
Assessing Gut Microbiota in an Infant with Congenital Propionic Acidemia before and after Probiotic Supplementation

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
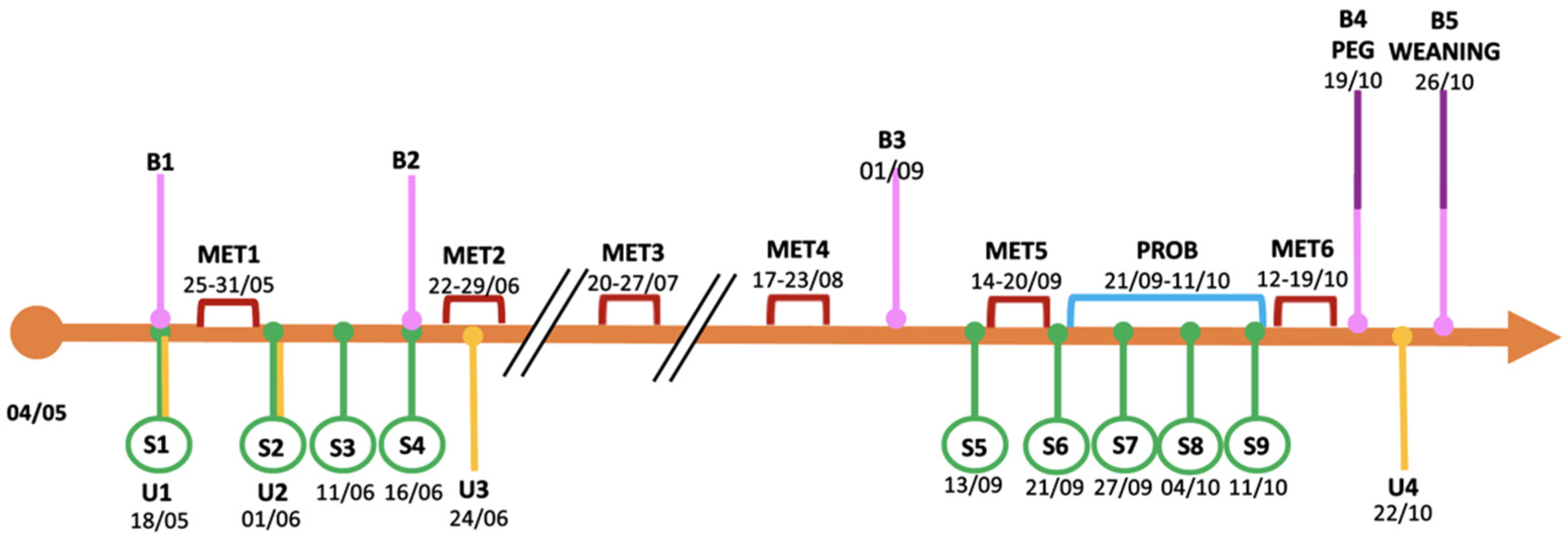
2.1. Patient Description
2.2. Clinical and Biochemical Disease Specific Follow-Up
2.3. Diet Monitoring
2.4. Urine Analysis with the Electronic Nose
2.5. Probiotic Supplementation
2.6. Stool Sample Collection and Sequencing
2.7. Bioinformatic Analysis
3. Results
3.1. Clinical and Biochemical Outcomes
3.2. Dietary and Nutritional Issues
3.3. Urine Analysis
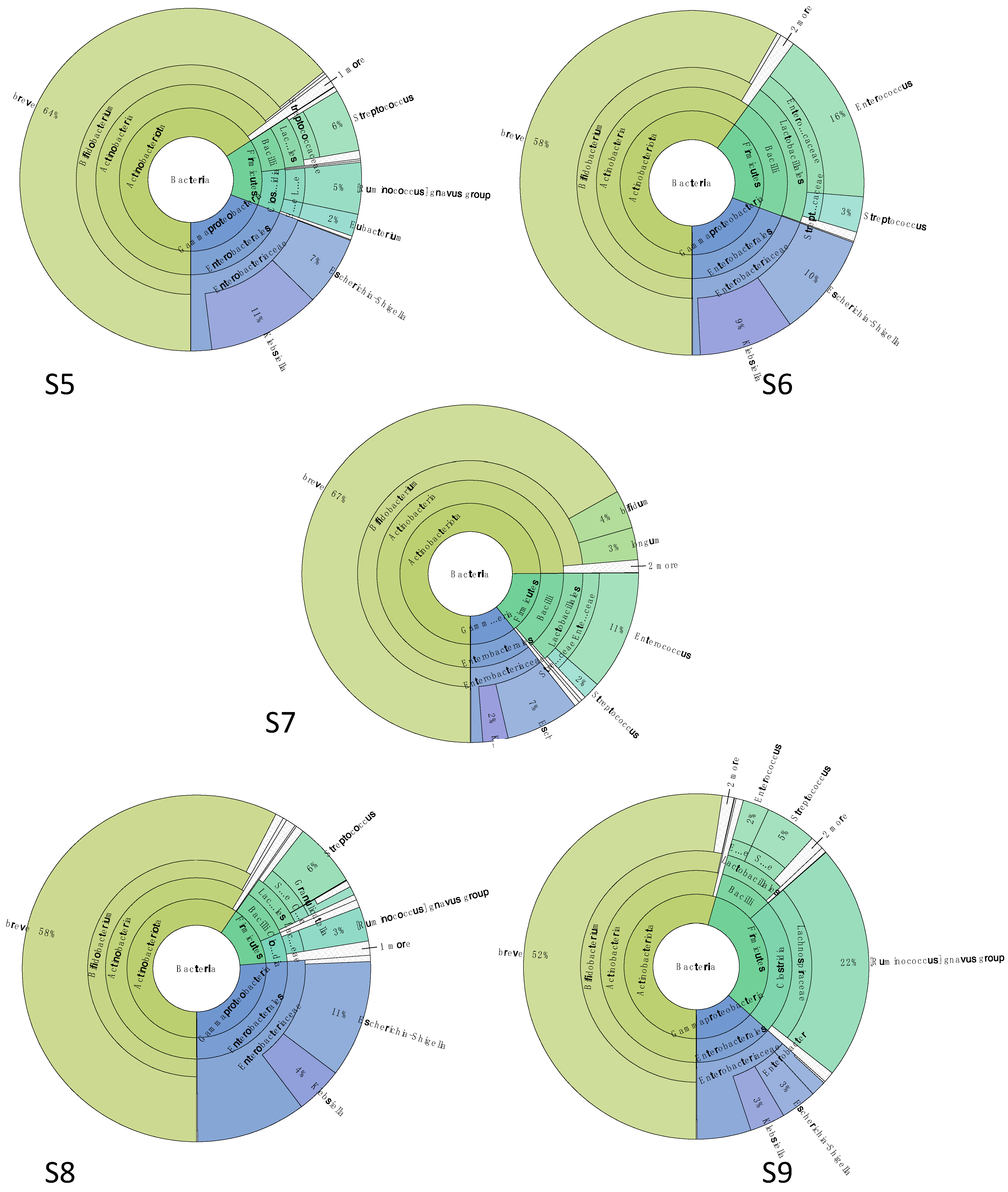
3.4. Microbiota Data
3.4.1. Alpha- and Beta- Diversity Analysis Shows the Impact of Antibiotic Treatment
3.4.2. Microbiota Assessment at First Month of Life Unravels Bac. fragilis as Propionate Producer
3.4.3. Gut Microbiota Was Characterized by Bifidobacterium spp. and Ruminococcus gnavus after Probiotic Supplementation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wendel, U.; de Baulny, H.O. Branched-Chain Organic Acidurias/Acidemias. In Inborn Metabolic Diseases: Diagnosis and Treatment; Fernandes, J., Saudubray, J.-M., van den Berghe, G., Walter, J.H., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 245–262. [Google Scholar]
- Jurecki, E.; Ueda, K.; Frazier, D.; Rohr, F.; Thompson, A.; Hussa, C.; Obernolte, L.; Reineking, B.; Roberts, A.M.; Yannicelli, S.; et al. Nutrition management guideline for propionic acidemia: An evidence- and consensus-based approach. Mol. Genet. Metab. 2019, 126, 341–354. [Google Scholar] [CrossRef]
- Pena, L.; Franks, J.; Chapman, K.A.; Gropman, A.; Ah Mew, N.; Chakrapani, A.; Island, E.; MacLeod, E.; Matern, D.; Smith, B.; et al. Natural history of propionic acidemia. Mol. Genet. Metab. 2012, 105, 5–9. [Google Scholar] [CrossRef]
- Baumgartner, M.R.; Hörster, F.; Dionisi-Vici, C.; Haliloglu, G.; Karall, D.; Chapman, K.A.; Huemer, M.; Hochuli, M.; Assoun, M.; Ballhausen, D.; et al. Proposed guidelines for the diagnosis and management of methylmalonic and propionic acidemia. Orphanet J. Rare Dis. 2014, 9, 130. [Google Scholar] [CrossRef] [PubMed]
- Fenton, W.A.; Gravel, R.A.; Rosenblatt, D.S. Disorders of Propionate and Methylmalonate Metabolism. In The Metabolic and Molecular Bases of Inherited Disease, 8th ed.; Scriver, C.R., Beaudet, A.L., Sly, W.S., Valle, D., Eds.; McGraw-Hill: New York, NY, USA, 2001; Volume II, p. 2176. [Google Scholar]
- Forny, P.; Hörster, F.; Ballhausen, D.; Chakrapani, A.; Chapman, K.A.; Dionisi-Vici, C.; Dixon, M.; Grünert, S.C.; Grunewald, S.; Haliloglu, G.; et al. Guidelines for the diagnosis and management of methylmalonic acidaemia and propionic acidaemia: First revision. J. Inherit. Metab. Dis. 2021, 44, 566–592. [Google Scholar] [CrossRef] [PubMed]
- Kirby, T.O.; Brown, M.; Ochoa-Repáraz, J.; Roullet, J.B.; Gibson, K.M. Microbiota Manipulation as a Metagenomic Therapeutic Approach for Rare Inherited Metabolic Disorders. Clin. Pharmacol. Ther. 2019, 106, 505–507. [Google Scholar] [CrossRef] [PubMed]
- Richard, E.; Desviat, L.R.; Pérez, B.; Pérez-Cerdá, C.; Ugarte, M. Genetic heterogeneity in propionic acidemia patients with alpha-subunit defects. Identification of five novel mutations, one of them causing instability of the protein. Biochim. Biophys. Acta 1999, 1453, 351–358. [Google Scholar] [CrossRef]
- Kraus, J.P.; Spector, E.; Venezia, S.; Estes, P.; Chiang, P.W.; Creadon-Swindell, G.; Müllerleile, S.; de Silva, L.; Barth, M.; Walter, M.; et al. Mutation analysis in 54 propionic acidemia patients. J. Inherit. Metab. Dis. 2012, 35, 51–63. [Google Scholar] [CrossRef]
- FAO/WHO/ONU. Protein and Amino Acid Requirements in Human Nutrition; Report of a Joint FAO/WHO/UNU Expert Consultation on Food and Agriculture Organization: Geneva, Switzerland, 2002. [Google Scholar]
- Takahashi, S.; Tomita, J.; Nishioka, K.; Hisada, T.; Nishijima, M. Development of a prokaryotic universal primer for simultaneous analysis of Bacteria and Archaea using next-generation sequencing. PLoS ONE 2014, 9, e105592. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 3. [Google Scholar] [CrossRef]
- Wright, E.S. Using DECIPHERv2.0 to Analyze Big Biological Sequence Data in R. R J. 2016, 8, 352–359. [Google Scholar] [CrossRef]
- Schliep, K.P. phangorn: Phylogenetic analysis in R. Bioinformatics 2011, 27, 592–593. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Wood, D.E.; Lu, J.; Langmead, B. Improved metagenomic analysis with Kraken 2. Genome Biol. 2019, 20, 257. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Breitwieser, F.P.; Thielen, P.; Salzberg, S.L. Bracken: Estimating species abundance in metagenomics data. PeerJ Comput. Sci. 2017, 3, e104. [Google Scholar] [CrossRef]
- Ondov, B.D.; Bergman, N.H.; Phillippy, A.M. Interactive metagenomic visualization in a Web browser. BMC Bioinform. 2011, 12, 385. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Maines, E.; Catesini, G.; Boenzi, S.; Mosca, A.; Candusso, M.; Dello Strologo, L.; Martinelli, D.; Maiorana, A.; Liguori, A.; Olivieri, G.; et al. Plasma methylcitric acid and its correlations with other disease biomarkers: The impact in the follow up of patients with propionic and methylmalonic acidemia. J. Inherit. Metab. Dis. 2020, 43, 1173–1185. [Google Scholar] [CrossRef] [PubMed]
- Montanari, C.; Parolisi, S.; Borghi, E.; Putignani, L.; Bassanini, G.; Zuvadelli, J.; Bonfanti, C.; Tummolo, A.; Dionisi Vici, C.; Biasucci, G.; et al. Dysbiosis, Host Metabolism, and Non-communicable Diseases: Trialogue in the Inborn Errors of Metabolism. Front. Physiol. 2021, 12, 716520. [Google Scholar] [CrossRef]
- Grünert, S.C.; Müllerleile, S.; De Silva, L.; Barth, M.; Walter, M.; Walter, K.; Meissner, T.; Lindner, M.; Ensenauer, R.; Santer, R.; et al. Propionic acidemia: Clinical course and outcome in 55 pediatric and adolescent patients. Orphanet J. Rare Dis. 2013, 8, 6. [Google Scholar] [CrossRef]
- Daly, A.; Pinto, A.; Evans, S.; Almeida, M.F.; Assoun, M.; Belanger-Quintana, A.; Bernabei, S.M.; Bollhalder, S.; Cassiman, D.; Champion, H.; et al. Dietary practices in propionic acidemia: A European survey. Mol. Genet. Metab. Rep. 2017, 13, 83–89. [Google Scholar] [CrossRef]
- Molema, F.; Haijes, H.A.; Janssen, M.C.; Bosch, A.M.; van Spronsen, F.J.; Mulder, M.F.; Verhoeven-Duif, N.M.; Jans, J.J.M.; van der Ploeg, A.T.; Wagenmakers, M.A.; et al. High protein prescription in methylmalonic and propionic acidemia patients and its negative association with long-term outcome. Clin. Nutr. 2021, 40, 3622–3630. [Google Scholar] [CrossRef]
- Burlina, A.; Tims, S.; van Spronsen, F.; Sperl, W.; Burlina, A.P.; Kuhn, M.; Knol, J.; Rakhshandehroo, M.; Coşkun, T.; Singh, R.H.; et al. The potential role of gut microbiota and its modulators in the management of propionic and methylmalonic acidemia. Expert Opin. Orphan Drugs 2018, 6, 683–692. [Google Scholar] [CrossRef]
- Leonard, J.V. Stable isotope studies in propionic and methylmalonic acidaemia. Eur. J. Pediatr. 1997, 156 (Suppl. 1), S67–S69. [Google Scholar] [CrossRef] [PubMed]
- Rafique, M. Clinical spectrum of propionic acidaemia. J. Nutr. Metab. 2013, 2013, 975964. [Google Scholar] [CrossRef] [PubMed]
- Thompson, G.N.; Chalmers, R.A.; Walter, J.H.; Bresson, J.L.; Lyonnet, S.L.; Reed, P.J.; Saudubray, J.M.; Leonard, J.V.; Halliday, D. The use of metronidazole in management of methylmalonic and propionic acidaemias. Eur. J. Pediatr. 1990, 149, 792–796. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Marques, J.; Hout, A.; Ferreira, R.M.; Weber, M.; Pinto-Ribeiro, I.; van Doorn, L.J.; Knetsch, C.W.; Figueiredo, C. Impact of Host DNA and Sequencing Depth on the Taxonomic Resolution of Whole Metagenome Sequencing for Microbiome Analysis. Front. Microbiol. 2019, 10, 1277. [Google Scholar] [CrossRef]
- Dalby, M.J.; Hall, L.J. Recent advances in understanding the neonatal microbiome. F1000Research 2020, 9. [Google Scholar] [CrossRef]
- Sagheddu, V.; Patrone, V.; Miragoli, F.; Puglisi, E.; Morelli, L. Infant Early Gut Colonization by Lachnospiraceae: High Frequency of Ruminococcus gnavus. Front. Pediatr. 2016, 4, 57. [Google Scholar] [CrossRef]
- Di Gioia, D.; Aloisio, I.; Mazzola, G.; Biavati, B. Bifidobacteria: Their impact on gut microbiota composition and their applications as probiotics in infants. Appl. Microbiol. Biotechnol. 2014, 98, 563–577. [Google Scholar] [CrossRef]
- Yatsunenko, T.; Rey, F.E.; Manary, M.J.; Trehan, I.; Dominguez-Bello, M.G.; Contreras, M.; Magris, M.; Hidalgo, G.; Baldassano, R.N.; Anokhin, A.P.; et al. Human gut microbiome viewed across age and geography. Nature 2012, 486, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Rios-Covian, D.; Cuesta, I.; Alvarez-Buylla, J.R.; Ruas-Madiedo, P.; Gueimonde, M.; de Los Reyes-Gavilán, C.G. Bacteroides fragilis metabolises exopolysaccharides produced by bifidobacteria. BMC Microbiol. 2016, 16, 150. [Google Scholar] [CrossRef]
- Reichardt, N.; Duncan, S.H.; Young, P.; Belenguer, A.; McWilliam Leitch, C.; Scott, K.P.; Flint, H.J.; Louis, P. Phylogenetic distribution of three pathways for propionate production within the human gut microbiota. ISME J. 2014, 8, 1323–1335. [Google Scholar] [CrossRef]
- Wexler, H.M. Bacteroides: The good, the bad, and the nitty-gritty. Clin. Microbiol Rev. 2007, 20, 593–621. [Google Scholar] [CrossRef]
- Löfmark, S.; Edlund, C.; Nord, C.E. Metronidazole is still the drug of choice for treatment of anaerobic infections. Clin. Infect. Dis. 2010, 50 (Suppl. 1), S16–S23. [Google Scholar] [CrossRef]
- Cao, L.; Chen, H.; Wang, Q.; Li, B.; Hu, Y.; Zhao, C.; Yin, Y. Literature-Based Phenotype Survey and In Silico Genotype Investigation of Antibiotic Resistance in the Genus Bifidobacterium. Curr. Microbiol. 2020, 77, 4104–4113. [Google Scholar] [CrossRef] [PubMed]
- Kierzkowska, M.; Majewska, A.; Mlynarczyk, G. Trends and Impact in Antimicrobial Resistance Among Bacteroides and Parabacteroides Species in 2007–2012 Compared to 2013–2017. Microb. Drug Resist. 2020, 26, 1452–1457. [Google Scholar] [CrossRef] [PubMed]
- Jian, C.; Carpén, N.; Helve, O.; de Vos, W.M.; Korpela, K.; Salonen, A. Early-life gut microbiota and its connection to metabolic health in children: Perspective on ecological drivers and need for quantitative approach. EBioMedicine 2021, 69, 103475. [Google Scholar] [CrossRef]
- Mizutani, T.; Aboagye, S.Y.; Ishizaka, A.; Afum, T.; Mensah, G.I.; Asante-Poku, A.; Asandem, D.A.; Parbie, P.K.; Abana, C.Z.; Kushitor, D.; et al. Gut microbiota signature of pathogen-dependent dysbiosis in viral gastroenteritis. Sci. Rep. 2021, 11, 13945. [Google Scholar] [CrossRef] [PubMed]
- Stewart, C.J.; Ajami, N.J.; O’Brien, J.L.; Hutchinson, D.S.; Smith, D.P.; Wong, M.C.; Ross, M.C.; Lloyd, R.E.; Doddapaneni, H.; Metcalf, G.A.; et al. Temporal development of the gut microbiome in early childhood from the TEDDY study. Nature 2018, 562, 583–588. [Google Scholar] [CrossRef]
- Vandenplas, Y.; Carnielli, V.P.; Ksiazyk, J.; Luna, M.S.; Migacheva, N.; Mosselmans, J.M.; Picaud, J.C.; Possner, M.; Singhal, A.; Wabitsch, M. Factors affecting early-life intestinal microbiota development. Nutrition 2020, 78, 110812. [Google Scholar] [CrossRef]
- Gren, C.; Spiegelhauer, M.R.; Rotbain, E.C.; Ehmsen, B.K.; Kampmann, P.; Andersen, L.P. Ruminococcus gnavus bacteraemia in a patient with multiple haematological malignancies. Access Microbiol. 2019, 1, e000048. [Google Scholar] [CrossRef] [PubMed]
- Png, C.W.; Lindén, S.K.; Gilshenan, K.S.; Zoetendal, E.G.; McSweeney, C.S.; Sly, L.I.; McGuckin, M.A.; Florin, T.H. Mucolytic bacteria with increased prevalence in IBD mucosa augment in vitro utilization of mucin by other bacteria. Am. J. Gastroenterol. 2010, 105, 2420–2428. [Google Scholar] [CrossRef]
- O’Toole, P.; Claesson, M. Gut microbiota: Changes throughout the lifespan from infancy to elderly. Int. Dairy J. 2010, 20, 281–291. [Google Scholar] [CrossRef]
- Blanton, L.V.; Charbonneau, M.R.; Salih, T.; Barratt, M.J.; Venkatesh, S.; Ilkaveya, O.; Subramanian, S.; Manary, M.J.; Trehan, I.; Jorgensen, J.M.; et al. Gut bacteria that prevent growth impairments transmitted by microbiota from malnourished children. Science 2016, 351, 6275. [Google Scholar] [CrossRef]
- Ezaki, T. Ruminococcus. In Bergey’s Manual of Systematics of Archaea and Bacteria; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012; pp. 1–5. [Google Scholar]
- Saxena, R.K.; Anand, P.; Saran, S.; Isar, J.; Agarwal, L. Microbial production and applications of 1,2-propanediol. Indian J. Microbiol. 2010, 50, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Scott, K.P.; Martin, J.C.; Campbell, G.; Mayer, C.D.; Flint, H.J. Whole-genome transcription profiling reveals genes up-regulated by growth on fucose in the human gut bacterium “Roseburia inulinivorans”. J. Bacteriol. 2006, 188, 4340–4349. [Google Scholar] [CrossRef] [PubMed]
- Vacca, M.; Celano, G.; Calabrese, F.M.; Portincasa, P.; Gobbetti, M.; De Angelis, M. The Controversial Role of Human Gut Lachnospiraceae. Microorganisms 2020, 8, 573. [Google Scholar] [CrossRef]
- Sanna, S.; van Zuydam, N.R.; Mahajan, A.; Kurilshikov, A.; Vich Vila, A.; Võsa, U.; Mujagic, Z.; Masclee, A.A.M.; Jonkers, D.; Oosting, M.; et al. Causal relationships among the gut microbiome, short-chain fatty acids and metabolic diseases. Nat. Genet. 2019, 51, 600–605. [Google Scholar] [CrossRef]
- Dragonieri, S.; Schot, R.; Mertens, B.J.; Le Cessie, S.; Gauw, S.A.; Spanevello, A.; Resta, O.; Willard, N.P.; Vink, T.J.; Rabe, K.F.; et al. An electronic nose in the discrimination of patients with asthma and controls. J. Allergy Clin. Immunol. 2007, 120, 856–862. [Google Scholar] [CrossRef]
- Tenero, L.; Sandri, M.; Piazza, M.; Paiola, G.; Zaffanello, M.; Piacentini, G. Electronic nose in discrimination of children with uncontrolled asthma. J. Breath Res. 2020, 14, 046003. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| B1 | B2 | B3 | B4 | B5 | 20 January 2021 | |
|---|---|---|---|---|---|---|
| Ammonia (µmol/L) | 73 | 75 | 91 | 98 | 34 | 79 |
| C0 plasma (µM) | 57.58 | 59.52 | 30.55 | 35.59 | 49.92 | 48.61 |
| C3 plasma (µM) | 80.95 | 58.17 | 40.68 | 49.69 | 35.17 | 52.79 |
| C3 DBS (µM) | 59.38 | 31.99 | 20.63 | |||
| MCA DBS (µM) | 37.5 | 12.9 | 16.8 | 17.4 | 7.6 | 11.3 |
| Glycine plasma (µM) | 939 | 429 | 768 | 1092 | 913 | 1030 |
| protein (g/kg/die | 1.52 | 1.16 | 1.19 | 1.36 | 1.18 | 1.14 |
| Energy (kcal/kg) | 131 | 128 | 88 | 90 | 82 | 83 |
| Weight (kg) | 3.016 | 3.950 | 6.450 | 7.5 | 7.6 | 8.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bordugo, A.; Salvetti, E.; Rodella, G.; Piazza, M.; Dianin, A.; Amoruso, A.; Piacentini, G.; Pane, M.; Torriani, S.; Vitulo, N.; et al. Assessing Gut Microbiota in an Infant with Congenital Propionic Acidemia before and after Probiotic Supplementation. Microorganisms 2021, 9, 2599. https://doi.org/10.3390/microorganisms9122599
Bordugo A, Salvetti E, Rodella G, Piazza M, Dianin A, Amoruso A, Piacentini G, Pane M, Torriani S, Vitulo N, et al. Assessing Gut Microbiota in an Infant with Congenital Propionic Acidemia before and after Probiotic Supplementation. Microorganisms. 2021; 9(12):2599. https://doi.org/10.3390/microorganisms9122599
Chicago/Turabian StyleBordugo, Andrea, Elisa Salvetti, Giulia Rodella, Michele Piazza, Alice Dianin, Angela Amoruso, Giorgio Piacentini, Marco Pane, Sandra Torriani, Nicola Vitulo, and et al. 2021. "Assessing Gut Microbiota in an Infant with Congenital Propionic Acidemia before and after Probiotic Supplementation" Microorganisms 9, no. 12: 2599. https://doi.org/10.3390/microorganisms9122599
APA StyleBordugo, A., Salvetti, E., Rodella, G., Piazza, M., Dianin, A., Amoruso, A., Piacentini, G., Pane, M., Torriani, S., Vitulo, N., & Felis, G. E. (2021). Assessing Gut Microbiota in an Infant with Congenital Propionic Acidemia before and after Probiotic Supplementation. Microorganisms, 9(12), 2599. https://doi.org/10.3390/microorganisms9122599