Enterobacteriaceae, Desulfovibrio spp. | In patients with IBD, these species have been shown to promote inflammation and increase mucus degradation, damaging epithelial cells and increasing intestinal permeability. AIEC has been shown to be present in 38% of individuals with active Crohn’s disease compared to 6% in healthy control subjects. | [23] |
| GI Cancer | Streptococcus bovis, Helicobacter pylori, Bacteroides fragilis, Enterococcus Faecalis, Fusobacterium spp., Escherichia coli (AIEC) | These species are suspected to aid in colorectal oncogenesis with relative risks of colorectal cancer and prevalence of other diseases (endocarditis, gastric cancer, periodontitis) increasing as well. | [18] |
| Fusobacterium nucleatum | Fusobacterium nucleatum has demonstrated an ability to colonize the GI tract and further promote microbial dysbiosis and subsequently colorectal cancer. Identical clones of oral Fusobacterium nucleatum have been isolated from the colorectal cancer lesions of patients with this disease. | [47] |
8. Newly Emerging Connections between Immune-Mediated Inflammatory Disease and Microbial Dysbiosis
9. Microbial Dysbiosis in the Oral Cavity Leading to Dysbiosis in the Gastrointestinal System
10. Therapeutics for Reversing Microbial Dysbiosis—Fecal Microbiota Transplantation
11. Therapeutics for Reversing Microbial Dysbiosis—Probiotics, Prebiotics, and Synbiotics
12. Targeted Therapy for Dysbiosis in Systemic Diseases
13. Targeted Therapy for Dysbiosis in Oral Diseases
14. Concluding Remarks, Limitations and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Iebba, V.; Totino, V.; Gagliardi, A.; Santangelo, F.; Cacciotti, F.; Trancassini, M.; Mancini, C.; Cicerone, C.; Corazziari, E.; Pantanella, F.; et al. Eubiosis and dysbiosis: The two sides of the microbiota. New Microbiol. 2016, 39, 1–12. [Google Scholar]
- Hillman, E.T.; Lu, H.; Yao, T.; Nakatsu, C.H. Microbial Ecology along the Gastrointestinal Tract. Microbes Environ. 2017, 32, 300–313. [Google Scholar] [CrossRef] [PubMed]
- Sampaio-Maia, B.; Caldas, I.; Pereira, M.; Pérez-Mongiovi, D.; Araujo, R. The Oral Microbiome in Health and Its Implication in Oral and Systemic Diseases. Adv. Appl. Microbiol. 2016, 97, 171–210. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, K.; Chen, T.; Paster, B.J. A practical guide to the oral microbiome and its relation to health and disease. Oral Dis. 2017, 23, 276–286. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.-X.; Hu, Y.-J.; Gao, L.; He, Z.-Y.; Zhu, C.-L.; Ma, R.; Huang, Z.-W. The Impact of Various Time Intervals on the Supragingival Plaque Dynamic Core Microbiome. PLoS ONE 2015, 10, e0124631. [Google Scholar] [CrossRef] [PubMed]
- Zaura, E.; Keijser, B.J.F.; Huse, S.M.; Crielaard, W. Defining the healthy “core microbiome” of oral microbial communities. BMC Microbiol. 2009, 9, 259. [Google Scholar] [CrossRef] [PubMed]
- Chimenos-Küstner, E.; Giovannoni, M.L.; Schemel-Suárez, M. Disbiosis como factor determinante de enfermedad oral y sistémica: Importancia del microbioma. Med. Clínica 2017, 149, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Aas, J.A.; Griffen, A.L.; Dardis, S.R.; Lee, A.M.; Olsen, I.; Dewhirst, F.E.; Leys, E.J.; Paster, B.J. Bacteria of Dental Caries in Primary and Permanent Teeth in Children and Young Adults. J. Clin. Microbiol. 2008, 46, 1407–1417. [Google Scholar] [CrossRef]
- Tanner, A.C.R.; Mathney, J.M.J.; Kent, R.L.; Chalmers, N.I.; Hughes, C.V.; Loo, C.Y.; Pradhan, N.; Kanasi, E.; Hwang, J.; Dahlan, M.A.; et al. Cultivable Anaerobic Microbiota of Severe Early Childhood Caries. J. Clin. Microbiol. 2011, 49, 1464–1474. [Google Scholar] [CrossRef]
- Manji, F.; Dahlén, G.; Fejerskov, O. Caries and Periodontitis: Contesting the Conventional Wisdom on Their Aetiology. Caries Res. 2018, 52, 548–564. [Google Scholar] [CrossRef]
- Socransky, S.S.; Haffajee, A.D.; Cugini, M.A.; Smith, C.; Kent, R.L., Jr. Microbial complexes in subgingival plaque. J. Clin. Periodontol. 1998, 25, 134–144. [Google Scholar] [CrossRef]
- Siqueira, J.; Rôças, I. Diversity of Endodontic Microbiota Revisited. J. Dent. Res. 2009, 88, 969–981. [Google Scholar] [CrossRef] [PubMed]
- Jia, G.; Zhi, A.; Lai, P.F.H.; Wang, G.; Xia, Y.; Xiong, Z.; Zhang, H.; Che, N.; Ai, L. The oral microbiota—A mechanistic role for systemic diseases. Br. Dent. J. 2018, 224, 447–455. [Google Scholar] [CrossRef]
- Chhibber-Goel, J.; Singhal, V.; Bhowmik, D.; Vivek, R.; Parakh, N.; Bhargava, B.; Sharma, A. Linkages between oral commensal bacteria and atherosclerotic plaques in coronary artery disease patients. NPJ Biofilms Microbiomes 2016, 2, 7. [Google Scholar] [CrossRef] [PubMed]
- Scannapieco, F.A.; Mylotte, J.M. Relationships Between Periodontal Disease and Bacterial Pneumonia. J. Periodontol. 1996, 67, 1114–1122. [Google Scholar] [CrossRef]
- Bansal, M.; Khatri, M.; Taneja, V. Potential role of periodontal infection in respiratory diseases—A review. J. Med. Life 2013, 6, 244–248. [Google Scholar] [PubMed]
- Thursby, E.; Juge, N. Introduction to the human gut microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef] [PubMed]
- Gagnière, J.; Raisch, J.; Veziant, J.; Barnich, N.; Bonnet, R.; Buc, E.; Bringer, M.-A.; Pezet, D.; Bonnet, M. Gut microbiota imbalance and colorectal cancer. World J. Gastroenterol. 2016, 22, 501–518. [Google Scholar] [CrossRef] [PubMed]
- Weiss, G.A.; Hennet, T. Mechanisms and consequences of intestinal dysbiosis. Cell. Mol. Life Sci. 2017, 74, 2959–2977. [Google Scholar] [CrossRef]
- Giuffrè, M.; Campigotto, M.; Campisciano, G.; Comar, M.; Crocè, L.S. A story of liver and gut microbes: How does the intestinal flora affect liver disease? A review of the literature. Am. J. Physiol. Liver Physiol. 2020, 318, G889–G906. [Google Scholar] [CrossRef]
- Giuffrè, M.; Moretti, R.; Campisciano, G.; Da Silveira, A.B.M.; Monda, V.M.; Comar, M.; Di Bella, S.; Antonello, R.M.; Luzzati, R.; Crocè, L.S. You Talking to Me? Says the Enteric Nervous System (ENS) to the Microbe. How Intestinal Microbes Interact with the ENS. J. Clin. Med. 2020, 9, 3705. [Google Scholar] [CrossRef]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.M.; et al. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Nishida, A.; Inoue, R.; Inatomi, O.; Bamba, S.; Naito, Y.; Andoh, A. Gut microbiota in the pathogenesis of inflammatory bowel disease. Clin. J. Gastroenterol. 2018, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Malard, F.; Dore, J.; Gaugler, B.; Mohty, M. Introduction to host microbiome symbiosis in health and disease. Mucosal Immunol. 2020, 1–8. [Google Scholar] [CrossRef]
- Le Chatelier, E.; Nielsen, T.; Qin, J.; Prifti, E.; Hildebrand, F.; Falony, G.; Almeida, M.; Arumugam, M.; Batto, J.-M.; Kennedy, S.; et al. Richness of human gut microbiome correlates with metabolic markers. Nature 2013, 500, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Cotillard, A.; Kennedy, S.P.; Kong, L.C.; Prifti, E.; Pons, N.; Le Chatelier, E.; Almeida, M.; Quinquis, B.; Levenez, F.; Galleron, N.; et al. Dietary intervention impact on gut microbial gene richness. Nature 2013, 500, 585–588. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.; Bibiloni, R.; Knauf, C.; Waget, A.; Neyrinck, A.M.; Delzenne, N.M.; Burcelin, R. Changes in Gut Microbiota Control Metabolic Endotoxemia-Induced Inflammation in High-Fat Diet-Induced Obesity and Diabetes in Mice. Diabetes 2008, 57, 1470–1481. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, Y.; Cai, Z.; Li, S.; Zhu, J.; Zhang, F.; Liang, S.; Zhang, W.; Guan, Y.; Shen, D.; et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 2012, 490, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Wu, G.D.; Albenberg, L.; Tomov, V.T. Gut microbiota and IBD: Causation or correlation? Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 573–584. [Google Scholar] [CrossRef]
- Atarashi, K.; Tanoue, T.; Oshima, K.; Suda, W.; Nagano, Y.; Nishikawa, H.; Fukuda, S.; Saito, T.; Narushima, S.; Hase, K.; et al. Treg induction by a rationally selected mixture of Clostridia strains from the human microbiota. Nature 2013, 500, 232–236. [Google Scholar] [CrossRef]
- Ahmed, I.; Roy, B.C.; Khan, S.A.; Septer, S.; Umar, S. Microbiome, Metabolome and Inflammatory Bowel Disease. Microorganisms 2016, 4, 20. [Google Scholar] [CrossRef] [PubMed]
- Couturier-Maillard, A.; Secher, T.; Rehman, A.; Normand, S.; De Arcangelis, A.; Häesler, R.; Huot, L.; Grandjean, T.; Bressenot, A.; Delanoye-Crespin, A.; et al. NOD2-mediated dysbiosis predisposes mice to transmissible colitis and colorectal cancer. J. Clin. Investig. 2013, 123, 700–711. [Google Scholar] [CrossRef]
- McCoy, W.C.; Mason, J.M. Enterococcal endocarditis associated with carcinoma of the sigmoid; report of a case. J. Med. Assoc. State Ala. 1951, 21, 162–166. [Google Scholar] [PubMed]
- Abdulamir, A.S.; Hafidh, R.R.; Abu Bakar, F. Molecular detection, quantification, and isolation of Streptococcus gallolyticus bacteria colonizing colorectal tumors: Inflammation-driven potential of carcinogenesis via IL-1, COX-2, and IL-8. Mol. Cancer 2010, 9, 249. [Google Scholar] [CrossRef]
- Guo, Y.; Li, H.-Y. Association between Helicobacter pylori infection and colorectal neoplasm risk: A meta-analysis Based on East Asian population. J. Cancer Res. Ther. 2014, 10, 263–266. [Google Scholar] [CrossRef] [PubMed]
- Zumkeller, N.; Brenner, H.; Zwahlen, M.; Rothenbacher, D. Helicobacter pylori Infection and Colorectal Cancer Risk: A Meta-Analysis. Helicobacter 2006, 11, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Cai, G.; Qiu, Y.; Fei, N.; Zhang, M.; Pang, X.; Jia, W.; Cai, S.; Zhao, L. Structural segregation of gut microbiota between colorectal cancer patients and healthy volunteers. ISME J. 2011, 6, 320–329. [Google Scholar] [CrossRef]
- Martin, H.M.; Campbell, B.J.; Hart, C.; Mpofu, C.; Nayar, M.; Singh, R.; Englyst, H.; Williams, H.F.; Rhodes, J.M. Enhanced Escherichia coli adherence and invasion in Crohn’s disease and colon cancer 1. Gastroenterology 2004, 127, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Arthur, J.C.; Perez-Chanona, E.; Mühlbauer, M.; Tomkovich, S.; Uronis, J.M.; Fan, T.-J.; Campbell, B.J.; Abujamel, T.; Dogan, B.; Rogers, A.B.; et al. Intestinal Inflammation Targets Cancer-Inducing Activity of the Microbiota. Science 2012, 338, 120–123. [Google Scholar] [CrossRef]
- Kostic, A.D.; Chun, E.; Robertson, L.; Glickman, J.N.; Gallini, C.A.; Michaud, M.; Clancy, T.E.; Chung, D.C.; Lochhead, P.; Hold, G.L.; et al. Fusobacterium nucleatum Potentiates Intestinal Tumorigenesis and Modulates the Tumor-Immune Microenvironment. Cell Host Microbe 2013, 14, 207–215. [Google Scholar] [CrossRef]
- Atarashi, K.; Suda, W.; Luo, C.; Kawaguchi, T.; Motoo, I.; Narushima, S.; Kiguchi, Y.; Yasuma, K.; Watanabe, E.; Tanoue, T.; et al. Ectopic colonization of oral bacteria in the intestine drives TH1 cell induction and inflammation. Science 2017, 358, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Kitamoto, S.; Nagao-Kitamoto, H.; Hein, R.; Schmidt, T.; Kamada, N. The Bacterial Connection between the Oral Cavity and the Gut Diseases. J. Dent. Res. 2020, 99, 1021–1029. [Google Scholar] [CrossRef] [PubMed]
- Walker, M.Y.; Pratap, S.; Southerland, J.H.; Farmer-Dixon, C.M.; Lakshmyya, K.; Gangula, P.R. Role of oral and gut microbiome in nitric oxide-mediated colon motility. Nitric Oxide 2018, 73, 81–88. [Google Scholar] [CrossRef]
- Schmidt, T.S.; Hayward, M.R.; Coelho, L.P.; Li, S.S.; Costea, P.I.; Voigt, A.Y.; Wirbel, J.; Maistrenko, O.M.; Alves, R.J.; Bergsten, E.; et al. Extensive transmission of microbes along the gastrointestinal tract. eLife 2019, 8, 8. [Google Scholar] [CrossRef]
- Qin, N.; Yang, F.; Li, A.; Prifti, E.; Chen, Y.; Shao, L.; Guo, J.; Le Chatelier, E.; Yao, J.; Wu, L.; et al. Alterations of the human gut microbiome in liver cirrhosis. Nature 2014, 513, 59–64. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, D.; Jia, H.; Feng, Q.; Wang, D.; Liang, D.; Wu, X.; Li, J.; Tang, L.; Li, Y.; et al. The oral and gut microbiomes are perturbed in rheumatoid arthritis and partly normalized after treatment. Nat. Med. 2015, 21, 895–905. [Google Scholar] [CrossRef]
- Komiya, Y.; Shimomura, Y.; Higurashi, T.; Sugi, Y.; Arimoto, J.; Umezawa, S.; Uchiyama, S.; Matsumoto, M.; Nakajima, A. Patients with colorectal cancer have identical strains of Fusobacterium nucleatum in their colorectal cancer and oral cavity. Gut 2018, 68, 1335–1337. [Google Scholar] [CrossRef] [PubMed]
- Van Der Meulen, T.A.; Harmsen, H.J.M.; Bootsma, H.; Spijkervet, F.K.L.; Kroese, F.G.M.; Vissink, A. The microbiome-systemic diseases connection. Oral Dis. 2016, 22, 719–734. [Google Scholar] [CrossRef] [PubMed]
- Corrêa, J.D.; Calderaro, D.C.; Ferreira, G.A.; Mendonça, S.M.S.; Fernandes, G.R.; Xiao, E.; Teixeira, A.L.; Leys, E.J.; Graves, D.T.; Silva, T.A. Subgingival microbiota dysbiosis in systemic Lupus erythematosus: Association with periodontal status. Microbiome 2017, 5, 1–13. [Google Scholar] [CrossRef]
- Nikitakis, N.G.; Papaioannou, W.; I Sakkas, L.; Kousvelari, E. The autoimmunity-oral microbiome connection. Oral Dis. 2016, 23, 828–839. [Google Scholar] [CrossRef] [PubMed]
- Acharya, C.; Sahingur, S.E.; Bajaj, J.S. Microbiota, cirrhosis, and the emerging oral-gut-liver axis. JCI Insight 2017, 2. [Google Scholar] [CrossRef]
- Hevia, A.; Milani, C.; López, P.; Cuervo, A.; Arboleya, S.; Duranti, S.; Turroni, F.; González, S.; Suárez, A.; Gueimonde, M.; et al. Intestinal Dysbiosis Associated with Systemic Lupus Erythematosus. mBio 2014, 5, e01548-14. [Google Scholar] [CrossRef] [PubMed]
- Van der Meulen, T.A.; Harmsen, H.J.; Vila, A.V.; Kurilshikov, A.; Liefers, S.C.; Zhernakova, A.; Fu, J.; Wijmenga, C.; Weersma, R.K.; de Leeuw, K.; et al. Shared gut, but distinct oral microbiota composition in primary Sjögren’s syndrome and systemic lupus erythematosus. J. Autoimmun. 2019, 97, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Comstock, L.E. Importance of Glycans to the Host-Bacteroides Mutualism in the Mammalian Intestine. Cell Host Microbe 2009, 5, 522–526. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Sandhya, P.; Vellarikkal, S.K.; Surin, A.K.; Jayarajan, R.; Verma, A.; Kumar, A.; Ravi, R.; Danda, D.; Sivasubbu, S.; et al. Saliva microbiome in primary Sjögren’s syndrome reveals distinct set of disease-associated microbes. Oral Dis. 2019, 26, 295–301. [Google Scholar] [CrossRef]
- Singh, M.; Teles, F.; Uzel, N.; Papas, A. Characterizing Microbiota from Sjögren’s Syndrome Patients. JDR Clin. Transl. Res. 2020. [Google Scholar] [CrossRef]
- Imai, K.; Inoue, H.; Tamura, M.; Cueno, M.E.; Inoue, H.; Takeichi, O.; Kusama, K.; Saito, I.; Ochiai, K. The periodontal pathogen Porphyromonas gingivalis induces the Epstein–Barr virus lytic switch transactivator ZEBRA by histone modification. Biochimie 2012, 94, 839–846. [Google Scholar] [CrossRef]
- Croia, C.; Astorri, E.; Murray-Brown, W.; Willis, A.; Brokstad, K.A.; Sutcliffe, N.; Piper, K.; Jonsson, R.; Tappuni, A.R.; Pitzalis, C.; et al. Implication of Epstein-Barr Virus Infection in Disease-Specific Autoreactive B Cell Activation in Ectopic Lymphoid Structures of Sjögren’s Syndrome. Arthritis Rheumatol. 2014, 66, 2545–2557. [Google Scholar] [CrossRef] [PubMed]
- Maddi, A.; Scannapieco, F.A. Oral biofilms, oral and periodontal infections, and systemic disease. Am. J. Dent. 2013, 26, 249–254. [Google Scholar] [PubMed]
- Scher, J.U.; Sczesnak, A.; Longman, R.S.; Segata, N.; Ubeda, C.; Bielski, C.; Rostron, T.; Cerundolo, V.; Pamer, E.G.; Abramson, S.B.; et al. Expansion of intestinal Prevotella copri correlates with enhanced susceptibility to arthritis. eLife 2013, 2, e01202. [Google Scholar] [CrossRef] [PubMed]
- Flemer, B.; Warren, R.D.; Barrett, M.P.; Cisek, K.; Das, A.; Jeffery, I.B.; Hurley, E.; O‘Riordain, M.; Shanahan, F.; O’Toole, P.W. The oral microbiota in colorectal cancer is distinctive and predictive. Gut 2017, 67, 1454–1463. [Google Scholar] [CrossRef]
- Komazaki, R.; Katagiri, S.; Takahashi, H.; Maekawa, S.; Shiba, T.; Takeuchi, Y.; Kitajima, Y.; Ohtsu, A.; Udagawa, S.; Sasaki, N.; et al. Periodontal pathogenic bacteria, Aggregatibacter actinomycetemcomitans affect non-alcoholic fatty liver disease by altering gut microbiota and glucose metabolism. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef]
- Arimatsu, K.; Yamada, H.; Miyazawa, H.; Minagawa, T.; Nakajima, M.; Ryder, M.I.; Gotoh, K.; Motooka, D.; Nakamura, S.; Iida, T.; et al. Oral pathobiont induces systemic inflammation and metabolic changes associated with alteration of gut microbiota. Sci. Rep. 2015, 4, 4828. [Google Scholar] [CrossRef]
- Nakajima, M.; Arimatsu, K.; Kato, T.; Matsuda, Y.; Minagawa, T.; Takahashi, N.; Ohno, H.; Yamazaki, K. Oral Administration of P. gingivalis Induces Dysbiosis of Gut Microbiota and Impaired Barrier Function Leading to Dissemination of Enterobacteria to the Liver. PLoS ONE 2015, 10, e0134234. [Google Scholar] [CrossRef]
- Yoneda, M.; Naka, S.; Nakano, K.; Wada, K.; Endo, H.; Mawatari, H.; Imajo, K.; Nomura, R.; Hokamura, K.; Ono, M.; et al. Involvement of a periodontal pathogen, Porphyromonas gingivalis on the pathogenesis of non-alcoholic fatty liver disease. BMC Gastroenterol. 2012, 12, 16. [Google Scholar] [CrossRef] [PubMed]
- Gough, E.; Shaikh, H.; Manges, A.R. Systematic Review of Intestinal Microbiota Transplantation (Fecal Bacteriotherapy) for Recurrent Clostridium difficile Infection. Clin. Infect. Dis. 2011, 53, 994–1002. [Google Scholar] [CrossRef]
- Vrieze, A.; Van Nood, E.; Holleman, F.; Salojärvi, J.; Kootte, R.S.; Bartelsman, J.F.; Dallinga–Thie, G.M.; Ackermans, M.T.; Serlie, M.J.; Oozeer, R.; et al. Transfer of Intestinal Microbiota From Lean Donors Increases Insulin Sensitivity in Individuals With Metabolic Syndrome. Gastroenterology 2012, 143, 913–916.e7. [Google Scholar] [CrossRef]
- Kootte, R.S.; Levin, E.; Salojärvi, J.; Smits, L.P.; Hartstra, A.V.; Udayappan, S.D.; Hermes, G.; Bouter, K.E.; Koopen, A.M.; Holst, J.J.; et al. Improvement of Insulin Sensitivity after Lean Donor Feces in Metabolic Syndrome Is Driven by Baseline Intestinal Microbiota Composition. Cell Metab. 2017, 26, 611–619.e6. [Google Scholar] [CrossRef] [PubMed]
- Colman, R.J.; Rubin, D.T. Fecal microbiota transplantation as therapy for inflammatory bowel disease: A systematic review and meta-analysis. J. Crohns Colitis 2014, 8, 1569–1581. [Google Scholar] [CrossRef]
- Borody, T.; Leis, S.; Campbell, J.; Torres, M.; Nowak, A. Fecal Microbiota Transplantation [FMT] in Multiple Sclerosis [MS]: 942. Am. J. Gastroenterol. 2011, 106, S352. [Google Scholar] [CrossRef]
- Borody, T.; Campbell, J.; Torres, M.; Nowak, A.; Leis, S. Reversal of Idiopathic Thrombocytopenic Purpura [ITP] with Fecal Microbiota Transplantation [FMT]: 941. Am. J. Gastroenterol. 2011, 106, S352. [Google Scholar] [CrossRef]
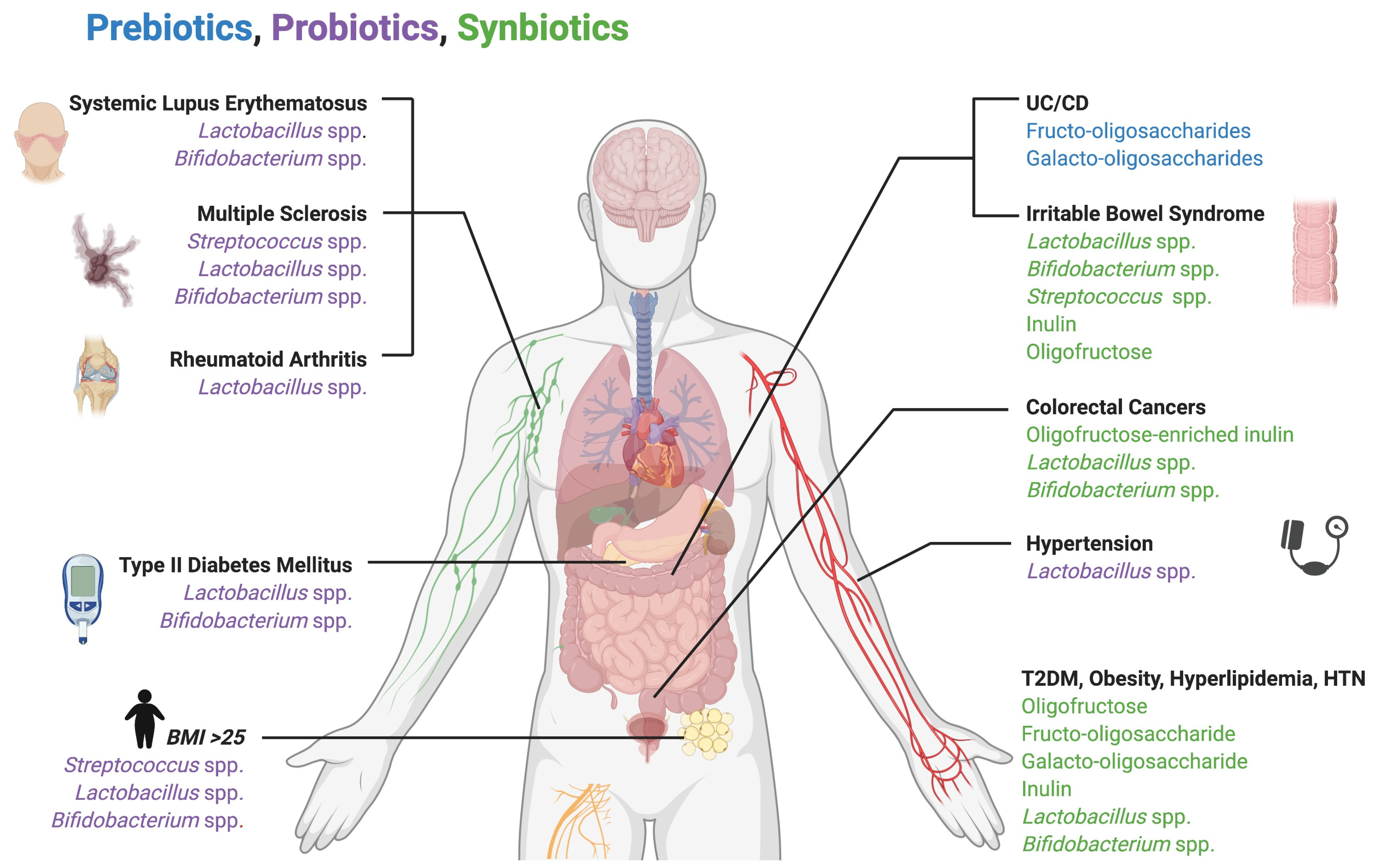
- Tonucci, L.B.; dos Santos, K.M.O.; de Oliveira, L.L.; Ribeiro, S.M.R.; Martino, H.S.D. Clinical application of probiotics in type 2 diabetes mellitus: A randomized, double-blind, placebo-controlled study. Clin. Nutr. 2017, 36, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Bose, S.; Seo, J.-G.; Chung, W.-S.; Lim, C.-Y.; Kim, H. The effects of co-administration of probiotics with herbal medicine on obesity, metabolic endotoxemia and dysbiosis: A randomized double-blind controlled clinical trial. Clin. Nutr. 2014, 33, 973–981. [Google Scholar] [CrossRef]
- Dong, J.-Y.; Szeto, I.M.Y.; Makinen, K.; Gao, Q.; Wang, J.; Qin, L.-Q.; Zhao, Y. Effect of probiotic fermented milk on blood pressure: A meta-analysis of randomised controlled trials. Br. J. Nutr. 2013, 110, 1188–1194. [Google Scholar] [CrossRef] [PubMed]
- Rajkumar, H.; Mahmood, N.; Kumar, M.; Varikuti, S.R.; Challa, H.R.; Myakala, S.P. Effect of Probiotic (VSL#3) and Omega-3 on Lipid Profile, Insulin Sensitivity, Inflammatory Markers, and Gut Colonization in Overweight Adults: A Randomized, Controlled Trial. Mediat. Inflamm. 2014, 2014, 1–8. [Google Scholar] [CrossRef]
- Kellow, N.J.; Coughlan, M.T.; Reid, C.M. Metabolic benefits of dietary prebiotics in human subjects: A systematic review of randomised controlled trials. Br. J. Nutr. 2014, 111, 1147–1161. [Google Scholar] [CrossRef]
- Lindsay, J.O.; Whelan, K.; Stagg, A.J.; Gobin, P.; Al-Hassi, H.O.; Rayment, N.; Kamm, M.A.; Knight, S.C.; Forbes, A. Clinical, microbiological, and immunological effects of fructo-oligosaccharide in patients with Crohn’s disease. Gut 2006, 55, 348–355. [Google Scholar] [CrossRef]
- Matijašić, B.B.; Obermajer, T.; Lipoglavšek, L.; Sernel, T.; Locatelli, I.; Kos, M.; Šmid, A.; Rogelj, I. Effects of synbiotic fermented milk containing Lactobacillus acidophilus La-5 and Bifidobacterium animalis ssp. lactis BB-12 on the fecal microbiota of adults with irritable bowel syndrome: A randomized double-blind, placebo-controlled trial. J. Dairy Sci. 2016, 99, 5008–5021. [Google Scholar] [CrossRef]
- Esmaeili, S.-A.; Mahmoudi, M.; Momtazi, A.A.; Sahebkar, A.; Doulabi, H.; Rastin, M. Tolerogenic probiotics: Potential immunoregulators in Systemic Lupus Erythematosus. J. Cell. Physiol. 2017, 232, 1994–2007. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, A.T.; Khattab, M.; Ahmed, A.M.; Turk, T.; Sakr, N.; Khalil, A.M.; Abdelhalim, M.; Sawaf, B.; Hirayama, K.; Huy, N.T. The therapeutic effect of probiotics on rheumatoid arthritis: A systematic review and meta-analysis of randomized control trials. Clin. Rheumatol. 2017, 36, 2697–2707. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Hoang, T.K.; Tian, X.; Taylor, C.M.; Blanchard, E.; Luo, M.; Bhattacharjee, M.B.; Freeborn, J.; Park, S.; Couturier, J.; et al. Lactobacillus reuteri Reduces the Severity of Experimental Autoimmune Encephalomyelitis in Mice by Modulating Gut Microbiota. Front. Immunol. 2019, 10, 385. [Google Scholar] [CrossRef] [PubMed]
- Tankou, S.K.; Regev, K.; Healy, B.C.; Tjon, E.; Laghi, L.; Cox, L.M.; Kivisäkk, P.; Pierre, I.V.; Hrishikesh, L.; Gandhi, R.; et al. A probiotic modulates the microbiome and immunity in multiple sclerosis. Ann. Neurol. 2018, 83, 1147–1161. [Google Scholar] [CrossRef] [PubMed]
- Rafter, J.; Bennett, M.; Caderni, G.; Clune, Y.; Hughes, R.; Karlsson, P.C.; Klinder, A.; O’Riordan, M.; O’Sullivan, G.C.; Pool-Zobel, B.; et al. Dietary synbiotics reduce cancer risk factors in polypectomized and colon cancer patients. Am. J. Clin. Nutr. 2007, 85, 488–496. [Google Scholar] [CrossRef]
- Agah, S.; Alizadeh, A.M.; Mosavi, M.; Ranji, P.; Khavari-Daneshvar, H.; Ghasemian, F.; Bahmani, S.; Tavassoli, A. More Protection of Lactobacillus acidophilus Than Bifidobacterium bifidum Probiotics on Azoxymethane-Induced Mouse Colon Cancer. Probiotics Antimicrob. Proteins 2018, 11, 857–864. [Google Scholar] [CrossRef]
- Munjal, U.; Glei, M.; Pool-Zobel, B.L.; Scharlau, D. Fermentation products of inulin-type fructans reduce proliferation and induce apoptosis in human colon tumour cells of different stages of carcinogenesis. Br. J. Nutr. 2009, 102, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Winter, M.G.; Byndloss, M.X.; Spiga, L.; Duerkop, B.A.; Hughes, E.R.; Büttner, L.; Romão, E.D.L.; Behrendt, C.L.; Lopez, C.A.; et al. Precision editing of the gut microbiota ameliorates colitis. Nature 2018, 553, 208–211. [Google Scholar] [CrossRef]
- Hughes, E.R.; Winter, M.G.; Duerkop, B.A.; Spiga, L.; De Carvalho, T.F.; Zhu, W.; Gillis, C.C.; Büttner, L.; Smoot, M.P.; Behrendt, C.L.; et al. Microbial Respiration and Formate Oxidation as Metabolic Signatures of Inflammation-Associated Dysbiosis. Cell Host Microbe 2017, 21, 208–219. [Google Scholar] [CrossRef] [PubMed]
- Gates, A.J.; Hughes, R.O.; Sharp, S.R.; Millington, P.D.; Nilavongse, A.; Cole, J.A.; Leach, E.-R.; Jepson, B.; Richardson, D.J.; Butler, C.S. Properties of the periplasmic nitrate reductases from Paracoccus pantotrophus and Escherichia coli after growth in tungsten-supplemented media. FEMS Microbiol. Lett. 2003, 220, 261–269. [Google Scholar] [CrossRef]
- Sullivan, E.L.; Nousen, E.K.; Chamlou, K.A. Maternal high fat diet consumption during the perinatal period programs offspring behavior. Physiol. Behav. 2014, 123, 236–242. [Google Scholar] [CrossRef]
- Ma, J.; Prince, A.L.; Bader, D.; Hu, M.; Ganu, R.; Baquero, K.; Blundell, P.; Harris, R.A.; Frias, A.E.; Grove, K.L.; et al. High-fat maternal diet during pregnancy persistently alters the offspring microbiome in a primate model. Nat. Commun. 2014, 5, 1–11. [Google Scholar] [CrossRef]
- Buffington, S.A.; Di Prisco, G.V.; Auchtung, T.A.; Ajami, N.J.; Petrosino, J.F.; Costa-Mattioli, M. Microbial Reconstitution Reverses Maternal Diet-Induced Social and Synaptic Deficits in Offspring. Cell 2016, 165, 1762–1775. [Google Scholar] [CrossRef] [PubMed]
- Zaura, E.; Brandt, B.W.; Prodan, A.; De Mattos, M.J.T.; Imangaliyev, S.; Kool, J.; Buijs, M.J.; Jagers, F.L.; Hennequin-Hoenderdos, N.L.; E Slot, D.; et al. On the ecosystemic network of saliva in healthy young adults. ISME J. 2017, 11, 1218–1231. [Google Scholar] [CrossRef]
- Belibasakis, G.N.; Bostanci, N.; Marsh, P.D.; Zaura, E. Applications of the oral microbiome in personalized dentistry. Arch. Oral Biol. 2019, 104, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Wen, S.; Yang, L.; Zheng, G. Mucosal anti-caries DNA vaccine: A new approach to induce protective immunity against streptococcus mutans. Int. J. Exp. Pathol. 2017, 10, 853–857. [Google Scholar]
- Robinette, R.A.; Oli, M.W.; McArthur, W.P.; Brady, L.J. A therapeutic anti-Streptococcus mutans monoclonal antibody used in human passive protection trials influences the adaptive immune response. Vaccine 2011, 29, 6292–6300. [Google Scholar] [CrossRef][Green Version]
- Smith, D.J. Dental caries vaccines: Prospects and concerns. Expert Rev. Vaccines 2010, 9, 1–3. [Google Scholar] [CrossRef] [PubMed]
- López-López, A.; Camelo-Castillo, A.; Ferrer, M.D.; Simon-Soro, Á.; Mira, A. Health-Associated Niche Inhabitants as Oral Probiotics: The Case of Streptococcus dentisani. Front. Microbiol. 2017, 8, 379. [Google Scholar] [CrossRef]
- Liu, Y.-L.; Nascimento, M.; Burne, R.A. Progress toward understanding the contribution of alkali generation in dental biofilms to inhibition of dental caries. Int. J. Oral Sci. 2012, 4, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, L.; Zhou, X.; Hu, S.; Zhang, S.; Wu, H. Effect of the antimicrobial decapeptide KSL on the growth of oral pathogens and Streptococcus mutans biofilm. Int. J. Antimicrob. Agents 2011, 37, 33–38. [Google Scholar] [CrossRef]
- Weistroffer, P.L.; Joly, S.; Srikantha, R.; Tack, B.F.; Brogden, K.A.; Guthmiller, J.M. SMAP29 congeners demonstrate activity against oral bacteria and reduced toxicity against oral keratinocytes. Oral Microbiol. Immunol. 2008, 23, 89–95. [Google Scholar] [CrossRef]
- Franzman, M.R.; Burnell, K.K.; Dehkordi-Vakil, F.H.; Guthmiller, J.M.; Dawson, D.V.; Brogden, K.A. Targeted antimicrobial activity of a specific IgG–SMAP28 conjugate against Porphyromonas gingivalis in a mixed culture. Int. J. Antimicrob. Agents 2009, 33, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Ebersole, J.L.; Kirakodu, S.S.; Gonzalez, O.A. Oral microbiome interactions with gingival gene expression patterns for apoptosis, autophagy and hypoxia pathways in progressing periodontitis. Immunology 2020. [Google Scholar] [CrossRef]
- Vujkovic-Cvijin, I.; Sklar, J.; Jiang, L.; Natarajan, L.; Knight, R.; Belkaid, Y. Host variables confound gut microbiota studies of human disease. Nat. Cell Biol. 2020, 587, 448–454. [Google Scholar] [CrossRef]
- Mimee, M.; Citorik, R.J.; Lu, T.K. Microbiome therapeutics—Advances and challenges. Adv. Drug Deliv. Rev. 2016, 105, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Kumar, T.B.; Thotakura, P.L.; Tiwary, B.K.; Krishna, R. Target identification in Fusobacterium nucleatum by subtractive genomics approach and enrichment analysis of host-pathogen protein-protein interactions. BMC Microbiol. 2016, 16, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ismail, T.; Fatima, N.; Muhammad, S.A.; Zaidi, S.S.; Rehman, N.; Hussain, I.; Tariq, N.U.S.; Amirzada, I.; Mannan, A. Prioritizing and modelling of putative drug target proteins of Candida albicans by system biology approach. Acta Biochim. Pol. 2018, 65, 209–218. [Google Scholar] [CrossRef]
- O’Toole, P.W.; Marchesi, J.R.; Hill, C. Next-generation probiotics: The spectrum from probiotics to live biotherapeutics. Nat. Microbiol. 2017, 2, 17057. [Google Scholar] [CrossRef]


| Oral Health Status | Oral Microbiota | Importance | Reference |
|---|---|---|---|
| Health | Streptococcus mitis, Streptococcus salivarius, Streptococcus sanguinis | Initially, various Streptococcus spp. are acquired as pioneer species and begin to modulate pH and nutrient availability in the oral cavity, setting the tone for subsequent colonization by other members of the oral microbiota. | [4] |
| Fusobacterium spp., Granulicatella spp., Neisseria spp., Haemophilus spp., Corynebacterium spp., Rothia spp., Actinomyces spp., Prevotella spp., Capnocytophaga spp., Porphyromonas spp. | These phylotypes are commonly associated with healthy oral microbiomes and monitoring relative numbers of these species can indicate a change in homeostatic balance. | [6] | |
| Caries | Streptococcus mutans | Streptococcus mutans can resist high levels of oxidative stress and in-turn has the ability to out-compete other microorganisms under conditions of high carbohydrate metabolism. These abilities change the homeostatic balance of the oral microbiome and allow Streptococcus mutans to be the primary driver of carious lesions. | [7] |
| Lactobacillus spp., Propionibacterium spp., Atopobium spp. Bifidobacterium dentium | The listed microbes thrive in low-pH conditions even when Streptococcus mutans is not present. Carious lesions lacking Streptococcus mutans reported higher levels of Bifidobacterium dentium and Lactobacilllus spp. | [8] | |
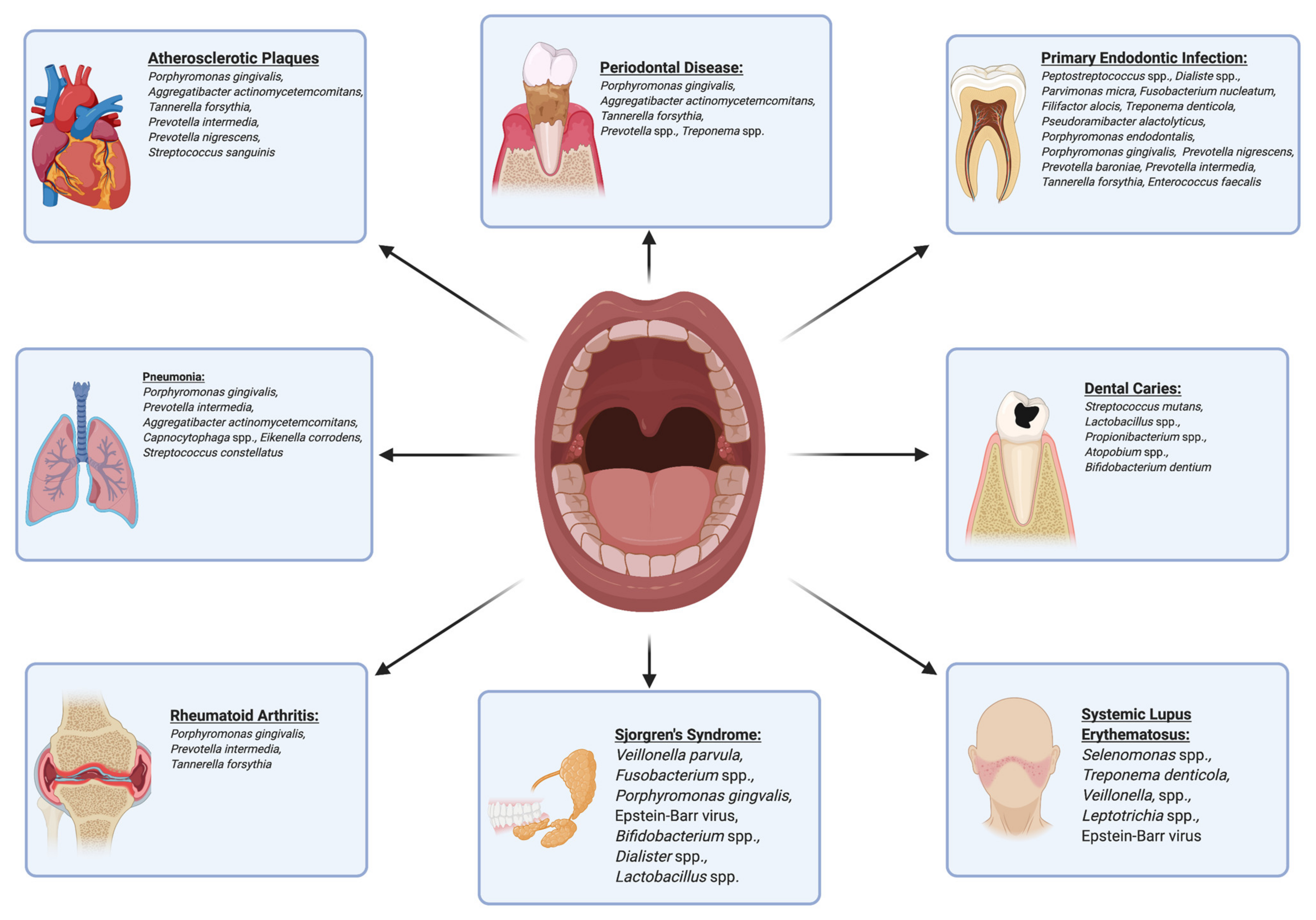
| Periodontitis | Aggregatibacter actinomycetemcomitans, Treponema spp., Prevotella spp. Tannerella forsythia, Treponema denticola, Porphyromonas gingivalis | Bacteria listed to the immediate left are all associated with periodontitis. Porphyromonas gingivalis, Tannerella forsythia, and Treponema denticola are all part of the “red complex,” which when present in large numbers, implicate a dysbiotic shift to periodontitis. | [4] [11] |
| Endodontic Infection | Peptostreptococcus spp., Dialister spp., Parvimonas micra, Fusobacterium nucleatum, Filifactor alocis, Treponema denticola, Pseudoramibacter alactolyticus, Porphyromonas endodontalis, Porphyromonas gingivalis, Prevotella nigrescens, Prevotella baroniae, Prevotella intermedia, Tannerella forsythia, Enterococcus faecalis | Primary infection of the pulp chamber commonly features the following species, while secondary infection of root canals is typically due to elevated levels of Enterococcus faecalis, Filifactor alocis, Pseudoramibacter alactolyticus, Parvimonas micra, Propionibacterium propionicum, Streptococcus constellatus, and Streptococcus anginosus. | [12] |
| Disease | Link to Oral/GI Microbiota | Significance | Ref. |
|---|---|---|---|
| Atherosclerotic Plaques | Porphyromonas gingivalis, Aggregatibacter actinomycetemcomitans, Tannerella forsythia, Prevotella intermedia, Prevotella nigrescens, Streptococcus sanguinis | These bacteria have been found in atherosclerotic plaque samples. Porphyromonas gingivalis and Aggregatibacter actinomycetemcomitans have shown high levels of inflammatory immune response and presence of these bacteria may lead to a significantly increased risk for developing coronary artery disease. | [14] |
| Pneumonia | Porphyromonas gingivalis, Prevotella intermedia, Aggregatibacter actinomycetemcomitans, Capnocytophaga spp., Eikenella corrodens, Streptococcus constellatus | These bacteria are thought to play direct roles in the pathogenesis of pneumonia. | [15] |
| Systemic Lupus Erythematosus (SLE) | Selenomonas spp., Treponema denticola, Veillonella spp., Leptotrichia spp. | Salivary levels of the following microorganisms have been shown to increase in patients with SLE and correlate directly with increased levels of inflammatory cytokines. | [49] |
| Systemic Lupus Erythematosus (SLE)/ Sjogren’s Syndrome (SS) | Lower Firmicutes to Bacteroidetes ratio | A lower Firmicutes to Bacteroidetes ratio has been shown in patients with SLE/SS and potentially increases inflammation. | [53] |
| Epstein–Barr virus (EBV) | EBV lytic phase antigens may be responsible for activation of SLE/SS immune responses creating auto-reactive antibodies. | [48] | |
| Sjogren’s Syndrome (SS) | Bifidobacterium spp., Dialister spp., Lactobacillus spp., Leptotrichia spp. | The first three bacteria are increased in salivary concentration for cases of primary SS. Leptotrichia spp. abundance was reduced in primary SS. | [55] |
| Veillonella parvula, Fusobacterium spp. | These bacteria have also shown elevated concentrations in patients with SS, with Veillonella parvula showing promise as a biomarker in the early detection of SS. | [56] | |
| Rheumatoid Arthritis (RA) | Porphyromonas gingivalis | Antibodies against human citrullinated alpha-enolase show cross reactivity with Porphyromonas gingivalis enolase and could be a potential source for autoimmunity directed against anticitrullinated protein antibodies (ACPAs). | [50] |
| Porphyromonas gingivalis, Prevotella intermedia, Tannerella forsythia | Patients with RA have elevated antibody levels against periodontal pathogens which correspond to increased serum levels of ACPAs and C-reactive protein. | [59] |
| Condition Treated by Fecal Microbiota Transplantation (FMT) | Findings | References |
|---|---|---|
| Recurrent Clostridium difficile infections | Studies provide evidence that FMT treatment for Clostridium difficile infections results in disease reduction with a success rate of approximately 92%. | [66] |
| Metabolic conditions including obesity and diabetes mellitus | Improved insulin sensitivity. Increased levels of short-chain fatty acid producing bacteria. Increased Roseburia intestinalis and Eubacterium hallii (butyrate-producing bacteria). Increased gut microbial diversity. Increased health-associated Bifidobacterium spp., Lactobacillus salivarius, Butyrivibrio spp., and Eubacterium spp. | [67,68] |
| Inflammatory Bowel Diseases (IBD) including Ulcerative Colitis (UC) and Crohn’s Disease (CD) | 45% entered disease remission; 22% clinical remission rate for patients with UC and 60.5% remission rate for patients with CD. | [69] |
| Autoimmune diseases: Multiple Sclerosis (MS) Idiopathic thrombocytopenic purpura (ITP) Ulcerative Colitis (UC) | Significantly improved gastrointestinal and neurological symptoms (MS) following FMT. Reduction in UC symptoms and an increase in platelet count to a normal level in patients with ITP. | [70,71] |
| Disease | Targeted Therapy for Dysbiosis in Systemic Diseases | Implication | Ref. |
|---|---|---|---|
| Rheumatoid arthritis (RA) | Disease modifying antirheumatic drugs Combination of methotrexate and T2 glycosides | Genetic and microbial sequencing in RA patients may provide foresight into treatment efficacy. | [46] |
| Colitis | Precision editing of microbiota Administration of tungstate | Specific targeting of dysbiosis causing microorganisms by inhibiting their pathogenicity and virulence factors Little effect on the complete microbiome. Host and other microbial taxa spared of negative effects. | [86] |
| Autism Spectrum Disorder (ASD) | Introduction of Lactobacillus reuteri | Precision medicine in ASD may lead to development of new strategies to rebalance the microbiome of these individuals, and perhaps reduce the associated morbidities. | [91] |
| Disease | Targeted Therapy for Dysbiosis in Oral Diseases | Implication | Reference |
|---|---|---|---|
| Caries | Immune complex administration via Guy’s 13 plantibody | Targeted therapy of Streptococcus mutans via inhibition of glucosyltransferases and PAc surface adhesin protein. | [94] |
| Introduction of Streptococcus dentisani | Novel strain from the Streptococcus mitis group shown to raise oral pH via breakdown of arginine into ammonia. | [98] | |
| Introduction of Streptococcus dentisani | Supernatants derived from Streptococcus dentisani have shown ability to inhibit pathogenic oral microorganisms including: Streptococcus mutans, Streptococcus sobrinus, Fusobacterium nucleatum, and Prevotella intermedia. | [97] | |
| Introduction of KSL decapeptide (KKVVFKVKFK–NH2) | KSL shows significant antimicrobial effects inhibiting biofilm formation of Streptococcus mutans and displaying antifungal properties against Candida albicans. | [99] | |
| Periodontitis | Introduction of SMAP 29 and SMAP 28 (Sheep myeloid antimicrobial peptides) | SMAP29 and the more potent SMAP28 exhibit non-specific antimicrobial properties targeting Porphyromonas gingivalis and Fusobacterium nucleatum among many others. | [100] |
| Introduction of IgG in combination with SMAPs | To increase specificity towards Porphyromonas gingivalis, specific IgG antibodies can be conjugated with SMAPs at a therapeutic concentration of 20 μg protein/mL. | [101] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khor, B.; Snow, M.; Herrman, E.; Ray, N.; Mansukhani, K.; Patel, K.A.; Said-Al-Naief, N.; Maier, T.; Machida, C.A. Interconnections between the Oral and Gut Microbiomes: Reversal of Microbial Dysbiosis and the Balance between Systemic Health and Disease. Microorganisms 2021, 9, 496. https://doi.org/10.3390/microorganisms9030496
Khor B, Snow M, Herrman E, Ray N, Mansukhani K, Patel KA, Said-Al-Naief N, Maier T, Machida CA. Interconnections between the Oral and Gut Microbiomes: Reversal of Microbial Dysbiosis and the Balance between Systemic Health and Disease. Microorganisms. 2021; 9(3):496. https://doi.org/10.3390/microorganisms9030496
Chicago/Turabian StyleKhor, Brandon, Michael Snow, Elisa Herrman, Nicholas Ray, Kunal Mansukhani, Karan A. Patel, Nasser Said-Al-Naief, Tom Maier, and Curtis A. Machida. 2021. "Interconnections between the Oral and Gut Microbiomes: Reversal of Microbial Dysbiosis and the Balance between Systemic Health and Disease" Microorganisms 9, no. 3: 496. https://doi.org/10.3390/microorganisms9030496
APA StyleKhor, B., Snow, M., Herrman, E., Ray, N., Mansukhani, K., Patel, K. A., Said-Al-Naief, N., Maier, T., & Machida, C. A. (2021). Interconnections between the Oral and Gut Microbiomes: Reversal of Microbial Dysbiosis and the Balance between Systemic Health and Disease. Microorganisms, 9(3), 496. https://doi.org/10.3390/microorganisms9030496