
Lower Recovery of Nontuberculous Mycobacteria from Outdoor Hawai’i Environmental Water Biofilms Compared to Indoor Samples

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Clinical Respiratory NTM Isolates
2.2. Environmental NTM Sampling
2.3. Recovery of NTM from Environmental Samples
2.4. NTM DNA Isolation
2.5. NTM Species and Subspecies Identification by Partial rpoB Gene Sequencing
2.6. Geospatial Modeling of NTM in Hawai’i through Machine Learning
2.7. Showerhead and Clinical Mycobacterium chimaera Phylogenetic Comparisons
2.8. Statistical Analyses
3. Results
3.1. M. chimaera Predominates Hawai’i Respiratory Samples
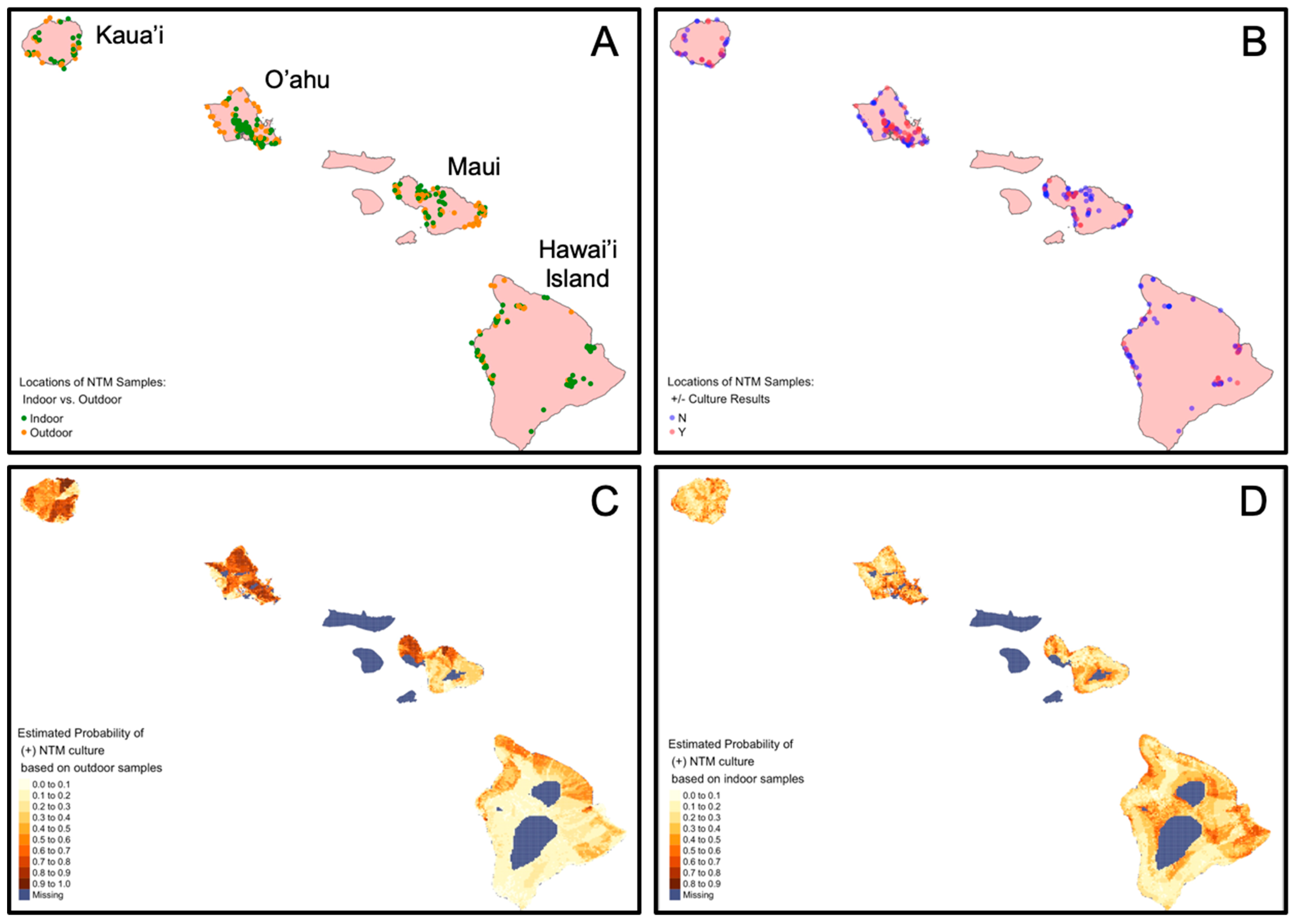
3.2. Geospatial Variables Associated with NTM between Outdoor and Indoor Samples
3.3. Outdoor Water Biofilm Samples Show Significantly Less NTM Compared to Indoor Water Biofilm Samples
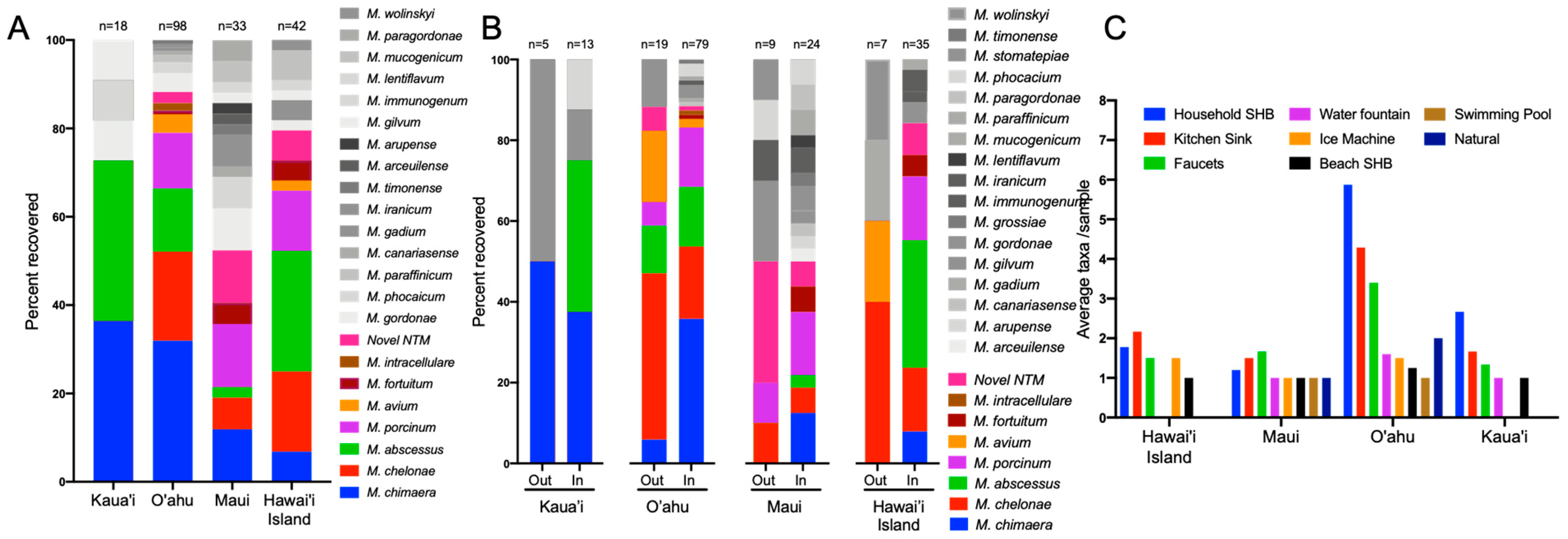
3.4. NTM Species Diversity and Recovery Proportions Vary by Island
3.5. Low Recovery of NTM from Natural Environments of Hawai’i
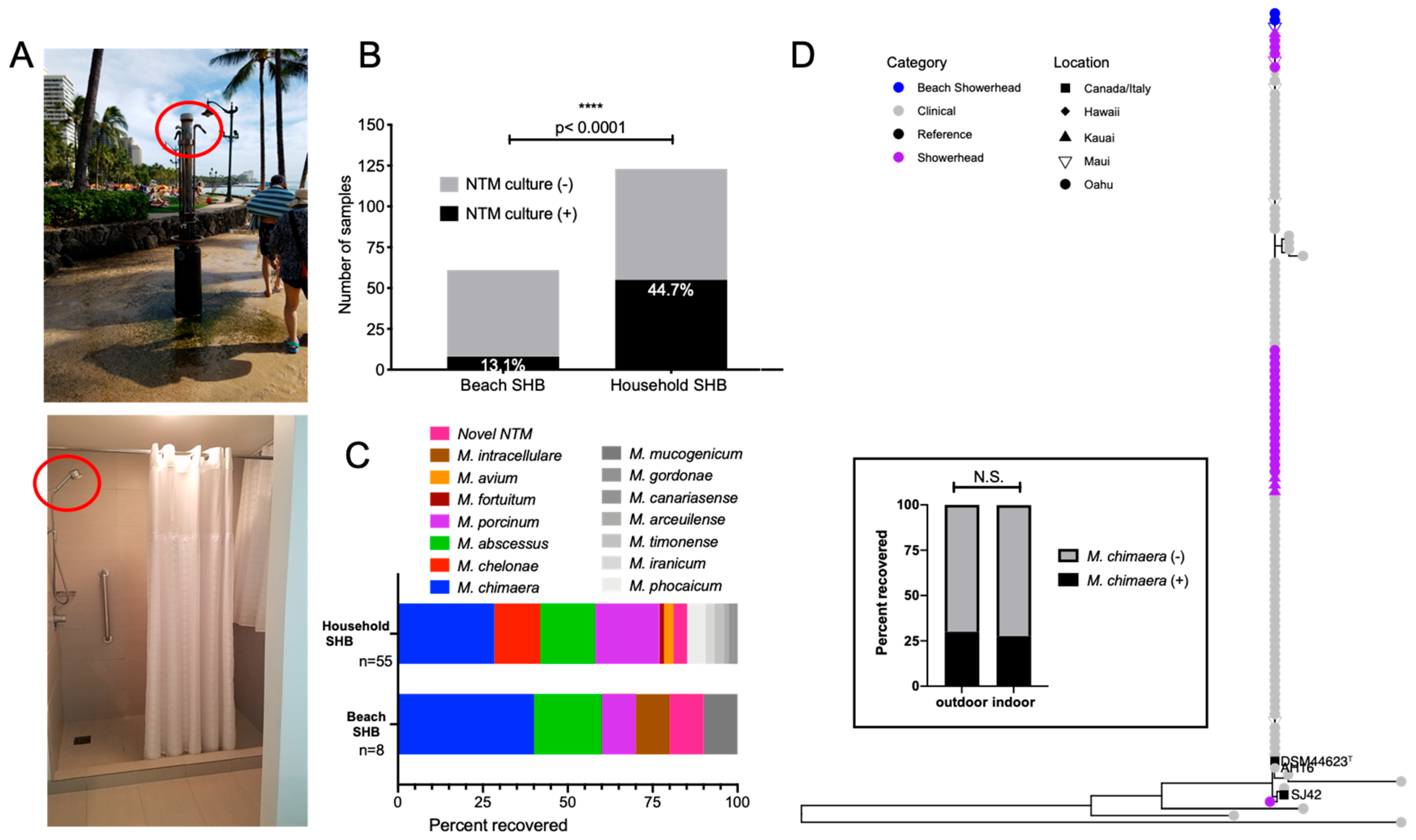
3.6. Significantly Less NTM from Outdoor Beach Showerhead Biofilms (SHB) Compared to Indoor SHB, but Equal Possibility of M. chimaera Exposure
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yeh, J.J.; Wang, Y.C.; Lin, C.L.; Chou, C.Y.; Yeh, T.C.; Wu, B.T.; Sung, F.C.; Kao, C.H. Nontuberculous mycobacterial infection is associated with increased respiratory failure: A nationwide cohort study. PLoS ONE 2014, 9, e99260. [Google Scholar] [CrossRef]
- Novosad, S.A.; Henkle, E.; Schafer, S.; Hedberg, K.; Ku, J.; Siegel, S.A.R.; Choi, D.; Slatore, C.G.; Winthrop, K.L. Mortality after Respiratory Isolation of Nontuberculous Mycobacteria. A Comparison of Patients Who Did and Did Not Meet Disease Criteria. Ann. Am. Thorac. Soc. 2017, 14, 1112–1119. [Google Scholar] [CrossRef] [PubMed]
- Park, T.Y.; Chong, S.; Jung, J.W.; Park, I.W.; Choi, B.W.; Lim, C.; Lee, C.U.; Kim, Y.S.; Choi, H.W.; Choi, J.C. Natural course of the nodular bronchiectatic form of Mycobacterium Avium complex lung disease: Long-term radiologic change without treatment. PLoS ONE 2017, 12, e0185774. [Google Scholar] [CrossRef]
- Daley, C.L.; Iaccarino, J.M.; Lange, C.; Cambau, E.; Wallace, R.J.; Andrejak, C.; Bottger, E.C.; Brozek, J.; Griffith, D.E.; Guglielmetti, L.; et al. Treatment of Nontuberculous Mycobacterial Pulmonary Disease: An Official ATS/ERS/ESCMID/IDSA Clinical Practice Guideline. Clin. Infect. Dis. 2020, 71, 905–913. [Google Scholar] [CrossRef] [PubMed]
- Adjemian, J.; Olivier, K.N.; Seitz, A.E.; Holland, S.M.; Prevots, D.R. Prevalence of nontuberculous mycobacterial lung disease in U.S. Medicare beneficiaries. Am. J. Respir. Crit. Care Med. 2012, 185, 881–886. [Google Scholar] [CrossRef]
- Nouioui, I.; Carro, L.; Garcia-Lopez, M.; Meier-Kolthoff, J.P.; Woyke, T.; Kyrpides, N.C.; Pukall, R.; Klenk, H.P.; Goodfellow, M.; Goker, M. Genome-Based Taxonomic Classification of the Phylum Actinobacteria. Front. Microbiol. 2018, 9, 2007. [Google Scholar] [CrossRef]
- Oren, A.; Garrity, G. List of new names and new combinations previously effectively, but not validly, published. Int. J. Syst. Evol. Microbiol. 2018, 68, 1411–1417. [Google Scholar] [CrossRef]
- Honda, J.R.; Hasan, N.A.; Davidson, R.M.; Williams, M.D.; Epperson, L.E.; Reynolds, P.R.; Smith, T.; Iakhiaeva, E.; Bankowski, M.J.; Wallace, R.J., Jr.; et al. Environmental Nontuberculous Mycobacteria in the Hawaiian Islands. PLoS Negl. Trop. Dis. 2016, 10, e0005068. [Google Scholar] [CrossRef]
- Kazda, J.; Muller, K.; Irgens, L.M. Cultivable mycobacteria in sphagnum vegetation of moors in South Sweden and coastal Norway. Acta. Pathol. Microbiol. Scand. B 1979, 87B, 97–101. [Google Scholar] [CrossRef]
- Eila, K.; Iivanainen, P.J.M.; Räisänen, M.L.; Katila, M.L. Mycobacteria in boreal coniferous forest soils. FEMS Microbiol. Ecol. 1997, 23, 325–332. [Google Scholar]
- Thorel, M.F.; Falkinham, J.O., 3rd; Moreau, R.G. Environmental mycobacteria from alpine and subalpine habitats. FEMS Microbiol. Ecol. 2004, 49, 343–347. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Griffith, D.E.; Aksamit, T.; Brown-Elliott, B.A.; Catanzaro, A.; Daley, C.; Gordin, F.; Holland, S.M.; Horsburgh, R.; Huitt, G.; Iademarco, M.F.; et al. An official ATS/IDSA statement: Diagnosis, treatment, and prevention of nontuberculous mycobacterial diseases. Am. J. Respir. Crit. Care Med. 2007, 175, 367–416. [Google Scholar] [CrossRef] [PubMed]
- Falkinham, J.O.; Norton, C.D.; LeChevallier, M.W. Factors influencing numbers of Mycobacterium avium, Mycobacterium intracellulare, and other Mycobacteria in drinking water distribution systems. Appl. Environ. Microbiol. 2001, 67, 1225–1231. [Google Scholar] [CrossRef] [PubMed]
- Epperson, L.E.; Strong, M. A scalable, efficient, and safe method to prepare high quality DNA from mycobacteria and other challenging cells. J. Clin. Tuberc. Other Mycobact. Dis. 2020, 19, 100150. [Google Scholar] [CrossRef] [PubMed]
- Telenti, A.; Marchesi, F.; Balz, M.; Bally, F.; Bottger, E.C.; Bodmer, T. Rapid identification of mycobacteria to the species level by polymerase chain reaction and restriction enzyme analysis. J. Clin. Microbiol. 1993, 31, 175–178. [Google Scholar] [CrossRef]
- Adekambi, T.; Colson, P.; Drancourt, M. rpoB-based identification of nonpigmented and late-pigmenting rapidly growing mycobacteria. J. Clin. Microbiol. 2003, 41, 5699–5708. [Google Scholar] [CrossRef]
- Kuhn, M. Caret: Classification and Regression Training. 2020. Available online: https://CRAN.R-project.org/package=caret. (accessed on 30 November 2020).
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2020; Available online: https://www.r-project.org (accessed on 30 November 2020).
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Stecher, G.; Tamura, K.; Kumar, S. Molecular Evolutionary Genetics Analysis (MEGA) for macOS. Mol. Biol. Evol. 2020, 37, 1237–1239. [Google Scholar] [CrossRef]
- Yu, G.; Smith, D.K.; Zhu, H.; Guan, Y.; Lam, T.T.Y. Ggtree: An R package for visualization and annotation of phylogenetic trees with their covariates and other associated data. Methods Ecol. Evol. 2017, 8, 28–36. [Google Scholar] [CrossRef]
- Casal, M.; Calero, J.R. Mycobacterium gadium sp. nov. a new species of rapid-growing scotochromogenic mycobacteria. Tubercle 1974, 55, 299–308. [Google Scholar] [CrossRef]
- Jimenez, M.S.; Campos-Herrero, M.I.; Garcia, D.; Luquin, M.; Herrera, L.; Garcia, M.J. Mycobacterium canariasense sp. nov. Int. J. Syst. Evol. Microbiol. 2004, 54, 1729–1734. [Google Scholar] [CrossRef] [PubMed]
- Gebert, M.; Delgado-Baquerizo, M.; Oliverio, A.; Webster, T.; Nichols, L.; Honda, J.; Chan, E.; Adjemian, J.; Dunn, R.; Fierer, N. Ecological analyses of mycobacteria in showerhead biofilms and their relevance to human health. bioRxiv 2018. [Google Scholar] [CrossRef] [PubMed]
- Feazel, L.M.; Baumgartner, L.K.; Peterson, K.L.; Frank, D.N.; Harris, J.K.; Pace, N.R. Opportunistic pathogens enriched in showerhead biofilms. Proc. Natl. Acad. Sci. USA 2009, 106, 16393–16399. [Google Scholar] [CrossRef] [PubMed]
- Adjemian, J.; Olivier, K.N.; Seitz, A.E.; Falkinham, J.O., 3rd; Holland, S.M.; Prevots, D.R. Spatial clusters of nontuberculous mycobacterial lung disease in the United States. Am. J. Respir Crit. Care Med. 2012, 186, 553–558. [Google Scholar] [CrossRef]
- Corumluoglu, O.; Asri, I. The effect of urban heat island on Izmir’s city ecosystem and climate. Environ. Sci. Pollut. Res. Int. 2015, 22, 3202–3211. [Google Scholar] [CrossRef]
- Mirsaeidi, M.; Vu, A.; Leitman, P.; Sharifi, A.; Wisliceny, S.; Leitman, A.; Schmid, A.; Campos, M.; Falkinham, J.; Salathe, M. A Patient-Based Analysis of the Geographic Distribution of Mycobacterium avium complex, Mycobacterium abscessus, and Mycobacterium kansasii Infections in the United States. Chest 2017, 151, 947–950. [Google Scholar] [CrossRef]
- Cassidy, P.M.; Hedberg, K.; Saulson, A.; McNelly, E.; Winthrop, K.L. Nontuberculous mycobacterial disease prevalence and risk factors: A changing epidemiology. Clin. Infect. Dis. 2009, 49, e124–e129. [Google Scholar] [CrossRef]
- Glickman, C.M.; Virdi, R.; Hasan, N.A.; Epperson, L.E.; Brown, L.; Dawrs, S.N.; Crooks, J.L.; Chan, E.D.; Strong, M.; Nelson, S.T.; et al. Assessment of soil features on the growth of environmental nontuberculous mycobacterial isolates from Hawai’i. Appl. Environ. Microbiol. 2020. [Google Scholar] [CrossRef]
- Hasan, N.A.; Epperson, L.E.; Lawsin, A.; Rodger, R.R.; Perkins, K.M.; Halpin, A.L.; Perry, K.A.; Moulton-Meissner, H.; Diekema, D.J.; Crist, M.B.; et al. Genomic Analysis of Cardiac Surgery-Associated Mycobacterium chimaera Infections, United States. Emerg. Infect. Dis. 2019, 25, 559–563. [Google Scholar] [CrossRef]
- Walker, J.; Moore, G.; Collins, S.; Parks, S.; Garvey, M.I.; Lamagni, T.; Smith, G.; Dawkin, L.; Goldenberg, S.; Chand, M. Microbiological problems and biofilms associated with Mycobacterium chimaera in heater-cooler units used for cardiopulmonary bypass. J. Hosp. Infect. 2017, 96, 209–220. [Google Scholar] [CrossRef]
- September, S.M.; Brozel, V.S.; Venter, S.N. Diversity of nontuberculoid Mycobacterium species in biofilms of urban and semiurban drinking water distribution systems. Appl. Environ. Microbiol. 2004, 70, 7571–7573. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.C.; Chiou, C.S.; Chen, J.H.; Shen, G.H. Molecular epidemiology of Mycobacterium abscessus infections in a subtropical chronic ventilatory setting. J. Med. Microbiol. 2010, 59, 1203–1211. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Van Ingen, J.; Blaak, H.; de Beer, J.; de Roda Husman, A.M.; van Soolingen, D. Rapidly growing nontuberculous mycobacteria cultured from home tap and shower water. Appl. Environ. Microbiol. 2010, 76, 6017–6019. [Google Scholar] [CrossRef] [PubMed]
- Thomson, R.; Tolson, C.; Sidjabat, H.; Huygens, F.; Hargreaves, M. Mycobacterium abscessus isolated from municipal water—a potential source of human infection. BMC Infect. Dis. 2013, 13, 241. [Google Scholar] [CrossRef] [PubMed]
- Spaulding, A.B.; Lai, Y.L.; Zelazny, A.M.; Olivier, K.N.; Kadri, S.S.; Prevots, D.R.; Adjemian, J. Geographic Distribution of Nontuberculous Mycobacterial Species Identified among Clinical Isolates in the United States, 2009–2013. Ann. Am. Thorac. Soc. 2017. [Google Scholar] [CrossRef]
- Yano, H.; Iwamoto, T.; Nishiuchi, Y.; Nakajima, C.; Starkova, D.A.; Mokrousov, I.; Narvskaya, O.; Yoshida, S.; Arikawa, K.; Nakanishi, N.; et al. Population Structure and Local Adaptation of MAC Lung Disease Agent Mycobacterium avium subsp. hominissuis. Genome Biol. Evol. 2017, 9, 2403–2417. [Google Scholar] [CrossRef]
- Arikawa, K.; Ichijo, T.; Nakajima, S.; Nishiuchi, Y.; Yano, H.; Tamaru, A.; Yoshida, S.; Maruyama, F.; Ota, A.; Nasu, M.; et al. Genetic relatedness of Mycobacterium avium subsp. hominissuis isolates from bathrooms of healthy volunteers, rivers, and soils in Japan with human clinical isolates from different geographical areas. Infect. Genet. Evol. 2019, 74, 103923. [Google Scholar] [CrossRef]
- Lahiri, A.; Kneisel, J.; Kloster, I.; Kamal, E.; Lewin, A. Abundance of Mycobacterium avium ssp. hominissuis in soil and dust in Germany—Implications for the infection route. Lett. Appl. Microbiol. 2014, 59, 65–70. [Google Scholar] [CrossRef]
- Falkinham, J.O., 3rd. Environmental sources of nontuberculous mycobacteria. Clin. Chest Med. 2015, 36, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Lande, L.; Alexander, D.C.; Wallace, R.J., Jr.; Kwait, R.; Iakhiaeva, E.; Williams, M.; Cameron, A.D.S.; Olshefsky, S.; Devon, R.; Vasireddy, R.; et al. Mycobacterium avium in Community and Household Water, Suburban Philadelphia, Pennsylvania, USA, 2010–2012. Emerg Infect. Dis. 2019, 25, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Falkinham, J.O., 3rd. Nontuberculous mycobacteria from household plumbing of patients with nontuberculous mycobacteria disease. Emerg. Infect. Dis. 2011, 17, 419–424. [Google Scholar] [CrossRef] [PubMed]
- Covert, T.C.; Rodgers, M.R.; Reyes, A.L.; Stelma, G.N. Occurrence of nontuberculous mycobacteria in environmental samples. Appl. Environ. Microbiol. 1999, 65, 2492–2496. [Google Scholar] [CrossRef] [PubMed]
- Jeon, D. Infection Source and Epidemiology of Nontuberculous Mycobacterial Lung Disease. Tuberc. Respir Dis. 2019, 82, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Strollo, S.E.; Adjemian, J.; Adjemian, M.K.; Prevots, D.R. The Burden of Pulmonary Nontuberculous Mycobacterial Disease in the United States. Ann. Am. Thorac. Soc. 2015. [Google Scholar] [CrossRef] [PubMed]
- Winthrop, K.L.; Henkle, E.; Walker, A.; Cassidy, M.; Hedberg, K.; Schafer, S. On the Reportability of Nontuberculous Mycobacterial Disease to Public Health Authorities. Ann. Am. Thorac. Soc. 2017, 14, 314–317. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Outdoor (n = 179) | Indoor (n = 366) |
|---|---|
| Outdoor (beach) showerheads (n = 61) | Showerheads (n = 123) |
| Water fountain (n = 24) | Kitchen sink faucets (n = 84) |
| Outdoor faucets (n = 9) | Bathroom sink faucets (n = 154) |
| Ice machines (n = 12) | Water dispensers (n = 5) |
| Swimming pools (n = 12) | |
| Freshwater streams (n = 28) Sphagnum moss (n = 26) | |
| Lava rock (n = 7) |
| Category | Total Isolates (n=119) | Beach Showerheads (Mean SNPs ± SD) | Indoor Showerheads (Mean SNPs ± SD) | Clinical Respiratory (Mean SNPs ± SD) |
|---|---|---|---|---|
| Beach Showerheads | 3 | 0 SNPs ± 0 | ||
| Indoor Showerheads | 29 | 0 SNPs ± 0 | 0 SNPs ± 0 | |
| Clinical Respiratory | 87 | 0 SNPs ± 8.62 | 1.72 SNPs ± 8.62 | 3.4 SNPs ± 11.99 |
| Rank: | Outdoor: | Indoor: |
|---|---|---|
| 1 | Elevation | Evapotranspiration |
| 2 | Lowest income quintile | Elevation |
| 3 | Population | Rainfall |
| 4 | Watershed area | Median gross rent |
| 5 | Population of Pacific Islanders | GINI coefficient |
| 6 | GINI coefficient | Land development indicator |
| 7 | Median gross rent | Watershed area |
| 8 | Rainfall | Population with multiracial ancestry |
| 9 | Rural population | Soil water infiltration rate |
| 10 | Population with multiracial ancestry | Population Pacific Islander |
| 11 | Evapotranspiration | Lowest income quintile |
| 12 | Soil water infiltration rate | Population on public assistance |
| 13 | Land development indicator | Soil pH |
| 14 | Soil organic matter | Relative geologic age |
| 15 | Soil cation-exchange capacity | Relative humidity |
| 16 | Soil shrink-swell capacity | Surface temperature |
| 17 | Urban population | Urban population |
| 18 | Relative geologic age | Soil cation-exchange capacity |
| 19 | Relative humidity | Population |
| 20 | Soil pH | Soil organic matter |
| Outdoor Samples | Indoor Samples | ||||
|---|---|---|---|---|---|
| Total (n=) | % NTM Culture Positive | Total (n=) | % NTM Culture Positive | p-Value | |
| Kaua’i | 21 | 23.8% (5) | 30 | 43.3% (13) | not significant |
| O’ahu | 71 | 12.7% (9) | 146 | 54.1% (79) | <0.0001 |
| Maui | 56 | 33.9% (19) | 88 | 27.3% (24) | not significant |
| Hawai’i | 31 | 22.6% (7) | 102 | 34.3% (35) | not significant |
| Totals | 179 | 22.3% (40) | 366 | 41.3% (151) | <0.0001 |
| (a) | |||||
| Freshwater Streams | Moss | Lava Rock | |||
| Kaua’i | 0/3 (0%) | Not sampled | Not sampled | ||
| O’ahu | 8/10 (80%) | 0/2 (0%) | Not sampled | ||
| Maui | 1/11 (9%) | 2/19 (10.5%) | 0/3 (0%) | ||
| Hawai’i Island | 0/4 (0%) | 1/5 (20%) | 0/4 (0%) | ||
| Total | 9/28 (32.1%) | 3/26 (11.5%) | 0/7 (0%) | ||
| (b) | |||||
| Island: | Freshwater Stream: | NTM Species Identified from Freshwater Streams: | Island: | Moss: | NTM Species Identified from Moss: |
| O’ahu | Stream 1 | M. avium | Maui | Moss 1 | M. gordonae; M. iranicum; M. mucogenicum |
| Stream 2 | M. chelonae | Moss 2 | Novel NTM | ||
| Stream 3 | M. avium; M. chimaera | Hawai’i Island | Moss 1 | M. rhodesiae | |
| Stream 4 | M. abscessus | ||||
| Stream 5 | M. avium; M. timonese | ||||
| Stream 6 | M. chelonae; M. intracellulare | ||||
| Stream 7 | M. chelonae; M. chimaera | ||||
| Stream 8 | M. chelonae | ||||
| Maui | Stream 1 | Novel NTM | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Virdi, R.; Lowe, M.E.; Norton, G.J.; Dawrs, S.N.; Hasan, N.A.; Epperson, L.E.; Glickman, C.M.; Chan, E.D.; Strong, M.; Crooks, J.L.; et al. Lower Recovery of Nontuberculous Mycobacteria from Outdoor Hawai’i Environmental Water Biofilms Compared to Indoor Samples. Microorganisms 2021, 9, 224. https://doi.org/10.3390/microorganisms9020224
Virdi R, Lowe ME, Norton GJ, Dawrs SN, Hasan NA, Epperson LE, Glickman CM, Chan ED, Strong M, Crooks JL, et al. Lower Recovery of Nontuberculous Mycobacteria from Outdoor Hawai’i Environmental Water Biofilms Compared to Indoor Samples. Microorganisms. 2021; 9(2):224. https://doi.org/10.3390/microorganisms9020224
Chicago/Turabian StyleVirdi, Ravleen, Melissa E. Lowe, Grant J. Norton, Stephanie N. Dawrs, Nabeeh A. Hasan, L. Elaine Epperson, Cody M. Glickman, Edward D. Chan, Michael Strong, James L. Crooks, and et al. 2021. "Lower Recovery of Nontuberculous Mycobacteria from Outdoor Hawai’i Environmental Water Biofilms Compared to Indoor Samples" Microorganisms 9, no. 2: 224. https://doi.org/10.3390/microorganisms9020224
APA StyleVirdi, R., Lowe, M. E., Norton, G. J., Dawrs, S. N., Hasan, N. A., Epperson, L. E., Glickman, C. M., Chan, E. D., Strong, M., Crooks, J. L., & Honda, J. R. (2021). Lower Recovery of Nontuberculous Mycobacteria from Outdoor Hawai’i Environmental Water Biofilms Compared to Indoor Samples. Microorganisms, 9(2), 224. https://doi.org/10.3390/microorganisms9020224